Tree Biomass Equations from Terrestrial LiDAR: A Case Study in Guyana

 ,
,  ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
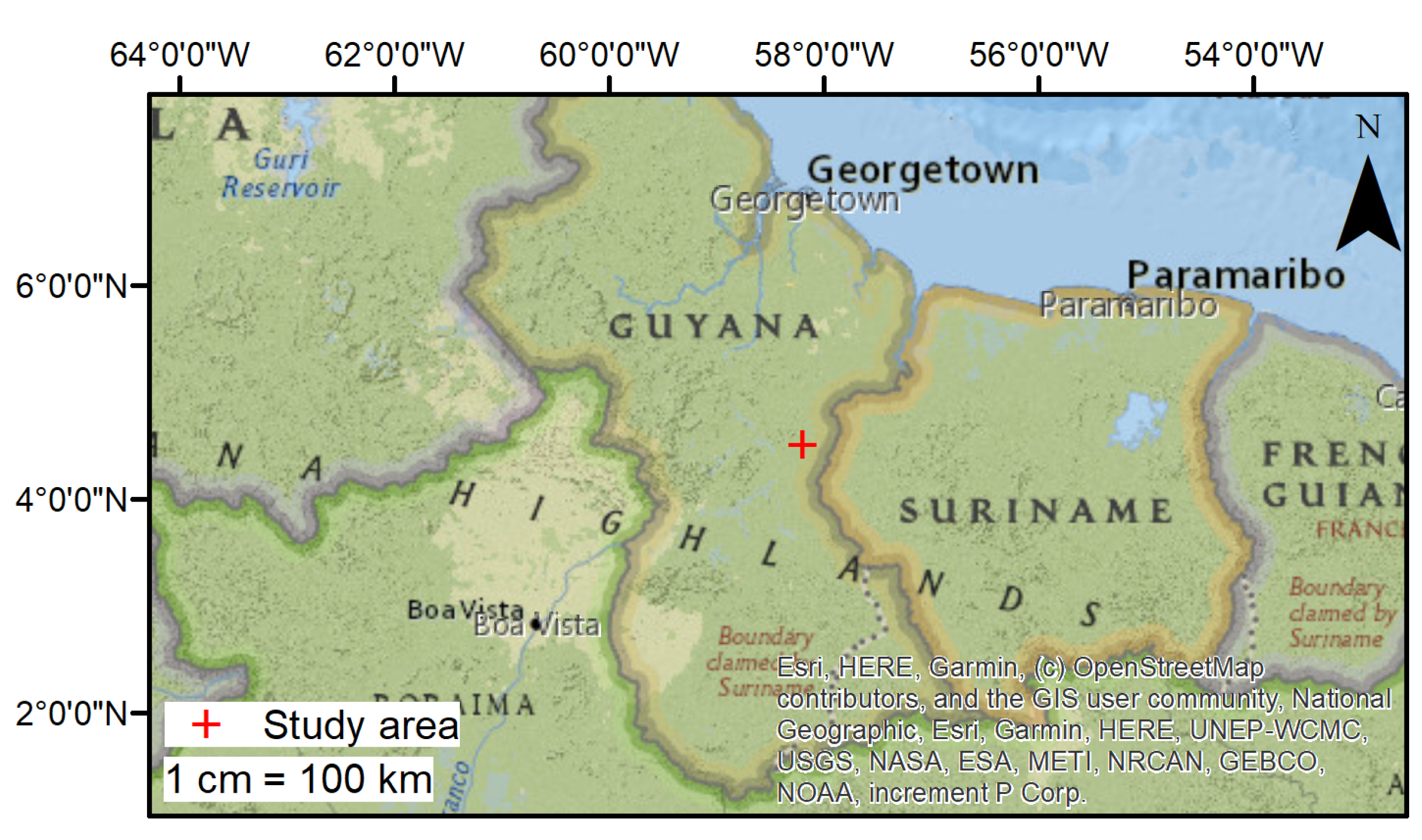
2.1. Study Area
2.2. Tree Selection and Data Collection
2.2.1. Tree Inventory
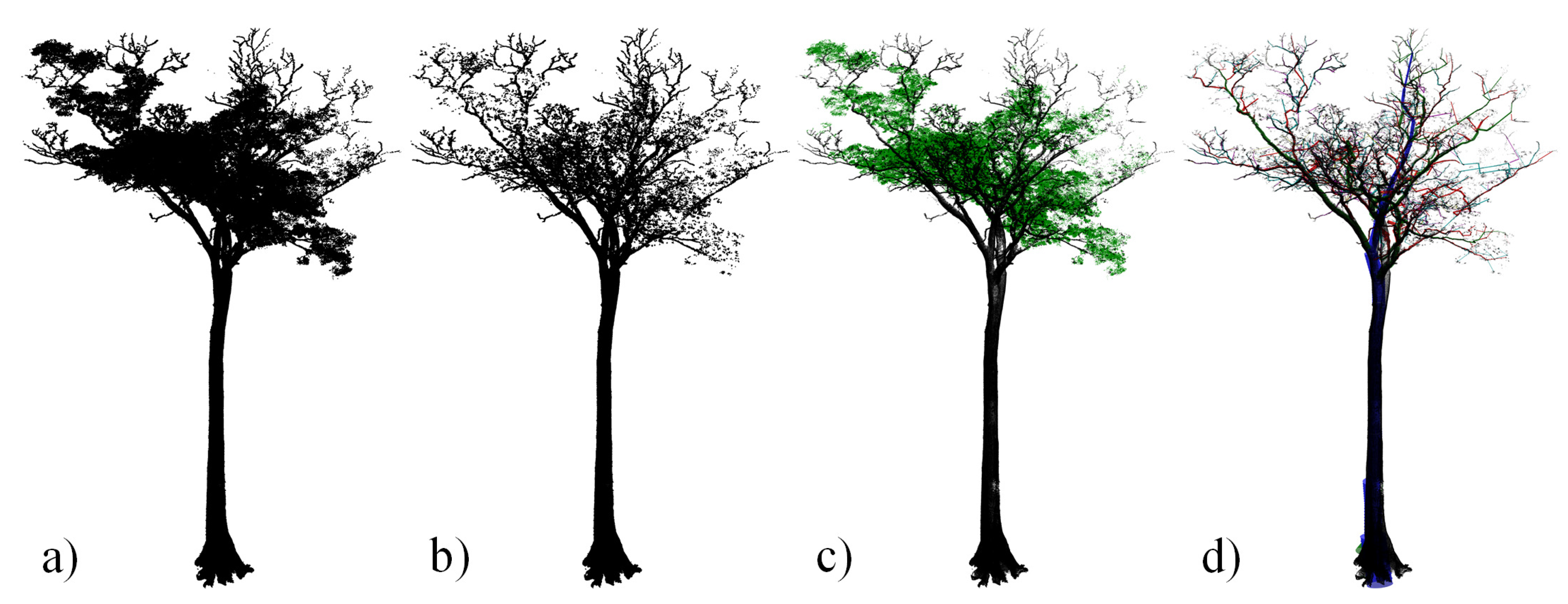
2.2.2. TLS Data Acquisition
2.2.3. Destructive Harvesting and Fresh Mass Sampling
2.2.4. Laboratory Analysis
2.3. Diameter, Tree Height and Crown Diameter from TLS Data
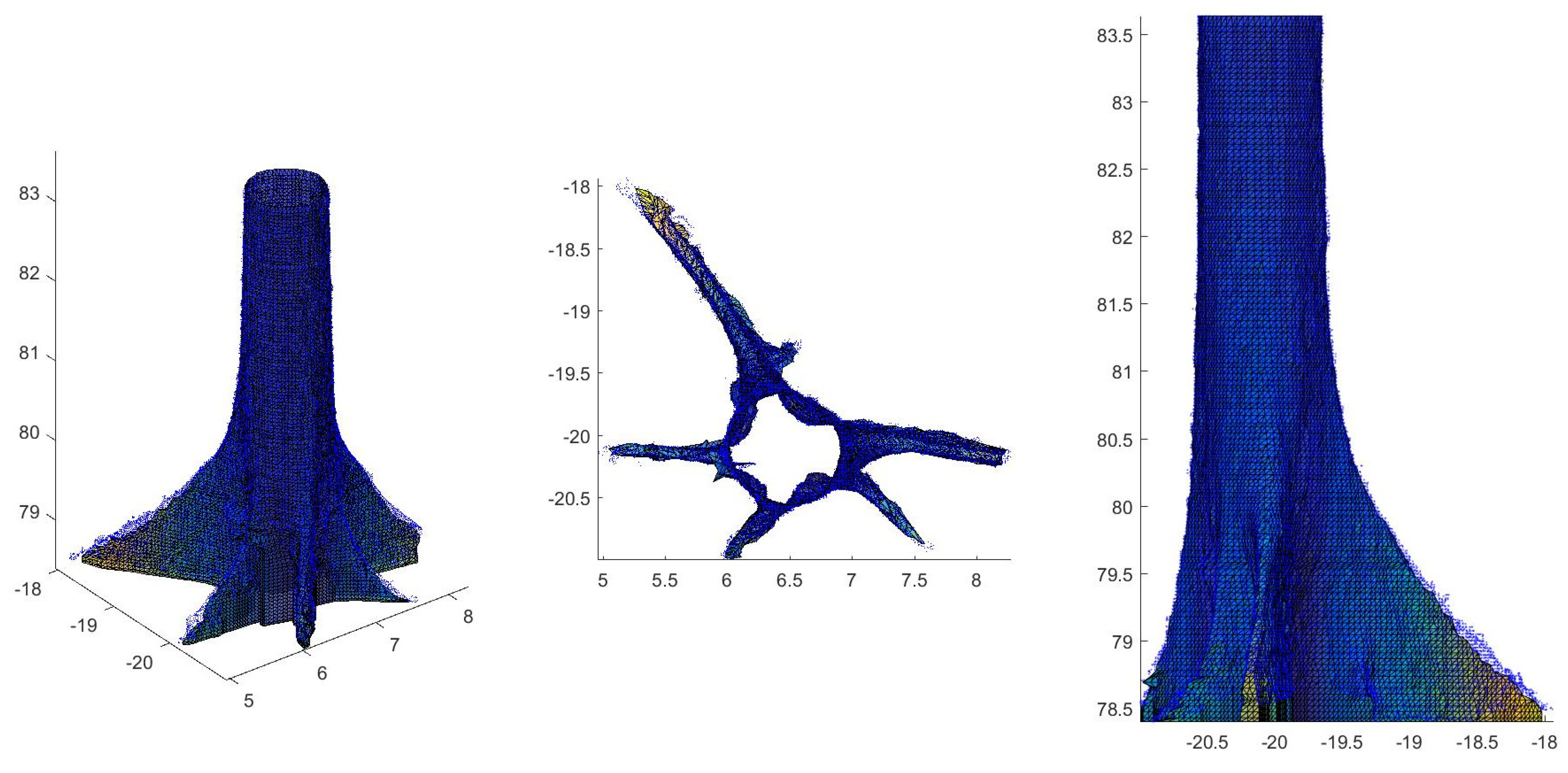
2.4. Tree Volume and Biomass from TLS Data
2.5. TLS-Derived Allometric Models
2.6. Tree Aboveground Biomass Estimation from Pantropical Allometric Models
2.7. Assessment of Allometric Models
3. Results
3.1. Tree Attributes and Estimated Biomass
3.2. Allometric Models Using TLS-Derived Measurements
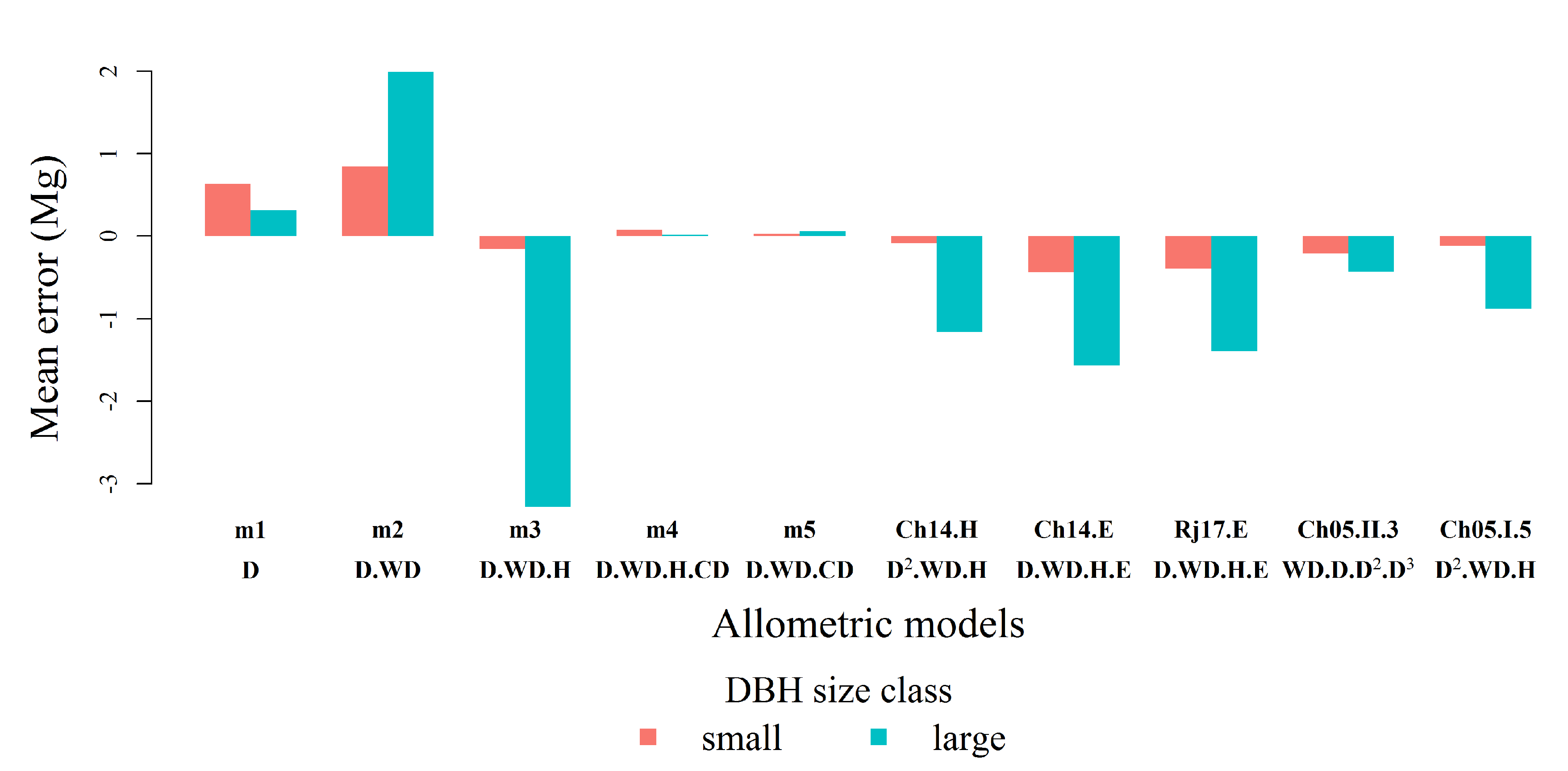
3.3. Evaluation of Allometric Models
4. Discussion
4.1. Developing Allometric Models from TLS-Derived Attributes
4.2. Choosing the Adequate Tree Attributes for Allometric Models
4.3. Local or Pantropical Allometric Models?
4.4. Challenges and Outlook
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AGB | Aboveground biomass |
| adj- | Adjusted R-square |
| AICc | Akaike’s information criterion |
| CCC | Concordance correlation coefficient |
| CD | Crown diameter |
| CF | Correction factor |
| Ch05.I.5 | Chave et al. [12] Equation I.5 |
| Ch05.II.3 | Chave et al. [12] Equation II.3 |
| Ch14.E | Chave et al. [11] Equation (7) |
| Ch14.H | Chave et al. [11] Equation (4) |
| CV RMSE | Coefficient of variation of RMSE |
| D | Diameter at breast height |
| df | degrees of freedom |
| dmf | dry mass fraction |
| GWDD | Global wood density database [8] |
| H | Height |
| LiDAR | Light Detection And Ranging |
| MMRV | Monitoring, measurement, reporting and verification |
| POM | Point of measurement |
| Rj17.E | Réjou-Méchain et al. [46] Equation (1) |
| QSM | Quantitative structure models |
| TLS | Terrestrial laser scanning |
| RDVC | Reference Dummy Variable Correction |
| REDD+ | Reducing emissions from deforestation and degradation |
| RMSE | Root mean square error |
| RSE | Residual standard error |
| UAV | Unnamed aerial vehicle |
| UAV-LS | Unnamed aerial vehicle laser scanning |
| WD | Wood density |
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Type | RMSE | CCC | Mean Error (Mg) | Sum Error (Mg) | SD Error (Mg) | Mean rel. Error (%) | SD. rel. Error (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Small | Large | Small | Large | Small | Large | Small | Large | Small | Large | Small | Large | Small | Large | Small | Large | ||
| m5 | D.WD.CD | 0.83 | 0.84 | 1.27 | 2.69 | 0.87 | 0.90 | 0.03 | 0.06 | 0.50 | 0.53 | 1.31 | 2.86 | 44.08 | 0.11 | 70.75 | 22.73 |
| m4 | D.WD.H.CD | 0.83 | 0.81 | 1.22 | 2.89 | 0.89 | 0.89 | 0.08 | 0.01 | 1.23 | 0.13 | 1.26 | 3.07 | 44.29 | −0.04 | 65.74 | 23.30 |
| Ch05.II.3 | WD.D.D.D | 0.70 | 0.78 | 1.57 | 3.30 | 0.79 | 0.86 | −0.21 | −0.43 | −3.61 | −3.88 | 1.60 | 3.47 | 10.71 | −4.22 | 57.51 | 22.36 |
| Ch05.I.5 | D.WD.H | 0.76 | 0.67 | 1.40 | 4.25 | 0.84 | 0.79 | −0.11 | −0.88 | −1.95 | −7.91 | 1.44 | 4.42 | 15.26 | −7.57 | 47.45 | 24.95 |
| Ch14.H | (D.WD.H) | 0.75 | 0.67 | 1.41 | 4.11 | 0.84 | 0.79 | −0.09 | −1.16 | −1.45 | −10.44 | 1.45 | 4.19 | 19.54 | −9.21 | 48.42 | 23.74 |
| m1 | D | 0.74 | 0.59 | 1.55 | 3.71 | 0.81 | 0.76 | 0.64 | 0.32 | 10.81 | 2.84 | 1.46 | 3.92 | 99.92 | 7.74 | 118.63 | 26.22 |
| Rj17.E | D.D.WD.E | 0.70 | 0.77 | 1.65 | 3.45 | 0.75 | 0.85 | −0.40 | −1.39 | −6.72 | −12.55 | 1.65 | 3.34 | 3.61 | −11.50 | 52.33 | 20.40 |
| Ch14.E | D.D.WD.E | 0.70 | 0.77 | 1.68 | 3.55 | 0.74 | 0.84 | −0.44 | −1.57 | −7.45 | −14.11 | 1.67 | 3.37 | 1.39 | −13.08 | 51.60 | 20.94 |
| m3 | D.WD.H | 0.67 | 0.74 | 1.67 | 4.39 | 0.75 | 0.72 | −0.16 | −3.27 | −2.66 | −29.45 | 1.71 | 3.11 | 35.20 | −23.72 | 65.90 | 19.41 |
| m2 | D.WD | 0.64 | 0.77 | 1.90 | 4.30 | 0.76 | 0.81 | 0.84 | 1.99 | 14.30 | 17.91 | 1.75 | 4.04 | 105.06 | 14.74 | 109.05 | 28.47 |

References
- Guyana Forestry Commission. Terms of Reference for Developing Capacities for a National Monitoring, Reporting and Verification System to Support REDD+ Participation of Guyana. Background, Capacity Assessment and Roadmap; Technical Report; Guyana Foresty Commission: Georgetown, TX, USA, 2009.
- Butt, N.; Epps, K.; Overman, H.; Iwamura, T.; Fragoso, J.M. Assessing carbon stocks using indigenous peoples’ field measurements in Amazonian Guyana. For. Ecol. Manag. 2015, 338, 191–199. [Google Scholar] [CrossRef]
- Henry, M.; Cifuentes Jara, M.; Réjou-Méchain, M.; Piotto, D.; Michel Fuentes, J.M.; Wayson, C.; Alice Guier, F.; Castañeda Lombis, H.; Castellanos López, E.; Cuenca Lara, R.; et al. Recommendations for the use of tree models to estimate national forest biomass and assess their uncertainty. Ann. For. Sci. 2015, 72, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, E.; Duque, A.; Saldarriaga, J.; Cabrera, K.; de las Salas, G.; del Valle, I.; Lema, A.; Moreno, F.; Orrego, S.; Rodríguez, L. Tree above-ground biomass allometries for carbon stocks estimation in the natural forests of Colombia. For. Ecol. Manag. 2012, 267, 297–308. [Google Scholar] [CrossRef]
- Goodman, R.C.; Phillips, O.L.; Baker, T.R. The importance of crown dimensions to improve tropical tree biomass estimates. Ecol. Appl. 2014, 24, 680–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manuri, S.; Brack, C.; Nugroho, N.P.; Hergoualc’h, K.; Novita, N.; Dotzauer, H.; Verchot, L.; Putra, C.A.S.; Widyasari, E.; Hergoualc’h, K.; et al. Tree biomass equations for tropical peat swamp forest ecosystems in Indonesia. For. Ecol. Manag. 2014, 334, 241–253. [Google Scholar] [CrossRef]
- Gibbs, H.K.; Brown, S.; Niles, J.O.; Foley, J.A. Monitoring and estimating tropical forest carbon stocks: Making REDD a reality. Environ. Res. Lett. 2007, 2, 045023. [Google Scholar] [CrossRef]
- Zanne, A.E.; Lopez-Gonzalez, G.; Coomes, D.A.; Ilic, J.; Jansen, S.; Lewis, S.L.S.; Miller, R.B.; Swenson, N.G.; Wiemann, M.C.; Chave, J. Data from: Towards a Worldwide Wood Economics Spectrum. Available online: https://datadryad.org/bitstream/handle/10255/dryad.235/GlobalWoodDensityDatabase.xls?sequence=1 (accessed on 18 June 2019).
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef]
- Feldpausch, T.R.; Lloyd, J.; Lewis, S.L.; Brienen, R.J.W.; Gloor, M.; Monteagudo Mendoza, A.; Lopez-Gonzalez, G.; Banin, L.; Abu Salim, K.; Affum-Baffoe, K.; et al. Tree height integrated into pantropical forest biomass estimates. Biogeosciences 2012, 9, 3381–3403. [Google Scholar] [CrossRef] [Green Version]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Larjavaara, M.; Muller-Landau, H.C. Measuring tree height: A quantitative comparison of two common field methods in a moist tropical forest. Methods Ecol. Evol. 2013, 4, 793–801. [Google Scholar] [CrossRef]
- Slik, J.W.F.; Paoli, G.; McGuire, K.; Amaral, I.; Barroso, J.; Bastian, M.; Blanc, L.; Bongers, F.; Boundja, P.; Clark, C.; et al. Large trees drive forest aboveground biomass variation in moist lowland forests across the tropics. Glob. Ecol. Biogeogr. 2013, 22, 1261–1271. [Google Scholar] [CrossRef]
- Ploton, P.; Barbier, N.; Takoudjou Momo, S.; Réjou-Méchain, M.; Boyemba Bosela, F.; Chuyong, G.; Dauby, G.; Droissart, V.; Fayolle, A.; Goodman, R.C.; et al. Closing a gap in tropical forest biomass estimation: Taking crown mass variation into account in pantropical allometries. Biogeosciences 2016, 13, 1571–1585. [Google Scholar] [CrossRef]
- Meyer, V.; Saatchi, S.; Clark, D.B.; Keller, M.; Vincent, G.; Ferraz, A.; Espírito-Santo, F.; Oliveira, M.V.N.; Kaki, D.; Chave, J.; et al. Canopy Area of Large Trees Explains Aboveground Biomass Variations across Nine Neotropical Forest Landscapes. Biogeosci. Discuss. 2018, 1–38. [Google Scholar] [CrossRef]
- Calders, K.; Newnham, G.G.; Burt, A.; Murphy, S.; Raumonen, P.; Herold, M.; Culvenor, D.; Avitabile, V.; Disney, M.; Armston, J.; et al. Nondestructive estimates of above-ground biomass using terrestrial laser scanning. Methods Ecol. Evol. 2015, 6, 198–208. [Google Scholar] [CrossRef]
- Gonzalez de Tanago, J.; Lau, A.; Bartholomeus, H.; Herold, M.; Avitabile, V.; Raumonen, P.; Martius, C.; Goodman, R.C.; Disney, M.; Manuri, S.; et al. Estimation of above-ground biomass of large tropical trees with terrestrial LiDAR. Methods Ecol. Evol. 2018, 9, 223–234. [Google Scholar] [CrossRef]
- Clark, D.B.; Kellner, J.R. Tropical forest biomass estimation and the fallacy of misplaced concreteness. J. Veg. Sci. 2012, 23, 1191–1196. [Google Scholar] [CrossRef]
- Goodman, R.C.; Phillips, O.L.; Baker, T.R. Tightening up on tree carbon estimates. Nature 2012, 491, 527. [Google Scholar] [CrossRef]
- Sheil, D.; Eastaugh, C.S.; Vlam, M.; Zuidema, P.A.; Groenendijk, P.; van der Sleen, P.; Jay, A.; Vanclay, J. Does biomass growth increase in the largest trees? Flaws, fallacies and alternative analyses. Funct. Ecol. 2017, 31, 568–581. [Google Scholar] [CrossRef]
- Disney, M.I.; Boni Vicari, M.; Burt, A.; Calders, K.; Lewis, S.L.; Raumonen, P.; Wilkes, P. Weighing trees with lasers: Advances, challenges and opportunities. Interface Focus 2018, 8, 20170048. [Google Scholar] [CrossRef]
- Wilkes, P.; Lau, A.; Disney, M.; Calders, K.; Burt, A.; Gonzalez de Tanago, J.; Bartholomeus, H.; Brede, B.; Herold, M. Data acquisition considerations for Terrestrial Laser Scanning of forest plots. Remote Sens. Environ. 2017, 196, 140–153. [Google Scholar] [CrossRef]
- Malhi, Y.; Jackson, T.; Patrick Bentley, L.; Lau, A.; Shenkin, A.; Herold, M.; Calders, K.; Bartholomeus, H.; Disney, M.I. New perspectives on the ecology of tree structure and tree communities through terrestrial laser scanning. Interface Focus 2018, 8, 20170052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newnham, G.G.; Armston, J.D.; Calders, K.; Disney, M.I.; Lovell, J.L.; Schaaf, C.B.; Strahler, A.H.; Danson, F.M. Terrestrial Laser Scanning for Plot-Scale Forest Measurement. Curr. For. Rep. 2015, 1, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Momo Takoudjou, S.; Ploton, P.; Sonké, B.; Hackenberg, J.; Griffon, S.; de Coligny, F.; Kamdem, N.G.; Libalah, M.; Mofack, G.I.I.; Le Moguédec, G.; et al. Using terrestrial laser scanning data to estimate large tropical trees biomass and calibrate allometric models: A comparison with traditional destructive approach. Methods Ecol. Evol. 2018, 9, 905–916. [Google Scholar] [CrossRef]
- Abd Rahman, M.; Abu Bakar, M.; Razak, K.; Rasib, A.; Kanniah, K.; Wan Kadir, W.; Omar, H.; Faidi, A.; Kassim, A.; Abd Latif, Z. Non-Destructive, Laser-Based Individual Tree Aboveground Biomass Estimation in a Tropical Rainforest. Forests 2017, 8, 86. [Google Scholar] [CrossRef]
- Paynter, I.; Genest, D.; Peri, F.; Schaaf, C. Bounding uncertainty in volumetric geometric models for terrestrial lidar observations of ecosystems. Interface Focus 2018, 8, 20170043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stovall, A.E.; Anderson-Teixeira, K.J.; Shugart, H.H. Assessing terrestrial laser scanning for developing non-destructive biomass allometry. For. Ecol. Manag. 2018, 427, 217–229. [Google Scholar] [CrossRef]
- Burt, A.; Disney, M.; Raumonen, P.; Armston, J.; Calders, K.; Lewis, P. Rapid characterisation of forest structure from TLS and 3D modelling. In Proceedings of the 2013 IEEE International Geoscience and Remote Sensing Symposium—IGARSS, Melbourne, Australia, 21–26 July 2013; pp. 3387–3390. [Google Scholar] [CrossRef]
- Krooks, A.; Kaasalainen, S.; Kankare, V.; Joensuu, M.; Raumonen, P.; Kaasalainen, M. Tree structure vs. height from terrestrial laser scanning and quantitative structure models. Silva Fenn. 2014, 48, 1–11. [Google Scholar] [CrossRef]
- Holopainen, M.; Vastaranta, M.; Kankare, V. Biomass estimation of individual trees using stem and crown diameter TLS measurements. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2011, XXXVIII, 29–31. [Google Scholar] [CrossRef]
- Raumonen, P.; Kaasalainen, M.; Åkerblom, M.; Kaasalainen, S.; Kaartinen, H.; Vastaranta, M.; Holopainen, M.; Disney, M.; Lewis, P. Fast Automatic Precision Tree Models from Terrestrial Laser Scanner Data. Remote Sens. 2013, 5, 491–520. [Google Scholar] [CrossRef] [Green Version]
- Hackenberg, J.; Spiecker, H.; Calders, K.; Disney, M.; Raumonen, P. SimpleTree—An Efficient Open Source Tool to Build Tree Models from TLS Clouds. Forests 2015, 6, 4245–4294. [Google Scholar] [CrossRef]
- Kaasalainen, S.; Krooks, A.; Liski, J.; Raumonen, P.; Kaartinen, H.; Kaasalainen, M.; Puttonen, E.; Anttila, K.; Mäkipää, R. Change detection of tree biomass with terrestrial laser scanning and quantitative structure modelling. Remote Sens. 2014, 6, 3906–3922. [Google Scholar] [CrossRef]
- Åkerblom, M.; Raumonen, P.; Mäkipää, R.; Kaasalainen, M. Automatic tree species recognition with quantitative structure models. Remote Sens. Environ. 2017, 191, 1–12. [Google Scholar] [CrossRef]
- Guyana Lands and Surveys Commission. Guyana National Land Use Plan; Number June; Guyana Lands and Surveys Commission: Georgetown, TX, USA, 2013; p. 174.
- Muñoz, G.; Grieser, J. Climwat 2.0 for CROPWAT. Available online: http://www.fao.org/land-water/databases-and-software/climwat-for-cropwat/en/ (accessed on 18 June 2019).
- Phillips, O.; Baker, T.; Feldpausch, T.; Brienen, R.; Almeida, S.; Arroyo, L.; Aymard, G.; Chave, J.; Cardozo, N.D.; Chao, K.J.; et al. RAINFOR Field Manual for Plot Establishment and Remeasurement. 2009. Available online: http://www.rainfor.org/upload/ManualsEnglish/RAINFOR_field_manual_version_June_2009_ENG.pdf (accessed on 18 June 2019).
- Kitajima, K.; Mulkey, S.S.; Wright, S.J. Variation in Crown Light Utilization Characteristics among Tropical Canopy Trees. Ann. Bot. 2004, 95, 535–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, G.B.; Wiemann, M.C. Measuring wood specific gravity...correctly. Am. J. Bot. 2010, 97, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Calders, K.; Burt, A.; Origo, N.; Disney, M.; Nightingale, M.; Raumonen, P.; Akerblom, M.; Lewis, P. Realistic Forest Stand Reconstruction from Terrestrial LiDAR for Radiative Transfer Modelling. Remote Sens. 2018, 10, 933. [Google Scholar] [CrossRef]
- Vicari, M.B.; Disney, M.; Wilkes, P.; Burt, A.; Calders, K.; Woodgate, W. Leaf and wood classification framework for terrestrial LiDAR point clouds. Methods Ecol. Evol. 2019, 10, 680–694. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.I.K. A Concordance Correlation Coefficient to Evaluate Reproducibility. Biometrics 1989, 45, 255–268. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Réjou-Méchain, M.; Tanguy, A.; Piponiot, C.; Chave, J.; Hérault, B. Biomass: An R package for estimating above-ground biomass and its uncertainty in tropical forests. Method. Ecol. Evol. 2017, 8, 1163–1167. [Google Scholar] [CrossRef]
- Baskerville, G.L. Use of Logarithmic Regression in the Estimation of Plant Biomass. Can. J. For. Res. 1972, 2, 49–53. [Google Scholar] [CrossRef]
- Jucker, T.; Caspersen, J.; Chave, J.; Antin, C.; Barbier, N.; Bongers, F.; Dalponte, M.; van Ewijk, K.Y.; Forrester, D.I.; Haeni, M.; et al. Allometric equations for integrating remote sensing imagery into forest monitoring programmes. Glob. Chang. Biol. 2017, 23, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Roşca, S.; Suomalainen, J.; Bartholomeus, H.; Herold, M. Comparing terrestrial laser scanning and unmanned aerial vehicle structure from motion to assess top of canopy structure in tropical forests. Interface Focus 2018, 8, 20170038. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.; Keller, M.; Victoria, D.; Morton, D.C. Tree height and tropical forest biomass estimation. Biogeosciences 2013, 10, 8385–8399. [Google Scholar] [CrossRef] [Green Version]
- Brede, B.; Lau, A.; Bartholomeus, H.M.; Kooistra, L. Comparing RIEGL RiCOPTER UAV LiDAR derived canopy height and DBH with terrestrial LiDAR. Sensors 2017, 17, 2371. [Google Scholar] [CrossRef] [PubMed]
- Kuyah, S.; Dietz, J.; Muthuri, C.; Jamnadass, R.; Mwangi, P.; Coe, R.; Neufeldt, H. Allometric equations for estimating biomass in agricultural landscapes: I. Aboveground biomass. Agric. Ecosyst. Environ. 2012, 158, 216–224. [Google Scholar] [CrossRef]
- Oliveira, A.A.D.E.; Mori, S.A.; De Oliveira, A.A.; Mori, S.A. A central Amazonian terra firme forest. I. High tree species richness on poor soils. Biodivers. Conserv. 1999, 8, 1219–1244. [Google Scholar] [CrossRef]
- Lau, A.; Bentley, L.P.; Martius, C.; Shenkin, A.; Bartholomeus, H.; Raumonen, P.; Malhi, Y.; Jackson, T.; Herold, M. Quantifying branch architecture of tropical trees using terrestrial LiDAR and 3D modelling. Trees Struct. Funct. 2018, 32, 1219–1231. [Google Scholar] [CrossRef] [Green Version]
- Raumonen, P.; Casella, E.; Calders, K.; Murphy, S.; Åkerblom, M.; Kaasalainen, M. Massive-scale tree modelling from tls data. ISPRS Ann. Photogramm. Remote Sens. Spat. Inf. Sci. 2015, II-3/W4, 189–196. [Google Scholar] [CrossRef]
- Burt, A. New 3D Measurements of Forest Structure. Ph.D. Thesis, University College London, London, UK, 2017. [Google Scholar]
- Disney, M.I.; Burt, A.; Calders, K.; Schaaf, C.; Stovall, A. Innovations in ground and airborne technologies as reference and for training and validation: Terrestrial Laser Scanning (TLS). Surv. Geophys. 2019. [Google Scholar] [CrossRef]




| Model | Form AGB = |
|---|---|
| Ch05.II.3 | |
| Ch05.I.5 | |
| Ch14.H | |
| Ch14.E | |
| Rj17.E |
| Attributes | Allometric Model Dataset (n = 72) | Validation Dataset (n = 26) | ||
|---|---|---|---|---|
| Measuredpre | TLS-Derived | Measuredpost | TLS-Derived | |
| Diameter (cm) | 12.9 − 134.0 | 13.3 − 126.2 | 16.7 − 128.7 | 16.7 − 130.2 |
| Tree height (m) | 14 − 43.0 | 16.9 − 51.8 | 16.4 − 51.6 | 16.6 − 49.1 |
| Crown diameter (m) | 4.4 − 42.6 | 2.5 − 42.9 | 3.4 − 30.8 pre | 4.6 − 30.2 |
| WD (g cm) | 0.4 − 1.0 | 0.4 − 1.0 | 0.4 − 0.9 | 0.4 − 1.0 |
| AGB (Mg) | NA | 0.2 − 28.5 | 0.9 − 21.8 | 0.2 − 27.4 |
| Model | Type | Form | a | b | c | d | e | RDVC | df | RSE | adj-R | AICc |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| m1 | D | 0.6788 | 1.9337 | … | … | … | … | 70 | 0.360 | 0.90 | 61.52 | |
| m2 | D.WD | 0.6765 | 2.0246 | 1.0932 | … | … | −0.1968 | 69 | 0.274 | 0.94 | 23.61 | |
| m3 | D.WD.H | … | 1.9091 | 1.0978 | 0.3224 | … | −0.2138 | 69 | 0.266 | NA | 19.48 | |
| m4 | D.WD.H.CD | … | 1.7282 | 0.2603 | 1.1522 | 0.3698 | … | 68 | 0.240 | NA | 6.23 | |
| m5 | D.WD.CD | 0.5366 | 1.8124 | 1.1512 | … | 0.3878 | … | 68 | 0.246 | 0.96 | 9.28 |
| Model | Type | RMSE | CCC | Error (Mg) | Relative Error (%) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Sum | Mean | SD | Mean | SD | |||||
| m5 | D.WD.CD | 0.93 | 1.91 | 0.96 | 1.03 | 0.04 | 1.95 | 28.25 | 61.35 |
| m4 | D.WD.H.CD | 0.92 | 1.99 | 0.96 | 1.36 | 0.05 | 2.03 | 28.33 | 57.91 |
| Ch05.II.3 | WD.D.D.D | 0.89 | 2.32 | 0.94 | −7.49 | −0.29 | 2.35 | 5.54 | 48.26 |
| Ch05.I.5 | D.WD.H | 0.85 | 2.75 | 0.92 | −9.86 | −0.38 | 2.78 | 7.35 | 41.98 |
| Ch14.H | (D.WD.H) | 0.85 | 2.67 | 0.92 | −11.89 | −0.46 | 2.69 | 9.59 | 43.31 |
| m1 | D | 0.87 | 2.52 | 0.93 | 13.65 | 0.53 | 2.51 | 68.01 | 105.95 |
| Rj17.E | D.WD.E | 0.88 | 2.43 | 0.93 | −19.28 | −0.74 | 2.36 | −1.62 | 44.04 |
| Ch14.E | D.WD.E | 0.88 | 2.49 | 0.93 | −21.56 | −0.83 | 2.39 | −3.62 | 43.52 |
| m3 | D.WD.H | 0.88 | 2.92 | 0.89 | −32.11 | −1.23 | 2.69 | 14.80 | 60.97 |
| m2 | D.WD | 0.89 | 2.96 | 0.92 | 32.21 | 1.24 | 2.74 | 73.80 | 98.95 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lau, A.; Calders, K.; Bartholomeus, H.; Martius, C.; Raumonen, P.; Herold, M.; Vicari, M.; Sukhdeo, H.; Singh, J.; Goodman, R.C. Tree Biomass Equations from Terrestrial LiDAR: A Case Study in Guyana. Forests 2019, 10, 527. https://doi.org/10.3390/f10060527
Lau A, Calders K, Bartholomeus H, Martius C, Raumonen P, Herold M, Vicari M, Sukhdeo H, Singh J, Goodman RC. Tree Biomass Equations from Terrestrial LiDAR: A Case Study in Guyana. Forests. 2019; 10(6):527. https://doi.org/10.3390/f10060527
Chicago/Turabian StyleLau, Alvaro, Kim Calders, Harm Bartholomeus, Christopher Martius, Pasi Raumonen, Martin Herold, Matheus Vicari, Hansrajie Sukhdeo, Jeremy Singh, and Rosa C. Goodman. 2019. "Tree Biomass Equations from Terrestrial LiDAR: A Case Study in Guyana" Forests 10, no. 6: 527. https://doi.org/10.3390/f10060527