Genome-Wide Identification of WOX Gene Family and Expression Analysis during Rejuvenational Rhizogenesis in Walnut (Juglans regia L.)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Bioinformatics Analysis of WUSCHEL-Related Homeobox Genes in J. regia Genomes
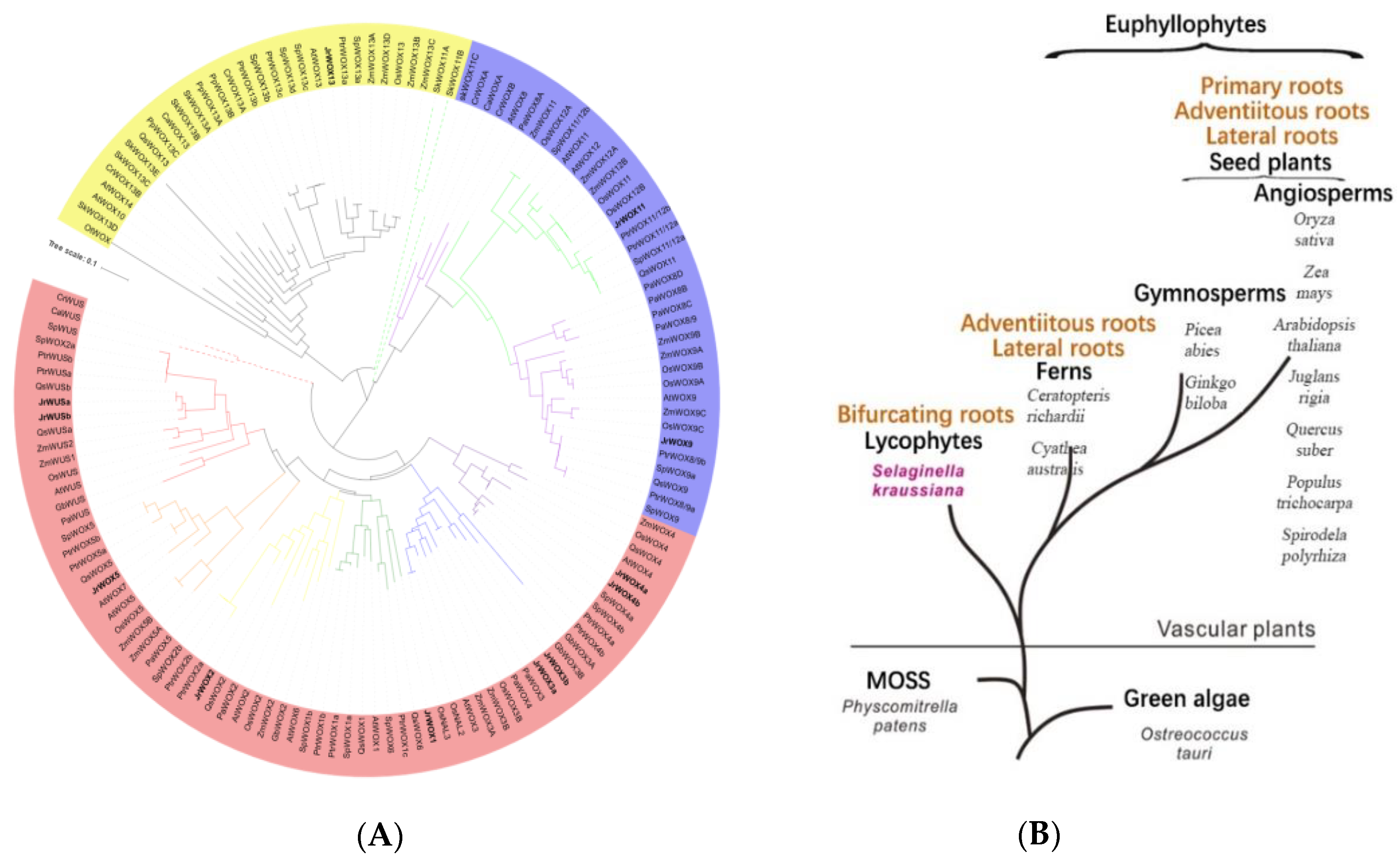
2.2. Phylogenetic Analysis
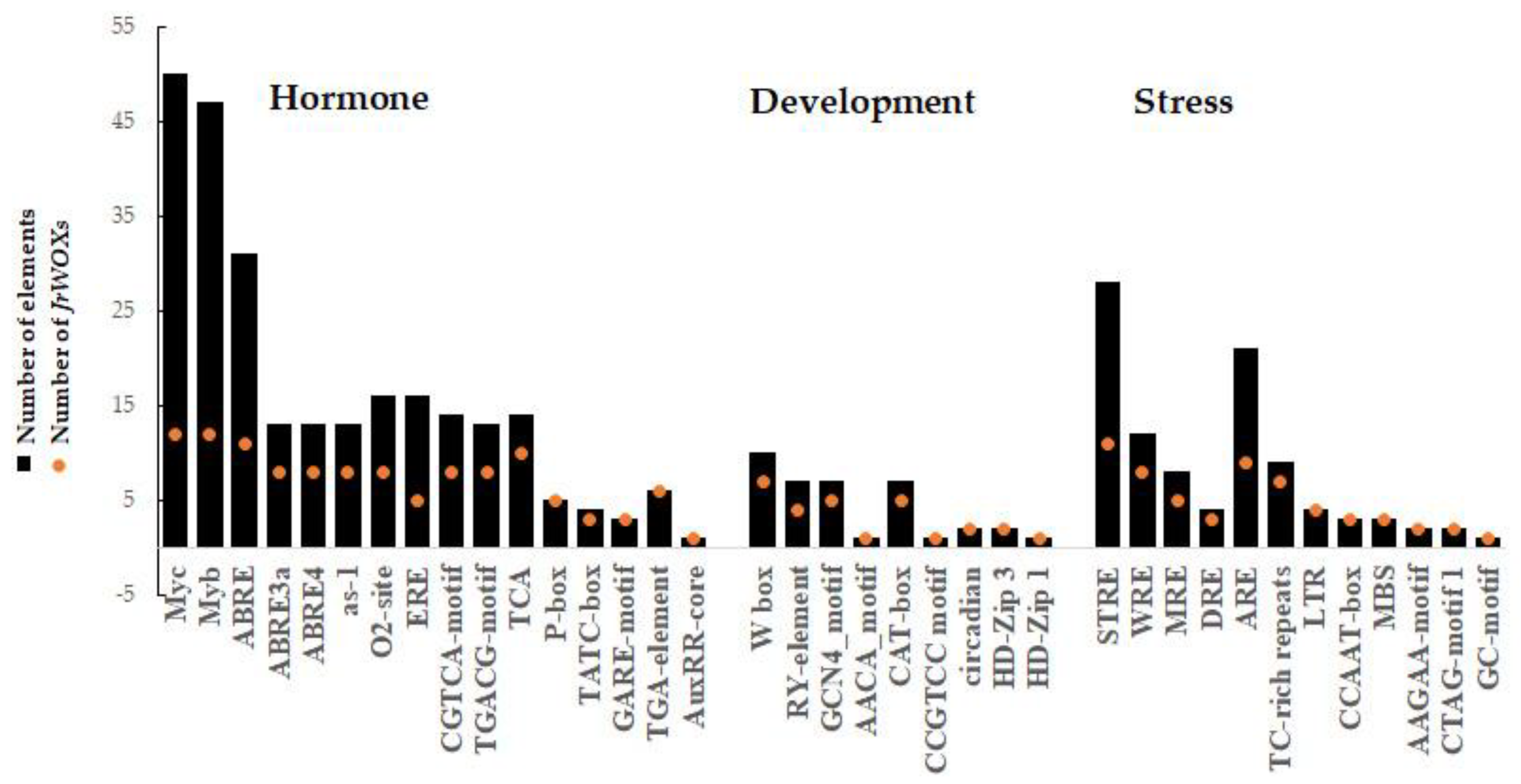
2.3. Analysis of Regulatory Elements in the Promoter Regions of the JrWOX Gene Family
2.4. Plant Materials, Growth Conditions, and Treatments
2.5. Expression Pattern Analysis of the JrWOX Genes
3. Results
3.1. Identification and Sequence Characteristics of JrWOX Genes
3.2. Gene Structure, Conservative Domain, and Motif Analysis of JrWOXs
3.3. Multiple Alignment the WOX Protein Sequence in J. regia
3.4. Characterization of Cis-Acting Elements in the Promoter Regions of JrWOXs
3.5. Phylogenetic Analysis of J. regia WOXs and Root Evolution in Green Plants
3.6. Rejuvenation-Changed Phenotype and Histological Features of Cuttings
3.7. Diversified Expression Patterns of JrWOX Genes
3.7.1. Expression Patterns of JrWOX Genes in Diversified Organs
3.7.2. Expression Patterns of JrWOX Genes in ARF of Mature and Rejuvenated Cuttings
4. Discussion
4.1. Diversification of WOX Genes in J. regia
4.2. The Characteristics of the WOX Gene Family of Walnut
4.3. The Promoter Regions of JrWOXs
4.4. The Evolution of JrWOX Genes and Root-Evolution Events
4.5. Expression Patterns of JrWOX Genes
4.6. Expression Profiling Revealed JrWOX Genes’ Responses to Rejuvenation and Involvement in ARF
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession No./Locus | Gene Name | Accession No. |
|---|---|---|---|
| Pa_WUS | JX512364 | Zm_WUS1 | AM234744 |
| Pa_WOX2 | AM286747 | Zm_WUS2 | AM234745 |
| Pa_WOX3 | JX411947 | Zm_WOX2 | AM490235 |
| Pa_WOX4 | JX411948 | Zm_WOX3B | AM491777 |
| Pa_WOX5 | JX411949 | Zm_WOX3A | AM490236 |
| Pa_WOX8/9 | GU944670 | Zm_WOX4 | AM490237 |
| Pa_WOX8A | JX411950 | Zm_WOX5A | AM490238 |
| Pa_WOX8B | JX411951 | Zm_WOX5B | AM490239 |
| Pa_WOX8C | JX411952 | Zm_WOX9A | AM490240 |
| Pa_WOX8D | JX411953 | Zm_WOX9B | AM490241 |
| At_WOX1 | AT3G18010 | Zm_WOX9C | AM490242 |
| At_WOX2 | AT5G59340 | Zm_WOX11 | EU954172 |
| At_WOX3 | AT2G28610 | Zm_WOX12A | AM234774 |
| At_WOX4 | AT1G46480 | Zm_WOX12B | AM234775 |
| At_WOX7 | AT5G05770 | Zm_WOX13A | AM234776 |
| At_WOX5 | AT3G11260 | Zm_WOX13B | AM234777 |
| At_WOX6 | AT2G01500 | Zm_WOX13C (ZmWOX14B) | EU952747 |
| At_WUS | AT2G17950 | Zm_WOX13D (ZmWOX14A) | EU961475 |
| At_WOX9 | AT2G33880 | Vv_WUS | AM447418 |
| At_WOX8 | AT5G45980 | Vv_WOX1 | AM439847 |
| At_WOX11 | AT3G03660 | Vv_WOX2 | AM488389 |
| At_WOX12 | AT5G17810 | Vv_WOX3 | AM429035 |
| At_WOX13 | AT4G35550 | Vv_WOX4 | AM447494 |
| At_WOX10 | AT1G20710 | Vv_WOX5 | AM454567 |
| At_WOX14 | AT1G20700 | Vv_WOX6 (VvWOX1B) | AM463144 |
| Gb_WUS | FM882128 | Vv_WOX9 | AM488026 |
| Gb_WOX2 | FM882124 | Vv_WOX11 | AM435207 |
| Gb_WOX3A | FM882125 | Vv_WOX13A | AM486367 |
| Gb_WOX3B | FM882126 | Vv_WOX13B | AM473516 |
| Pp_WOX13-LIKE A | AB699867 | Vv_WOX13C | AM463736 |
| Pp_WOX13-LIKE B | AB699868 | Sp_WOX4A | KF982703 |
| Pp_WOX13-LIKE C | XM_024511279 | Sp_WOX4B | KC691250 |
| Sm_WOX13 | XM_002962367 | Sp_WOX1B | KF982700 |
| Sm_WOX8 | XM_024668721 | Sp_WOX1A | KF982699 |
| Sm_WOX10 | XM_002981839 | Sp_WOX9A | KF982708 |
| Sm_WOX11 | XM_024684472 | Sp_WOX13B | KF982713 |
| Sm_WOX6 | XM_024673314 | Sp_WOX13C | AJ010810 |
| Sm_WOX5 | EFJ20992 | Sp_WOX13D(8) | AY479970 |
| Sm_WOX9 | EFJ33362 | Sp_WOX13A | KF982712 |
| Sm_WOX1 | EFJ33357 | Sp_WOX6 | KF982707 |
| Sm_WOX3 | EFJ33027 | Sp_WOX11/12B | KF982709 |
| Sm_WOX4 | EFJ21095 | Sp_WOX11/12A | KF982710 |
| Sm_WOX2 | EFJ35854 | Sp_WUSA | KF982698 |
| Ca_WUS | FR716459 | Sp_WOX2B | KF982702 |
| Ca_WOXA | FR716461 | Sp_WOX2A | FJ232064 |
| Ca_WOX13 | FR716460 | Sp_WOX5B | KF982706 |
| Cr_WUS | FR716458 | Sp_WOX9B | KF982709 |
| Cr_WOXA | FR716456 | PtrWUSa | Potri.005G114700 |
| Cr_WOXB | FR716457 | PtrWUSb | Potri.007G012100 |
| Cr_WOX13A | FR716454 | PtrWOX1a | Potri.012G047700 |
| Cr_WOX13B | FR716455 | PtrWOX1b | Potri.015G039100 |
| Os_WOX3a | LOC_Os11g01130.1 | PtrWOX1c | Potri.010G111400 |
| Os_WOX3b | LOC_Os12g01120.1 | PtrWOX2a | Potri.001G237900 |
| Os_WOX3c | LOC_Os11g01130.2 | PtrWOX2b | Potri.009G029200 |
| Os_WOX2 | LOC_Os05g02730.1 | PtrWOX4a | Potri.002G124100 |
| Os_WOX4 | LOC_Os04g55590.1 | PtrWOX4b | Potri.014G025300 |
| Os_WOX5 | LOC_Os01g62310.1 | PtrWOX5a | Potri.008G065400 |
| Os_WOX1 | LOC_Os04g56780.1 | PtrWOX5b | Potri.010G192100 |
| OsWOX9 | LOC_Os01g63510.1 | PtrWOX8/9a | Potri.004G051600 |
| OsWOX7 | LOC_Os01g47710.1 | PtrWOX8/9b | Potri.011G061400 |
| OsWOX12 | LOC_Os05g48990.1 | PtrWOX11/12a | Potri.013G066900 |
| OsWOX11 | LOC_Os07g48560.1 | PtrWOX11/12b | Potri.019G040800 |
| OsWOX6 | LOC_Os03g20910.1 | PtrWOX13a | Potri.005G101800. |
| OsWOX13 | LOC_Os07g34880.1 | PtrWOX13b | Potri.005G252800 |
| OsWOX10 | LOC_Os08g14400.1 | PtrWOX13c | Potri.002G008800 |
| OsWOX8 | LOC_Os01g60270.1 |
| Genes | Primer Sequence (F: 5′→3′) | Primer Sequence (F: 5′→3′) | Production/bp |
|---|---|---|---|
| GAPDH | ATGATGTCAAGGAAGGACTC | CACAATGATCTCAGCTCCG | 102 |
| JrWUSa | ACTCCTTCATCCAACGGCTT | TGAGCTCCAGGGAAGTACGA | 88 |
| JrWUSb | TCAATGGCTTCTGCAATGTTAG | AGAACCGCTCTTGCCACTGT | 104 |
| JrWOX1 | ACGAACACCAAAGTGCCCAT | ATCGGCTGGCTGTTTTCCT | 105 |
| JrWOX2 | CTCACGCTGGAATCCGACA | CAATGTGACCGTAAGCCCG | 128 |
| JrWOX3a | ACGGCTATGATGGGAATGCTA | AACAGGGAAGAGTTCAAGGGTC | 92 |
| JrWOX3b | CTGCTCTTCAACAACTTTCCCA | TTTGAGTTGTCTATCTCGCCCA | 157 |
| JrWOX4a | CAAGCAGGCTAACTCCGACA | GGGGAATCTCGTTTGTCGTC | 126 |
| JrWOX4b | AGCACGAACCCTCCCTCAC | AAAAGCCGCAACACCAGC | 106 |
| JrWOX5 | GACCCTCCAACTTTTCCCATT | AGTGCTCCATTTCTGTCCCAA | 122 |
| JrWOX9 | ACACAACTGCGGTCTCTCCCT | ACCCGATTCCCCAACACCT | 95 |
| JrWOX11 | GACGTAGTGTTGGTCCATTGCT | GCCCTCTTTCTTCCCCTTTAA | 135 |
| JrWOX13 | CAGGGCTGAAGATTTGTGCT | TACTGCCATCTGACGGGAAC | 107 |
References
- Bonga, J.M. Vegetative Propagation in Relation to Juvenility, Maturity, and Rejuvenation. In Tissue Culture in Forestry; Springer: Berlin/Heidelberg, Germany, 1982; pp. 387–412. [Google Scholar]
- Hackett, W.P.; Murray, J.R. Maturation and rejuvenation in woody species. In Micropropagation of Woody Plants; Springer: Berlin/Heidelberg, Germany, 1993; pp. 93–105. [Google Scholar]
- Wendling, I.; Trueman, S.J.; Xavier, A. Maturation and related aspects in clonal forestry—Part II: Reinvigoration, rejuvenation and juvenility maintenance. New For. 2014, 45, 473–486. [Google Scholar] [CrossRef]
- Von Aderkas, P.; Bonga, J.M. Influencing micropropagation and somatic embryogenesis in mature trees by manipulation of phase change, stress and culture environment. Tree Physiol. 2000, 20, 921–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Gao, Y.; Song, X.B.; Ma, Q.G.; Zhang, J.P.; Pei, D. A novel rejuvenation approach to induce endohormones and improve rhizogenesis in mature Juglans tree. Plant Methods 2018, 14, 13. [Google Scholar] [CrossRef] [PubMed]
- Suklabaidya, A.; Mehta, K. Rejuvenation of Senile Horticultural Plantations for Improved Productivity and Quality. Int. J. Hortic. Agric. Food Sci. 2019, 3, 4. [Google Scholar] [CrossRef]
- Chang, F.; Chen, P.-J.; Shen, C.-H.; Hsieh, T.-J.; Hsu, Y.-W.; Huang, B.-L.; Kuo, C.-I.; Chen, Y.-T.; Chu, H.-A.; Yeh, K.-W.; et al. Proteomic profiling of proteins associated with the rejuvenation of Sequoia sempervirens (D. Don) Endl. Proteome Sci. 2010, 8, 64. [Google Scholar] [CrossRef] [Green Version]
- Van Der Graaff, E.; Laux, T.; Rensing, S.A. The WUS homeobox-containing (WOX) protein family. Genome Biol. 2009, 10, 248. [Google Scholar] [CrossRef]
- Liu, B.; Wang, L.; Zhang, J.; Li, J.B.; Zheng, H.Q.; Chen, J.; Lu, M.Z. WUSCHEL-related Homeobox genes in Populus tomentosa: Diversified expression patterns and a functional similarity in adventitious root formation. BMC Genom. 2014, 15, 296. [Google Scholar] [CrossRef] [Green Version]
- Lian, G.; Ding, Z.W.; Wang, Q.; Zhang, D.B.; Xu, J. Origins and evolution of WUSCHEL-related homeobox protein family in plant kingdom. Sci. World J. 2014, 2014, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Yves, D.; Claire, T.N.; Gaelle, C.; Vincent, T.; Halima, M.; Patrick, L.; Hervé, M.; Martin, K.; Alain, L. Genes of the most conserved WOX clade in plants affect root and flower development in Arabidopsis. BMC Evol. Biol. 2008, 8, 291. [Google Scholar]
- Zhang, X.; Zong, J.; Liu, J.H.; Yin, J.Y.; Zhang, D.B. Genome-Wide Analysis of WOX Gene Family in Rice, Sorghum, Maize, Arabidopsis and Poplar. J. Integr. Plant Biol. 2010, 52, 1016–1026. [Google Scholar] [CrossRef]
- Haecker, A.; Groß-hardt, R.; Geiges, B.; Sarkar, A.; Breuninger, H.; Herrmann, M.; Laux, T. Expression dynamics of WOX genes mark cell fate decisions during early embryonic patterning in Arabidopsis thaliana. Development 2004, 131, 657–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoof, H.; Lenhard, M.; Haecker, A.; Mayer, K.F.X.; Laux, T. The Stem Cell Population of Arabidopsis Shoot Meristems is maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 2000, 100, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Luijten, M.; Miyashima, S.; Lenhard, M.; Hashimoto, T.; Nakajima, K.; Scheres, B.; Heidstra, R.; Laux, T.; Sarkar, A.K. Conserved factors regulate signalling in Arabidopsis thaliana shoot and root stem cell organizers. Nature 2007, 446, 811–814. [Google Scholar]
- Stahl, Y.; Wink, R.H.; Ingram, G.C.; Simon, R. A signaling module controlling the stem cell niche in Arabidopsis root meristems. Curr. Biol. 2009, 19, 909–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirakawa, Y.; Kondo, Y.; Fukuda, H. TDIF Peptide Signaling Regulates Vascular Stem Cell Proliferation via the WOX4 Homeobox Gene in Arabidopsis. Plant Cell 2010, 22, 2618–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, R.; Ji, J.; Kelsey, E.; Ohtsu, K.; Schnable, P.S.; Scanlon, M.J. Tissue Specificity and Evolution of Meristematic WOX3 Function. Plant Physiol. 2009, 149, 841–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breuninger, H.; Rikirsch, E.; Hermann, M.; Ueda, M.; Laux, T. Differential expression of WOX genes mediates apical-basal axis formation in the Arabidopsis embryo. Dev. Cell 2008, 14, 867–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.O.; Zheng, Z.; Oppenheimer, D.G.; Hauser, B.A. The PRETTY FEW SEEDS2 gene encodes an Arabidopsis homeodomain protein that regulates ovule development. Development 2005, 132, 841–849. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Chory, J.; Weigel, D. Combinations of WOX activities regulate tissue proliferation during Arabidopsis embryonic development. Dev. Biol. 2007, 309, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Sheng, L.; Xu, Y.; Li, J.; Yang, Z.; Huang, H.; Xu, L. WOX11 and 12 Are Involved in the First-Step Cell Fate Transition during de Novo Root Organogenesis in Arabidopsis. Plant Cell 2014, 26, 1081–1093. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Hu, X.; Qin, P.; Prasad, K.; Hu, Y.; Xu, L. The WOX11—LBD16 Pathway Promotes Pluripotency Acquisition in Callus Cells during de Novo Shoot Regeneration in Tissue Culture. Plant Cell Physiol. 2018, 59, 734–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, H.; Jia, Y.; Niu, T.; Yu, Q.; Ding, Z. The key players of the primary root growth and development also function in lateral roots inArabidopsis. Plant Cell Rep. 2014, 33, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Hu, Y.; Dai, M.; Huang, L.; Zhou, D.-X. The WUSCHEL-Related Homeobox Gene WOX11 Is Required to Activate Shoot-Borne Crown Root Development in Rice. Plant Cell Online 2009, 21, 736–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Cheng, S.; Song, Y.; Huang, Y.; Zhou, S.; Liu, X.; Zhou, D.-X. The Interaction between Rice ERF3 and WOX11 Promotes Crown Root Development by Regulating Gene Expression Involved in Cytokinin Signaling. Plant Cell 2015, 27, 2469–2483. [Google Scholar] [CrossRef]
- Jiang, W.; Zhou, S.; Zhang, Q.; Song, H.; Zhou, D.-X.; Zhao, Y. Transcriptional regulatory network of WOX11 is involved in the control of crown root development, cytokinin signals, and redox in rice. J. Exp. Bot. 2017, 68, 2787–2798. [Google Scholar] [CrossRef]
- Xu, M.; Xie, W.F.; Huang, M.R. Two WUSCHEL-related HOMEOBOX genes, PeWOX11a and PeWOX11b, are involved in adventitious root formation of poplar. Physiol. Plant. 2015, 155, 446–456. [Google Scholar] [CrossRef]
- Stevens, M.E.; Woeste, K.E.; Pijut, P.M. Localized gene expression changes during adventitious root formation in black walnut (Juglans nigra L.). Tree Physiol. 2018, 38, 877–894. [Google Scholar] [CrossRef] [Green Version]
- Song, X.B.; Xi, S.K.; Zhang, J.P.; Ma, Q.G.; Zhou, Y.; Xu, H.; Zhang, J.W.; Pei, D. ‘ZhongNingsheng’: A New Distant Hybrid Cultivar of Walnut. Hortscience 2019, 54, 2257–2259. [Google Scholar] [CrossRef] [Green Version]
- Pei, D.; Gu, R.S. Shoot Rooting in vitro for Walnut Cultivars. Sci. Silvae Sin. 2002, 38, 32–37. [Google Scholar]
- Pei, D.; Zhang, J.P.; Xu, H.Z. Cutting Propagation Method for Juglans: China. CN 200410009201 2005, 11 June 2004. [Google Scholar]
- Ma, Q.; Zhang, J.; Pei, D. Genetic Analysis of Walnut Cultivars in China Using Fluorescent Amplified Fragment Length Polymorphism. J. Am. Soc. Hortic. Sci. 2019, 136, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Song, X.B.; Sun, L.; Luo, H.T.; Ma, Q.G.; Zhao, Y.; Pei, D. Genome-wide identification and characterization of long non-coding RNAs from mulberry (Morus notabilis) RNA-Seq data. Genes 2016, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.B.; Chang, Y.Y.; Liu, H.; Xu, H.M.; Pei, D. Reference Gene Selection and Genes Expression Analysis During Adventitious Root Formation in Walnut. Acta Hortic. Sin. 2019, 46, 1907–1918. [Google Scholar] [CrossRef]
- Martínezgarcía, P.J.; Crepeau, M.W.; Puiu, D.; Gonzalezibeas, D.; Whalen, J.; Stevens, K.A.; Paul, R.; Butterfield, T.S.; Britton, M.T.; Reagan, R.L. The walnut (Juglans regia) genome sequence reveals diversity in genes coding for the biosynthesis of non-structural polyphenols. Plant J. Cell Mol. Biol. 2016, 87, 507–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mark, J.; Irena, Z.; Yan, R.; Yuri, M.; Scott, M.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; Mcwilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Hageleit, M. Validities of mRNA quantification using recombinant RNA and recombinant DNA external calibration curves in real-time RT-PCR. Biotechnol. Lett. 2001, 23, 275–282. [Google Scholar] [CrossRef]
- Mukherjee, K.; Brocchieri, L.; Burglin, T.R. A Comprehensive Classification and Evolutionary Analysis of Plant Homeobox Genes. Mol. Biol. Evol. 2009, 26, 2775–2794. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Xu, L. Recruitment of IC- WOX Genes in Root Evolution. Trends Plant Sci. 2018, 23, 490–496. [Google Scholar] [CrossRef]
- Zeng, M.; Hu, B.; Li, J.; Zhang, G.; Xu, L. Stem cell lineage in body layer specialization and vascular patterning of rice root and leaf. Sci. Bull. 2016, 61, 847–858. [Google Scholar] [CrossRef]
- Li, J.; Zhang, J.; Jia, H.; Liu, B.; Sun, P.; Hu, J.; Wang, L.; Lu, M. The WUSCHEL-related homeobox 5a (PtoWOX5a) is involved in adventitious root development in poplar. Tree Physiol. 2017, 38, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bi, C.; Xu, Y.; Ye, Q.; Yin, T.; Ye, N. Genome-wide identification and characterization of WRKY gene family in Salix suchowensis. PeerJ 2016, 4, e2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Niu, L.; McHale, N.A.; Ohme-Takagi, M.; Mysore, K.S.; Tadege, M. Evolutionarily conserved repressive activity of WOX proteins mediates leaf blade outgrowth and floral organ development in plants. Proc. Natl. Acad. Sci. USA 2013, 110, 366–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romerabranchat, M.; Ripoll, J.J.; Yanofsky, M.F.; Pelaz, S. The WOX13 homeobox gene promotes replum formation in the Arabidopsis thaliana fruit. Plant J. 2013, 73, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Qu, Y.; Sheng, L.; Liu, J.; Huang, H.; Xu, L. A simple method suitable to study de novo root organogenesis. Front. Plant Sci. 2014, 5, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, D.; Hao, Y.; Cui, H. The WUSCHEL Related Homeobox Protein WOX7 Regulates the Sugar Response of Lateral Root Development in Arabidopsis thaliana. Mol. Plant. 2016, 9, 261–270. [Google Scholar] [CrossRef] [Green Version]
- Nardmann, J.; Werr, W. The invention of WUS-like stem cell-promoting functions in plants predates leptosporangiate ferns. Plant Mol. Biol. 2012, 78, 123–134. [Google Scholar] [CrossRef]
- Rodriguez, K.; Perales, M.; Snipes, S.; Yadav, R.K.; Diaz-Mendoza, M.; Reddy, G.V. DNA-dependent homodimerization, sub-cellular partitioning, and protein destabilization control WUSCHEL levels and spatial patterning. Proc. Natl. Acad. Sci. USA 2016, 113, E6307–E6315. [Google Scholar] [CrossRef] [Green Version]
- Lu, G.; Bruce, W.B. A novel cis-acting element conferring root-preferred gene expression in maize. J. Plant Physiol. 2000, 156, 277–283. [Google Scholar] [CrossRef]
- Li, Z. Functional Charaterization of a PtoWOX11/12a gene in Stress Resistance of Populus tomentosa. Ph.D. Thesis, Chinese Academy of Forestry, Beijing, China, 2017. [Google Scholar]
- Caboni, E.; Tonelli, M.G.; Lauri, P.; Iacovacci, P.; Kevers, C.; Damiano, C.; Gaspar, T. Biochemical aspects of almond microcuttings related to in vitro rooting ability. Biol. Plant. 1997, 39, 91–97. [Google Scholar] [CrossRef]
- Berkefeld, H. Pyk10, a seedling and root specific gene and promoter from Arabidopsis thaliana. Plant Sci. 2001, 161, 337–346. [Google Scholar]
- He, P.; Zhang, Y.; Liu, H.; Yuan, Y.; Wang, C.; Yu, J.; Xiao, G. Comprehensive analysis of WOX genes uncovers that WOX13 is involved in phytohormone-mediated fiber development in cotton. BMC Plant Biol. 2019, 19, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amissah, J.N.; Paolillo, D.J.; Bassuk, N. Adventitious root formation in stem cuttings of Quercus bicolor and Quercus macrocarpa and its relationship to stem anatomy. J. Am. Soc. Hortic. Sci. 2008, 133, 479–486. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, J.; Yang, Z.; Matsui, A.; Seki, M.; Li, S.; Yan, X.; Kohnen, M.V.; Gu, L.; Prasad, K.; et al. PtWOX11 acts as master regulator conducting the expression of key transcription factors to induce de novo shoot organogenesis in poplar. Plant Mol. Biol. 2018, 98, 389–406. [Google Scholar] [CrossRef]
- Edwards, R.A.; Thomas, M.B. Observations on physical barriers to root formation in cuttings. Plant Propagator 1980, 26, 6–8. [Google Scholar]
- Amissah, J.N.; Bassuk, N. Cutting back stock plants promotes adventitious rooting of stems of Quercus bicolor and Quercus macrocarpa. J. Environ. Hortic. 2009, 73, 159–165. [Google Scholar]
- Kucukoglu, M.; Nilsson, J.; Zheng, B.; Chaabouni, S.; Nilsson, O. WUSCHEL-RELATED HOMEOBOX4 (WOX4)-like genes regulate cambial cell division activity and secondary growth in Populus trees. New Phytol. 2017, 215, 642–657. [Google Scholar] [CrossRef] [Green Version]
- Druege, U.; Franken, P.; Hajirezaei, M.R. Plant Hormone Homeostasis, Signaling, and Function during Adventitious Root Formation in Cuttings. Front. Plant Sci. 2016, 7, 381. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.H.; Paek, N.C. Regulatory role of the OsWOX3A transcription factor in rice root development. Plant Signal. Behav. 2016, 11, e1184807. [Google Scholar] [CrossRef] [Green Version]
- Pi, L.; Aichinger, E.; van der Graaff, E.; Llavata-Peris, C.I.; Weijers, D.; Hennig, L.; Groot, E.; Laux, T. Organizer-Derived WOX5 Signal Maintains Root Columella Stem Cells through Chromatin-Mediated Repression of CDF4 Expression. Dev. Cell 2015, 33, 576–588. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Feng, H.; Hu, Q.; Qu, H.; Chen, A.; Yu, L.; Xu, G. Improving rice tolerance to potassium deficiency by enhancing OsHAK16p: WOX11-controlled root development. Plant Biotechnol. J. 2015, 13, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Chen, N.; Zhao, M.; Wang, Y.; He, R.; Peng, X.; Shen, S. Identification and functional divergence analysis of WOX gene family in paper mulberry. Int. J. Mol. Sci. 2017, 18, 1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]







| Gene | Gene ID | Protein Length/aa | Molecular Weight (Mw)/Da | Isoelectric Point (pI) | Protein Hydrophobi-city | Aliphatic Amino Acid Index | Subcellular Localization |
|---|---|---|---|---|---|---|---|
| JrWUSa | XM_018977489.1 | 285 | 31,531.34 | 5.70 | −0.966 | 43.79 | Nucleus |
| JrWUSb | XM_018974029.1 | 283 | 31,356.19 | 6.15 | −0.925 | 44.77 | Nucleus |
| JrWOX1 | XM_018987154.1 | 360 | 41,092.06 | 9.43 | −0.891 | 58.00 | Nucleus |
| JrWOX2 | XM_018980445.1 | 251 | 27,979.82 | 6.40 | −0.726 | 53.63 | Nucleus |
| JrWOX3a | XM_018969027.1 | 204 | 23,370.43 | 8.77 | −0.590 | 61.72 | Nucleus |
| JrWOX3b | XM_019004669.1 | 206 | 23,981.82 | 6.15 | −0.879 | 62.96 | Nucleus |
| JrWOX4a | XM_018964283.1 | 226 | 25,844.20 | 8.92 | −1.000 | 54.87 | Nucleus |
| JrWOX4b | XM_019000128.1 | 226 | 25,541.73 | 9.16 | −0.908 | 54.82 | Nucleus |
| JrWOX5 | XM_018966208.1 | 184 | 20,891.56 | 8.73 | −0.708 | 65.16 | Nucleus |
| JrWOX9 | XM_018977986.1 | 387 | 42,711.37 | 6.62 | −0.597 | 58.71 | Nucleus |
| JrWOX11 | XM_018977839.1 | 258 | 28,171.61 | 5.86 | −0.434 | 71.36 | Nucleus |
| JrWOX13 | XM_019002154.1 | 275 | 31,410.80 | 5.61 | −0.923 | 65.56 | Nucleus |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, Y.; Song, X.; Zhang, Q.; Liu, H.; Bai, Y.; Lei, X.; Pei, D. Genome-Wide Identification of WOX Gene Family and Expression Analysis during Rejuvenational Rhizogenesis in Walnut (Juglans regia L.). Forests 2020, 11, 16. https://doi.org/10.3390/f11010016
Chang Y, Song X, Zhang Q, Liu H, Bai Y, Lei X, Pei D. Genome-Wide Identification of WOX Gene Family and Expression Analysis during Rejuvenational Rhizogenesis in Walnut (Juglans regia L.). Forests. 2020; 11(1):16. https://doi.org/10.3390/f11010016
Chicago/Turabian StyleChang, Yingying, Xiaobo Song, Qixiang Zhang, Hao Liu, Yongchao Bai, Xiashuo Lei, and Dong Pei. 2020. "Genome-Wide Identification of WOX Gene Family and Expression Analysis during Rejuvenational Rhizogenesis in Walnut (Juglans regia L.)" Forests 11, no. 1: 16. https://doi.org/10.3390/f11010016
APA StyleChang, Y., Song, X., Zhang, Q., Liu, H., Bai, Y., Lei, X., & Pei, D. (2020). Genome-Wide Identification of WOX Gene Family and Expression Analysis during Rejuvenational Rhizogenesis in Walnut (Juglans regia L.). Forests, 11(1), 16. https://doi.org/10.3390/f11010016