Growth Response of Sessile Oak and European Hornbeam to Traditional Coppice-with-Standards Management

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Measurements
2.4. Statistical Analyses
3. Results
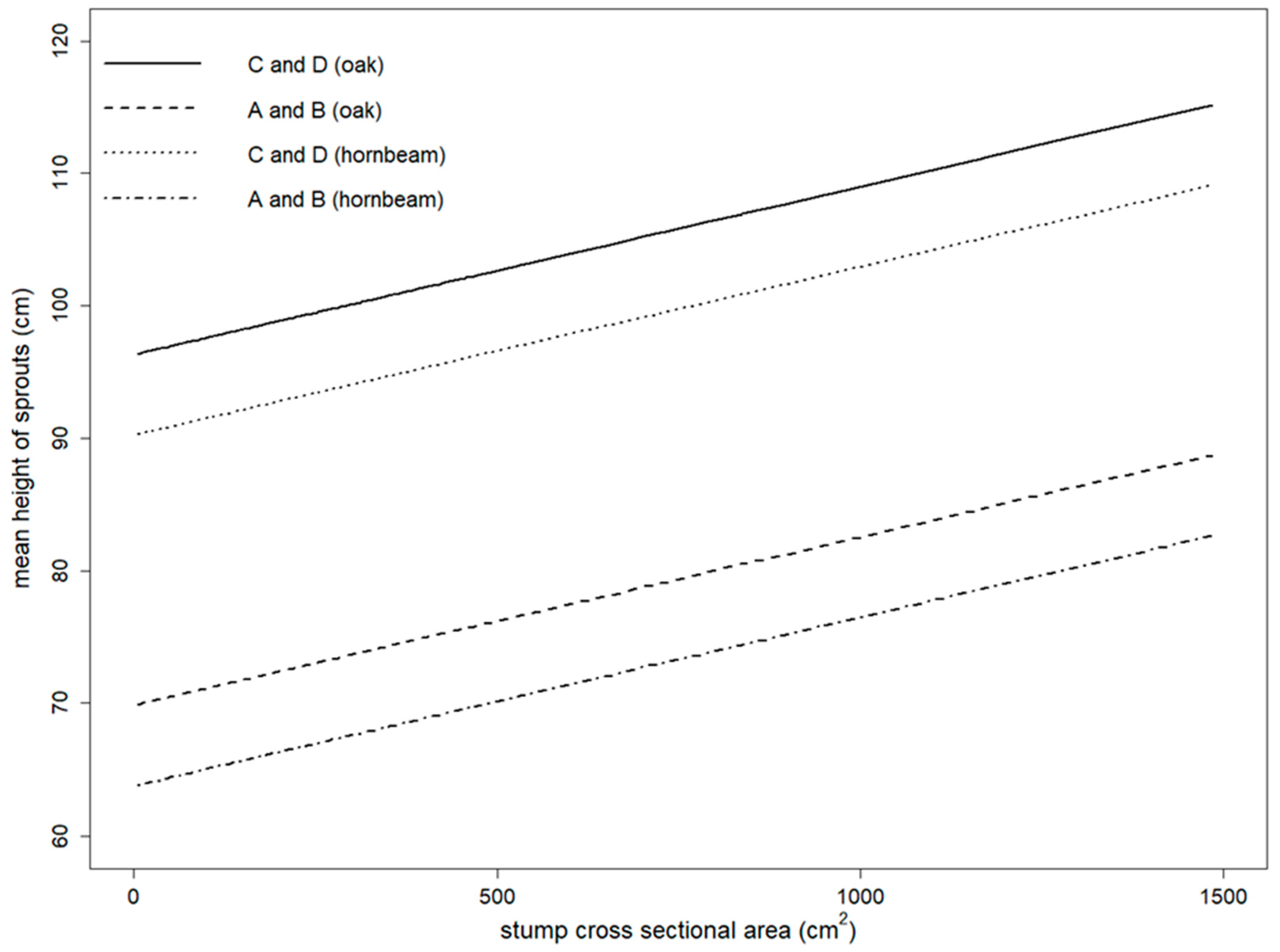
3.1. Growth of Sprouts
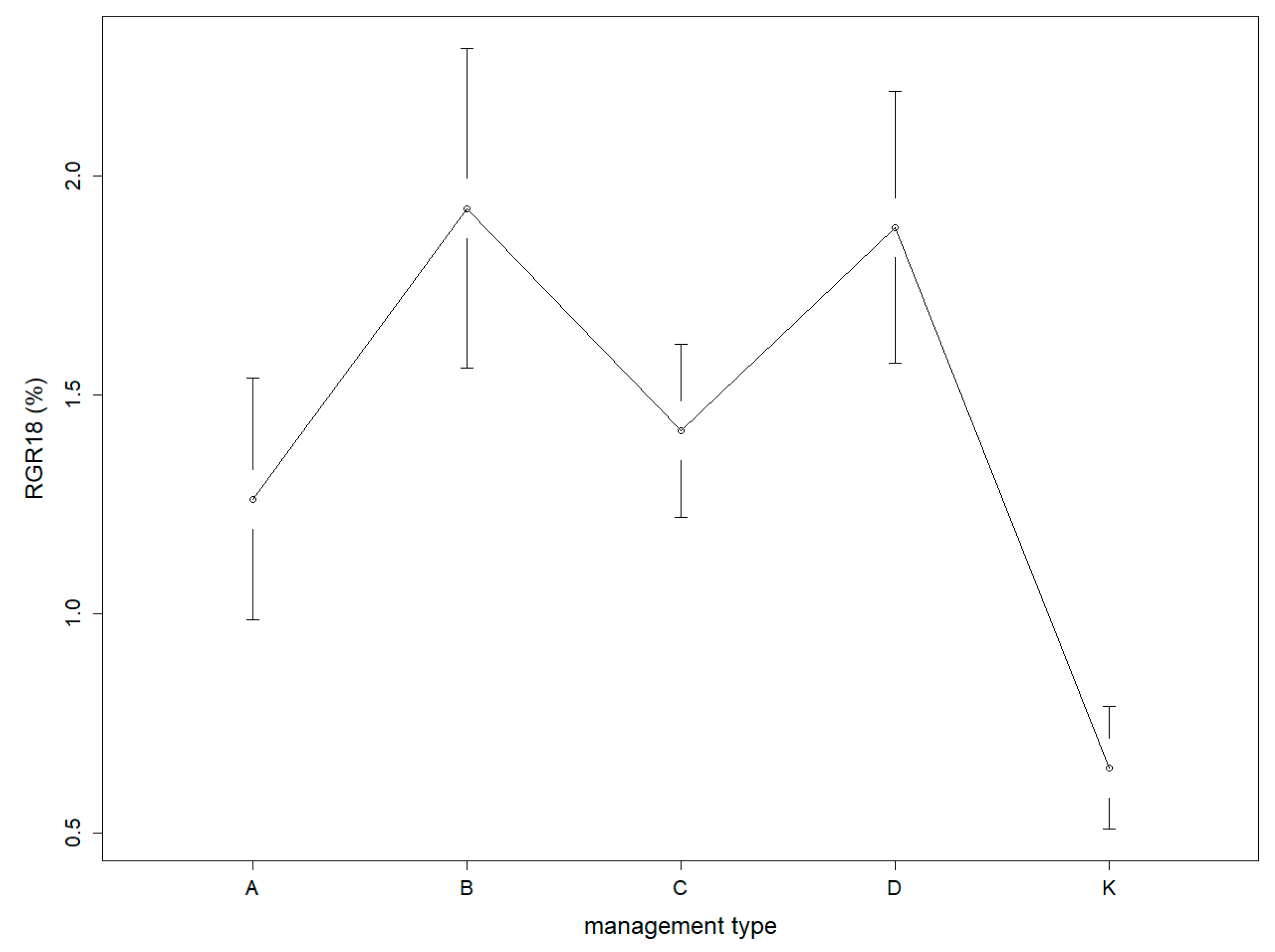
3.2. Growth of Sessile Oak Standards
4. Discussion
4.1. Impact of Livestock Grazing and Litter Raking on Coppices and Landscape Development
4.2. Growth of Sprouts in Coppices
4.3. Growth of the Standards in the Coppices
5. Conclusions
- the number of sprouts per stump was positively affected by the stump cross sectional area and applied management types; species had no effect on the number of sprouts.
- the mean diameter and the mean height of the sprouts were correlated with all of the involved factors, i.e., stump cross sectional area, applied management, and tree species as
- bigger stumps produced thicker and higher sprouts;
- the influence of traditional management types on sprout dimensions was significant as
- only the simultaneous application of raking and grazing decreased the mean sprout diameter, and
- the application of grazing (either individual or combined with raking) reduced the mean heights of sprouts; and
- European hornbeam produced thinner and lower sprouts than sessile oak.
- Standards responded to harvest by increased relative circumference increment. Only the simultaneous effect of grazing and raking decreased the circumference increment significantly.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Szabó, P.; Müllerová, J.; Silvie Suchánková, S.; Kotacka, M. Intensive woodland management in the Middle Ages: spatial modelling based on archival data. J. Hist Geogr. 2015, 48, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeffer, A. Hrabání steliva v lesích a lesní pastva v Normálních Dobách: Studie Lesnická, Národohospodářská a Vodohospodářská, 1st ed.; Masarykova Akademie Práce: Praha, Czech Republic, 1948; p. 32. [Google Scholar]
- Vild, O.; Stejskal, R. Vliv experimentální pastvy na lesní podrost v Národním Parku Podyjí. Thayensia 2013, 10, 27–38. [Google Scholar]
- Vild, O.; Kalwij, J.M.; Hédl, R. Effects of simulated historical tree litter raking on the understorey vegetation in a central European forest. Appl. Veg. Sci. 2015, 18, 569–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debussche, M.; Debussche, G.; Lepart, J. Changes in the vegetation of Quercus pubescens woodland after cessation of coppicing and grazing. J. Veg. Sci. 2001, 2, 81–92. [Google Scholar] [CrossRef]
- Evans, J. Coppice forestry–an overview. In Ecology and Management of Coppice Woodlands, 1st ed.; Buckley, G.P., Ed.; Chapman & Hall: London, UK, 1992; pp. 18–27. [Google Scholar]
- Müllerová, J.; Szabó, P.; Hédl, R. The rise and fall of traditional forest management in southern Moravia: A history of the past 700 years. For. Ecol. Manag. 2014, 331, 104–115. [Google Scholar] [Green Version]
- Zhu, W.Z.; Xiang, J.S.; Wang, S.G.; Li, M.H. Resprouting ability and mobile carbohydrate reserves in an oak shrubland decline with increasing elevation on the eastern edge of the Qinghai-Tibet Plateau. For. Ecol. Manag. 2012, 278, 118–126. [Google Scholar] [CrossRef]
- Carl, Ch.; Lehmann, J.R.K.; Landgraf, D.; Pretzsch, H. Robinia pseudoacacia L. in Short Rotation Coppice: Seed and Stump Shoot Reproduction as well as UAS-based Spreading Analysis. Forests 2019, 10, 235. [Google Scholar] [CrossRef]
- Vild, O.; Roleček, J.; Hédl, R.; Kopecký, M.; Utinek, D. Experimental restoration of coppice-with-standards: Response of understorey vegetation from the conservation perspective. For. Ecol. Manag. 2013, 310, 234–241. [Google Scholar] [CrossRef] [Green Version]
- Rydberg, D.; Falck, J. Urban forestry in Sweden from a silvicultural perspective: A review. Landsc. Urban. Plan. 2000, 47, 1–18. [Google Scholar] [CrossRef]
- Van Calster, H.; Baeten, L.; Verheyen, K.; De Keersmaeker, L.; Dekeyser, S.; Rogister, J.E.; Hermy, M. Diverging effects of overstorey conversion scenarios on the understorey vegetation in a former coppice-with-standards forest. For. Ecol. Manag. 2008, 256, 519–528. [Google Scholar] [CrossRef]
- Buček, A. Krajina České republiky a pastva. Veronica 2000, 14, 1–7. [Google Scholar]
- Mitchell, F.J.G.; Kirby, K.J. The Impact of Large Herbivores on the Conservation of Semi-natural Woods in the British Upland. Forestry 1990, 63, 333–353. [Google Scholar] [CrossRef]
- Hester, A.J.; Mitchell, F.J.G.; Kirby, K.J. Effects of season and intensity of sheep grazing on tree regeneration in a British upland woodland. For. Ecol. Manag. 1996, 88, 99–106. [Google Scholar] [CrossRef]
- Mládek, J.; Pavlů, V.; Hejcman, M.; Gaisler, J. Pastva jako prostředek údržby trvalých travních porostů v chráněných územích, 1st ed.; Výzkumný ústav rostlinné výroby: Praha, Czech Republic, 2006; p. 104. [Google Scholar]
- Mitscherlich, G. Untersuchungen über das Wachstumder Kiefer in Baden. 2. Teil: Die Streunutzungs- und Düngungsversuche. Allgemeine Forst- und Jagdzeitung 1955, 126, 193–204. [Google Scholar]
- Glatzel, G. The impact of historic land use and modern forestry on nutrient relations of Central European forest ecosystems. Fert. Res. 1991, 27, 1–8. [Google Scholar] [CrossRef]
- Kilian, W. Forest site degradation-temporary deviation from the natural site potential. Ecol. Eng. 1998, 10, 5–18. [Google Scholar] [CrossRef]
- Ebermayer, E. Die gesammte Lehre der Waldstreu mit Rücksicht auf die chemische Statik des Waldbaues, 1st ed.; Springer: Berlin, Germany, 1876; p. 416. [Google Scholar]
- Tschermak, L. Buchenlaubstreunutzung im Lande Salzburg. Centralblatt für das Gesamte Forstwesen 1926, 52, 336–349 & 577–663. [Google Scholar]
- Kreutzer, K. Über den Einfluss der Streunutzung auf den Stickstoffhaushalt von Kiefernbeständen (Pinus silvestris L.). Forstwiss. Cent.bl. 1972, 91, 263–270. [Google Scholar] [CrossRef]
- Bürgi, M. A case study of forest change in the Swiss lowlands. Landsc. Ecol. 1999, 14, 567–575. [Google Scholar] [CrossRef]
- Šilc, U.; Čarni, A.; Košir, P.; Marinšek, A.; Zelnik, I. Litter-raking forests in SE Slovenia and in Croatia. Hacquetia 2008, 7, 71–88. [Google Scholar] [CrossRef]
- Kadavý, J.; Kneifl, M.; Knott, R.; Hurt, V.; Flora, M. Možnosti a limity hospodaření s nízkým a středním lesem a jejich vliv na biodiverzitu. In Ochrana Přírody a Krajiny v České Republice; Vybrané aktuální problémy a možnosti jejich řešení. I. díl, Olomouc, Česká republika; Machar, I., Drobilová, L., Eds.; Univerzita Palackého v Olomouci: Olomouc, Czech Republic, 2012; pp. 290–300. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014. In International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 1st ed.; World Soil Resources Reports No. 106; FAO: Rome, Italy, 2014; p. 203. [Google Scholar]
- Jabiol, B.; Brêthes, A.; Ponge, J.F.; Toutain, F.; Brun, J.J. L´humus sous toutes ses forms, 1st ed.; École Nationale du Gńie Rural, des Eaux et des Forêts: Nancy, France, 1995; p. 63. [Google Scholar]
- ÚHÚL; VÚLHM. Taxační tabulky (Forest Taxation Tables), 1st ed.; Forest Management Institute and Forestry and Game Management Research Institute: Jíloviště-Strnady, Czech Republic, 1990; p. 32. [Google Scholar]
- Pretzsch, H.; Biber, P. A Re-Evaluation of Reineke’s Rule and Stand Density Index. For. Sci. 2005, 51, 304–320. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Savaliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009; p. 574. [Google Scholar]
- Akaike, H. Information theory and an extension of the maximum likelihood principle. In Proceedings of the 2nd International Symposium on Information Theory, Budapest, Hungary, 2–8 September 1973; Petrov, B.N., Csaki, F., Eds.; Akademiai Kiado: Budapest, Hungary, 1973; pp. 268–281. [Google Scholar]
- Nagelkerke, N.J.D. A note on a general definition of the coefficient of determination. Biometrika 1991, 78, 691–692. [Google Scholar] [CrossRef]
- Cotillas, M.; Sabaté, S.; Gracia, C.; Espelta, J.M. Growth response of mixed mediterranean oak coppices to rainfall reduction: Could selective thinning have any influence on it? For. Ecol. Manag. 2009, 258, 1677–1683. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: http://www.R-project.org/ (accessed on 18 February 2019).
- Van Uytvanck, J.; Hoffmann, M. Impact of grazing management with large herbivores on forest ground flora and bramble understorey. Acta Oecol. 2009, 35, 523–532. [Google Scholar] [CrossRef]
- McEvoy, P.M.; Flexen, M.; McAdam, J.H. The effects of livestock grazing on ground flora in broadleaf woodlands in Northern Ireland. For. Ecol. Manag. 2006, 225, 39–50. [Google Scholar] [CrossRef]
- Papachristou, T.G.; Platis, P.D. The impact of cattle and goats grazing on vegetation in oak stands of varying coppicing age. Acta Oecol. 2011, 37, 16–22. [Google Scholar] [CrossRef]
- Hart, S.P. Recent perspectives in using goats for vegetation management in the USA. J. Dairy Sci. 2001, 84 (Electronic Suppl.), E170–E176. [Google Scholar] [CrossRef]
- Peacok, C.; Sherman, D.M. Sustainable goat production. Some global perspectives. Small Rumin. Res. 2010, 89, 78–80. [Google Scholar] [CrossRef]
- Cochillo-Hilario, M.; Wrage-Moenning, N.; Isselstein, J. Forage selectivity by cattle and sheep co-grazing swards differing in plant species diversity. Grass Forage Sci. 2018, 73, 320–329. [Google Scholar] [CrossRef]
- Onatibia, G.R.; Aguiar, M.R. Grasses and grazers in arid rangelands: Impact of sheep management on forage and non-forage grass populations. J. Environ. Manag. 2019, 235, 42–50. [Google Scholar] [CrossRef]
- Flota-Banuelos, C.; Rivera-Lorca, J.A.; Candelaria-Martinez, B. Importance of sheep social hierarchy on feeding behavior and parasite load in silvopastoral and grass monoculture grazing systems. Rev. Mex. Cienc. Pecu. 2019, 10, 52–67. [Google Scholar]
- Bojkovski, D.; Štuhec, I.; Kompan, D.; Zupan, M. The behavior of sheep and goats co-grazing on pasture with different types of vegetation in the karst region. J. Anim. Sci. 2014, 92, 2752–2758. [Google Scholar] [CrossRef] [PubMed]
- Gimmi, U.; Poulter, B.; Wolf, A.; Portner, H.; Weber, P.; Bürgi, M. Soil carbon pools in Swiss forests show legacy effects from historic forest litter raking. J. Landsc. Ecol. 2013, 28, 835–846. [Google Scholar] [CrossRef]
- Del Tredici, P. Sprouting in temperate trees: A morphological and ecological review. Bot. Rev. 2001, 67, 121–140. [Google Scholar] [CrossRef]
- Vesk, P.A.; Westoby, M. Sprouting ability across diverse disturbances and vegetation types worldwide. J. Ecol. 2004, 92, 310–320. [Google Scholar] [CrossRef]
- Matula, R.; Svátek, M.; Kůrová, J.; Úradníček, L.; Kadavý, J.; Kneifl, M. The sprouting ability of the main tree species in Central European coppices: implications for coppice restoration. Eur. J. Forest. Res. 2012, 131, 1501–1511. [Google Scholar] [CrossRef]
- Šplíchalová, M.; Adamec, Z.; Kadavý, J.; Kneifl, M. Probability model of sessile oak (Quercus petraea (Matt.) Liebl.) stump sprouting in the Czech Republic. Eur. J. Forest. Res. 2012, 131, 1611–1618. [Google Scholar] [CrossRef]
- Johnson, P.S.; Shifley, S.R.; Rogers, R. The Ecology and Silviculture of Oaks, 2nd ed.; CAB International: Wallingford, UK, 2009; p. 580. [Google Scholar]
- Pietras, J.; Stojanović, M.; Knott, R.; Pokorný, R. Oak sprouts grow better than seedlings under drought stress. iForest 2016, 9, 529–535. [Google Scholar] [CrossRef] [Green Version]
- Stojanović, M.; Szatniewska, J.; Kyselová, I.; Pokorný, R.; Čater, M. Transpiration and water potential of young Quercus petraea (M.) Liebl. coppice sprouts and seedlings during favourable and drought conditions. J. For. Sci. 2017, 63, 313–323. [Google Scholar]
- Weigel, D.R.; Peng, C.-Y.J. Predicting stump sprouting and competitive success of five oak species in southern Indiana. Can. J. For. Res. 2002, 32, 703–712. [Google Scholar] [CrossRef]
- Reimoser, F.; Armstrong, H.; Suchant, R. Measuring forest damage of ungulates: what should be considered. For. Ecol. Manag. 1999, 120, 47–58. [Google Scholar] [CrossRef]
- Bullock, D.J.; Grabrovaz, M. The impact of goat farming on upland vegetation in Scotland—A preliminary survey. In Rural Development Report, 1st ed.; Nature Conservancy Council: Edinburgh, UK, 1990; Volume 1, pp. 1–28. [Google Scholar]
- Dale, M.E. Growth Response from Thinning Young Even-Aged White Oaks Stands; USDA Forest Service Research Paper NE-112; USDA Forest Service, Northeastern Forest Experiment Station: Newtown Square, PA, USA, 1968; p. 20.
- Hilt, D.E. Diameter Growth of Upland Oaks after Thinning; USDA Forest Service Research Paper NE-437; USDA Forest Service, Northeastern Forest Experiment Station: Newtown Square, PA, USA, 1979; p. 12.
- Hilt, D.E. Individual-Tree Diameter Growth Model for Managed, Even-Aged, Upland Oak Stands; USDA Forest Service Research Paper NE-533; USDA Forest Service, Northeastern Forest Experiment Station: Newtown Square, PA, USA, 1983; p. 15.
- Mitchell, R.J.; Dwyer, J.P.; Musbach, R.A.; Garrett, H.E.; Cox, G.S.; Kurtz, W.B. Crop tree release of a scarlet-black oak stand. North. J. Appl. For. 1988, 5, 96–99. [Google Scholar]
- Dzwonko, Z.; Gawroński, S. Effect of litter removal on species richness and acidification of a mixed oak-pine woodland. Biol. Cons. 2002, 106, 389–398. [Google Scholar] [CrossRef]
- Sayer, E.J. Using experimental manipulation to assess the roles of leaf litter in the functioning of forest ecosystems. Biol. Rev. 2006, 81, 1–31. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Label | Harvest | Treatment | Age (years) | Slope (°)/Exposition (°) | Density (tree ha−1) | SDI | Volume (m3 ha−1) |
|---|---|---|---|---|---|---|---|
| K1 | Control (K) | 52 | 7.0/245 | 1467 | 589 | 186 | |
| K2 | Not harvested | 40 | 10.5/350 | 1875 | 552 | 152 | |
| K3 | 66 | 13.0/270 | 1433 | 613 | 209 | ||
| 2A | C-W-S + raking + grazing (A) | 38 | 8.0/200 | 1933 | 569 | 129 | |
| 3A | Harvested | 40 | 6.0/0 | 1433 | 490 | 149 | |
| 5A | 38 | 10.5/270 | 1733 | 599 | 149 | ||
| 2B | C-W-S + grazing (B) | 38 | 7.0/210 | 2042 | 459 | 76 | |
| 3B | 40 | 5.5/0 | 1283 | 522 | 177 | ||
| 5B | 38 | 12.5/290 | 1225 | 516 | 148 | ||
| 1A | C-W-S + raking (C) | 40 | 5.5/345 | 1583 | 507 | 146 | |
| 4A | 38 | 9.0/270 | 1325 | 510 | 156 | ||
| 6A | 26 | 21.0/290 | 1792 | 542 | 124 | ||
| 1B | C-W-S (D) | 40 | 16.0/340 | 1833 | 603 | 177 | |
| 4B | 38 | 8.0/330 | 1233 | 465 | 147 | ||
| 6B | 26 | 19.0/270 | 1467 | 513 | 110 |
| Dependent Variable | DF | χ2 | F | p | AIC | R2 | Pseudo R2 |
|---|---|---|---|---|---|---|---|
| N | 4 | 24.11 | <0.0001 | 2385.3 | 0.0886 | ||
| d | 5; 254 | 15.55 | <0.0001 | 1124.25 | 0.2344 | ||
| h | 5; 254 | 24.93 | <0.0001 | 2321.41 | 0.3292 |
| Dependent Variable | Independent Variable | Parameter | ESTIMATION | SE | t | z | p |
|---|---|---|---|---|---|---|---|
| N | INT | β0 | 3.4801 | 0.0810 | 42.977 | <0.0001 | |
| SA | β1 | 0.0006 | 0.0001 | 4.101 | <0.0001 | ||
| C | β2 | 0.2792 | 0.0982 | 2.844 | 0.0045 | ||
| B | β3 | 0.1915 | 0.0999 | 1.917 | 0.0552 | ||
| A | β4 | 0.2718 | 0.0984 | 2.760 | 0.0058 | ||
| d | INT | β0 | 10.5928 | 0.3493 | 30.327 | <0.0001 | |
| SA | β1 | 0.0018 | 0.0006 | 3.213 | 0.0015 | ||
| SH | β2 | −1.3821 | 0.2705 | −5.108 | <0.0001 | ||
| C | β3 | −0.0724 | 0.3605 | −0.201 | 0.8411 | ||
| B | β4 | −0.5319 | 0.3663 | −1.452 | 0.1477 | ||
| A | β5 | −1.5259 | 0.3615 | −4.220 | <0.0001 | ||
| h | INT | β0 | 96.2987 | 3.4916 | 27.580 | <0.0001 | |
| SA | β1 | 0.0127 | 0.0057 | 2.237 | 0.0262 | ||
| SH | β2 | −6.0598 | 2.7045 | −2.241 | 0.0259 | ||
| C | β3 | 2.4656 | 3.6038 | 0.684 | 0.4945 | ||
| B | β4 | −23.4180 | 3.6619 | −6.395 | <0.0001 | ||
| A | β5 | −26.4539 | 3.6141 | −7.320 | <0.0001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kadavý, J.; Adamec, Z.; Uherková, B.; Kneifl, M.; Knott, R.; Kučera, A.; Friedl, M.; Dařenová, E.; Skládanka, J.; Drápela, K. Growth Response of Sessile Oak and European Hornbeam to Traditional Coppice-with-Standards Management. Forests 2019, 10, 515. https://doi.org/10.3390/f10060515
Kadavý J, Adamec Z, Uherková B, Kneifl M, Knott R, Kučera A, Friedl M, Dařenová E, Skládanka J, Drápela K. Growth Response of Sessile Oak and European Hornbeam to Traditional Coppice-with-Standards Management. Forests. 2019; 10(6):515. https://doi.org/10.3390/f10060515
Chicago/Turabian StyleKadavý, Jan, Zdeněk Adamec, Barbora Uherková, Michal Kneifl, Robert Knott, Aleš Kučera, Michal Friedl, Eva Dařenová, Jiří Skládanka, and Karel Drápela. 2019. "Growth Response of Sessile Oak and European Hornbeam to Traditional Coppice-with-Standards Management" Forests 10, no. 6: 515. https://doi.org/10.3390/f10060515