Needle Senescence Affects Fire Behavior in Aleppo Pine (Pinus halepensis Mill.) Stands: A Simulation Study

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Senescence Scenarios
2.2. Stand Structures and Fuel Features
2.3. Fire Behavior Modelization
2.4. Dead Mass Fraction Sensitivity Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barbero, M.; Loisel, R.; Quézel, P.; Romane, F.; Richardson, D.M. Pines of the Mediterranean Basin. In Ecology and Biogeography of Pinus; Cambridge University Press: Cambridge, UK, 1998; ISBN 9780521789103. [Google Scholar]
- Mauri, A.; Di Leo, M.; de Rigo, D.; Caudullo, G. Pinus halepensis and Pinus brutia in Europe: Distribution, habitat, usage and threats. Eur. Atlas For. Tree Species 2016, 1, 122–123. [Google Scholar]
- Keeley, J.E. Ecology and evolution of pine life histories. Ann. For. Sci. 2012, 69, 445–453. [Google Scholar] [CrossRef] [Green Version]
- Resco de Dios, V.; Arteaga, C.; Hedo, J.; Gil-Pelegrín, E.; Voltas, J. A trade-off between embolism resistance and bark thickness in conifers: Are drought and fire adaptations antagonistic? Plant Ecol. Divers. 2018, 11, 253–258. [Google Scholar] [CrossRef]
- Palmero-Iniesta, M.; Domènech, R.; Molina-Terrén, D.; Espelta, J.M. Fire behavior in Pinus halepensis thickets: Effects of thinning and woody debris decomposition in two rainfall scenarios. For. Ecol. Manag. 2017, 404, 230–240. [Google Scholar] [CrossRef]
- Quézel, P. Taxonomy and biogeography of Mediterranean pines (Pinus halepensis and P. brutia). In Ecology, Biogeography and Management of Pinus halepensis and Pinus brutia Forest Ecosystems in the Mediterranean Basin; Néeman, G., Trabaud, L., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2000; pp. 1–12. [Google Scholar]
- Van Wagner, C.E. Conditions for the start and spread of crown fire. Can. J. For. Res. 1977, 7, 23–34. [Google Scholar] [CrossRef]
- Cruz, M.G.; Alexander, M.E. Modelling the rate of fire spread and uncertainty associated with the onset and propagation of crown fires in conifer forest stands. Int. J. Wildl. Fire 2017, 26, 413–426. [Google Scholar] [CrossRef]
- Dimitrakopoulos, A.P.; Mitsopoulos, I.D.; Raptis, D.I. Nomographs for predicting crown fire initiation in Aleppo pine (Pinus halepensis Mill.) forests. Eur. J. For. Res. 2007, 126, 555–561. [Google Scholar] [CrossRef]
- Hoffman, C.M.; Canfield, J.; Linn, R.R.; Mell, W.; Sieg, C.H.; Pimont, F.; Ziegler, J. Evaluating Crown Fire Rate of Spread Predictions from Physics-Based Models. Fire Technol. 2016, 52, 221–237. [Google Scholar] [CrossRef] [Green Version]
- Rothermell, R.C. A Mathematical Model for Predicting Fire Spread; US Department of Agriculture: Washington, DC, USA, 1972.
- Rothermel, R.C. Predicting Behavior and Size of Crown Fires in the Northern Rocky Mountains; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Washington, DC, USA, 1991; Res. Pap.; Volume 438, p. 46.
- Van Wagner, C.E. Prediction of crown fire behavior in two stands of jack pine. Can. J. For. Res. 1993, 23, 442–449. [Google Scholar] [CrossRef]
- Alexander, M.E.; Cruz, M.G. Crown Fire Dynamics in Conifer Forests; USDA Forest Service: Washington, DC, USA, 2016; Gen. Tech. Rep. PNW-GTR; Volume 2016, pp. 163–258.
- Andrews, P.L. Current status and future needs of the BehavePlus Fire Modeling System. Int. J. Wildl. Fire 2014, 23, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Finney, M.A. An overview of FlamMap fire modeling capabilities. In Proceedings of the Fuels Management-How to Measure Success: Conference Proceedings, Portland, OR, USA, 28–30 March 2006; pp. 213–220. [Google Scholar]
- Scott, J.H.; Reinhardt, E.D. Assessing Crown Fire Potential by Linking Models of Surface and Crown Fire Behavior; USDA Forest Service: Washington, DC, USA, 2001; Res. Pap. RMRS-RP; pp. 1–62. [CrossRef] [Green Version]
- Keane, R.E. Wildland Fuel Fundamentals and Applications; Springer: Cham, Switzerland, 2015; ISBN 9783319090153. [Google Scholar]
- Resco de Dios, V. Plant-Fire Interactions; von Gadow, K., Pukkala, T., Tomé, M., Managing, F., Eds.; Springer: Cham, Switzerland, 2020; ISBN 9783030411916. [Google Scholar]
- Nolan, R.H.; Blackman, C.J.; De Dios, R.; Choat, B.; Medlyn, B.E.; Li, X.; Bradstock, R.A.; Boer, M.M. Linking Forest Flammability and Plant Vulnerability to Drought. Forests 2020, 11, 779. [Google Scholar] [CrossRef]
- López-Santalla, A.; López-Garcia, M. Los Incendios Forestales en España. Decenio 2006–2015; Ministerio de Agricultura Pesca y Alimentación, Secretaria General Técnica de Impresión: Madrid, Spain, 2019; p. 166. [Google Scholar]
- Karavani, A.; Boer, M.M.; Baudena, M.; Colinas, C.; Díaz-Sierra, R.; Pemán, J.; de Luis, M.; Enríquez-de-Salamanca, Á.; Resco de Dios, V. Fire-induced deforestation in drought-prone Mediterranean forests: Drivers and unknowns from leaves to communities. Ecol. Monogr. 2018, 88, 141–169. [Google Scholar] [CrossRef] [Green Version]
- Alexander, M.E.; Cruz, M.G. Assessing the effect of foliar moisture on the spread rate of crown fires. Int. J. Wildl. Fire 2013, 22, 415–427. [Google Scholar] [CrossRef]
- Jenkins, M.J.; Page, W.G.; Hebertson, E.G.; Alexander, M.E. Fuels and fire behavior dynamics in bark beetle-attacked forests in Western North America and implications for fire management. For. Ecol. Manag. 2012, 275, 23–34. [Google Scholar] [CrossRef]
- Talucci, A.C.; Krawchuk, M.A. Dead forests burning: The influence of beetle outbreaks on fire severity and legacy structure in sub-boreal forests. Ecosphere 2019, 10, e02744. [Google Scholar] [CrossRef] [Green Version]
- Reiner, A.L. Fire Behavior in Beetle-killed Stands: A Brief Review of Literature Focusing on Early Stages after Beetle Attack; US Forest Service Pacific Southwest Region: Vallejo, CA, USA, 2017; Volume 5, pp. 1–5.
- Nolan, R.H.; Hedo, J.; Arteaga, C.; Sugai, T.; Resco de Dios, V. Physiological drought responses improve predictions of live fuel moisture dynamics in a Mediterranean forest. Agric. For. Meteorol. 2018, 263, 417–427. [Google Scholar] [CrossRef] [Green Version]
- Hover, A.; Buissart, F.; Caraglio, Y.; Heinz, C.; Pailler, F.; Ramel, M.; Vennetier, M.; Prévosto, B.; Sabatier, S. Growth phenology in Pinus halepensis Mill: Apical shoot bud content and shoot elongation. Ann. For. Sci. 2017, 74. [Google Scholar] [CrossRef]
- Rossa, C.G.; Fernandes, P.M. On the effect of live fuel moisture content on fire rate of spread. For. Syst. 2017, 26. [Google Scholar] [CrossRef] [Green Version]
- Generalitat Valenciana; Servei de Prevenció D’Incendis Forestals Clave para la identificación de los modelos de combustible de la comunitat valenciana. Conselleria de Agricultura, Desarrollo Rural, Emergencia Climática y Transición Ecológica. 2020, 1, 1–38.
- Scott, J.H.; Burgan, R.E. Standard Fire Behavior Fuel Models: A Comprehensive Set for Use with Rothermel’s Surface Fire Spread Model; USDA Forest Service: Washington, DC, USA, 2005; Gen. Tech. Rep. RMRS-GTR; pp. 1–76. [CrossRef]
- Boer, M.M.; Nolan, R.H.; De Dios, V.R.; Clarke, H.; Owen, F.; Bradstock, R.A. Changing Weather Extremes Call for Early Warning of Potential for Catastrophic Fire. Earths Future. 2017. [Google Scholar] [CrossRef]
- Jervis, F.X.; Rein, G. Experimental study on the burning behaviour of Pinus halepensis needles using small-scale fire calorimetry of live, aged and dead samples. FIRE Mater. 2016, 40, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Soriano Sanchez, J.; Quilez Moraga, R. Análisis de la humedad del combustible vivo en la Comunitat Valenciana 1/13. Soc. Española Cienc. For. 2017, 1–14. [Google Scholar]
- Mitsopoulos, I.D.; Dimitrakopoulos, A.P. Canopy fuel characteristics and potential crown fire behavior in Aleppo pine (Pinus halepensis Mill.) forests. Ann. For. Sci. 2007, 64, 287–299. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R Development Core Team, R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; ISBN 3900051070. [Google Scholar]
- Alvarez, A.; Gracia, M.; Retana, J. Fuel types and crown fire potential in Pinus halepensis forests. Eur. J. For. Res. 2012, 131, 463–474. [Google Scholar] [CrossRef]
- Keeley, J.E.; Pfaff, A.H.; Safford, H.D. Fire suppression impacts on postfire recovery of Sierra Nevada chaparral shrublands. Int. J. Wildl. Fire 2005. [Google Scholar] [CrossRef] [Green Version]
- Pimont, F.; Ruffault, J.; Martin-StPaul, N.K.; Dupuy, J.L. Why is the effect of live fuel moisture content on fire rate of spread underestimated in field experiments in shrublands? Int. J. Wildl. Fire 2019, 28, 127–137. [Google Scholar] [CrossRef]
- Dennison, P.E.; Moritz, M.A.; Taylor, R.S. Evaluating predictive models of critical live fuel moisture in the Santa Monica Mountains, California. Int. J. Wildl. Fire 2008, 17, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Nolan, R.H.; Boer, M.M.; Resco De Dios, V.; Caccamo, G.; Bradstock, R.A. Large-scale, dynamic transformations in fuel moisture drive wildfire activity across southeastern Australia. Geophys. Res. Lett. 2016, 43, 4229–4238. [Google Scholar] [CrossRef] [Green Version]
- Luo, K.; Quan, X.; He, B.; Yebra, M. Effects of live fuel moisture content on wildfire occurrence in fire-prone regions over southwest China. Forests 2019, 10, 887. [Google Scholar] [CrossRef] [Green Version]
- Pimont, F.; Ruffault, J.; Martin-Stpaul, N.K.; Dupuy, J.L. A cautionary note regarding the use of cumulative burnt areas for the determination of fire danger index breakpoints. Int. J. Wildl. Fire 2019. [Google Scholar] [CrossRef]
- Kimmins, J.P. Forest Ecology—A Foundation for Sustainable Forest Management and Environmental Ethics in Forestry; Pearson: London, UK, 2004. [Google Scholar]
- Pausas, J. Fuego y evolución en el Mediterráneo. Investig. Cienc. 2010, 407, 56–63. [Google Scholar]
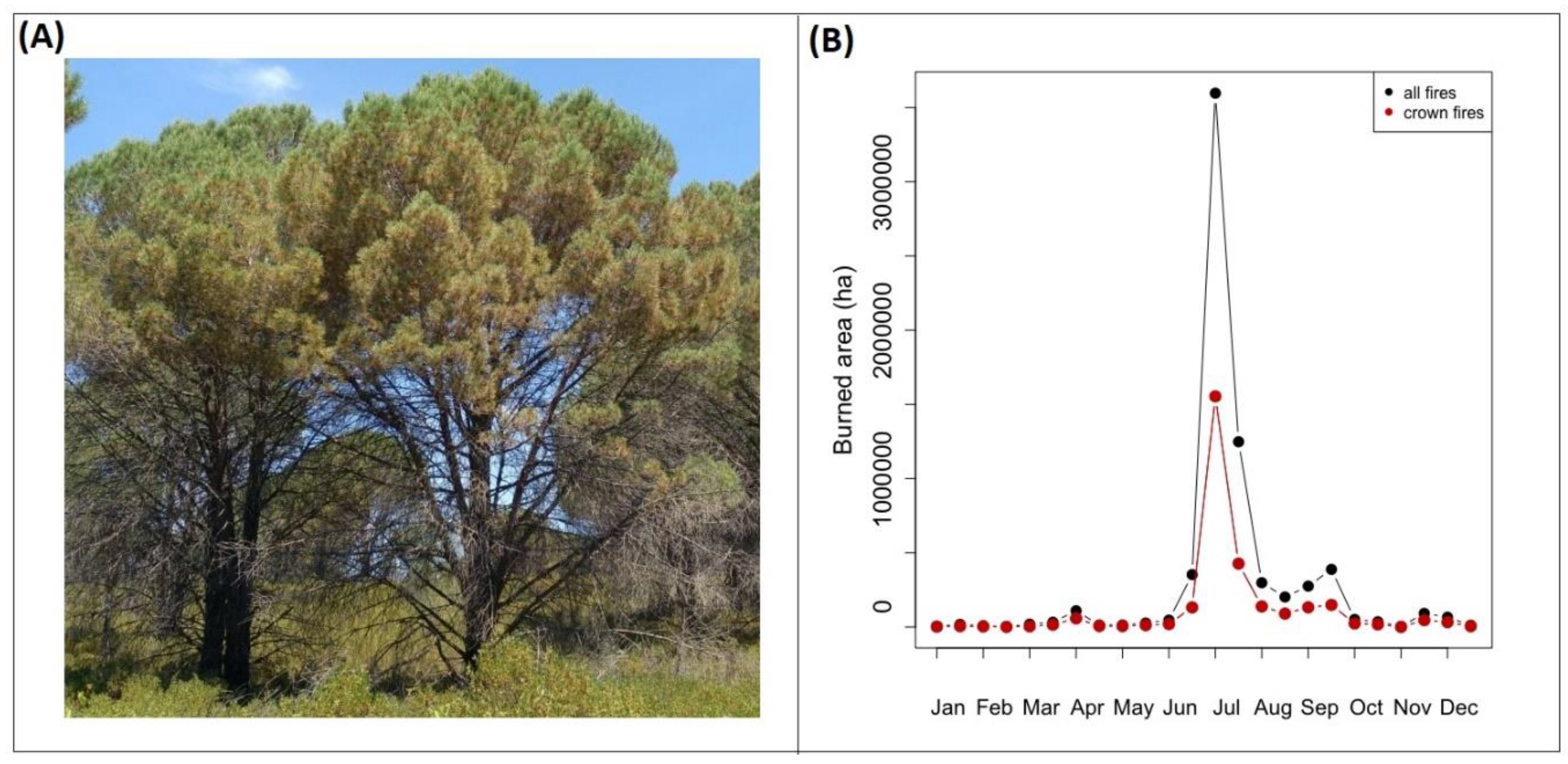




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest (TU-3) | A | B | C | D |
| Canopy Cover (%) | 35 | 35 | 35 | 35 |
| Canopy Height (m) | 8 | 8 | 8 | 8 |
| Canopy Base Height (m) | 1.5 | 1.5 | 1.5 | 1.5 |
| Canopy Bulk Density (kg/m3) | 0.15 | 0.15 | 0.1 | 0.1 |
| Fine Fuel Load (t/ha) | 2.5 | 2.5 | 3 | 2.5 |
| 1-h Dead Surface Fuel Moisture (%) | 6 | 5 | 5 | 9 |
| 10-h Dead Surface Fuel Moisture (%) | 7 | 6 | 6 | 10 |
| 100-h Dead Surface Fuel Moisture (%) | 8 | 7 | 7 | 11 |
| Foliar Moisture Content (%) | 105 | 74 | 100 | 100 |
| Shrub (SH-9) | A | B | C | D |
| Canopy Cover (%) | 100 | 100 | 100 | 100 |
| Canopy Height (m) | 5 | 5 | 5 | 5 |
| Canopy Base Height (m) | 1 | 1 | 1 | 1 |
| Canopy Bulk Density (kg/m3) | 0.22 | 0.22 | 0.15 | 0.15 |
| Fine Fuel Load (t/ha) | 10 | 10 | 10.7 | 10 |
| 1-h Dead Surface Fuel Moisture (%) | 6 | 5 | 5 | 9 |
| 10-h Dead Surface Fuel Moisture (%) | 7 | 6 | 6 | 10 |
| 100-h Dead Surface Fuel Moisture (%) | 8 | 7 | 7 | 11 |
| Foliar Moisture Content (%) | 105 | 74 | 100 | 100 |
| Fuel Parameters | Fuel Model TU-3 | Fuel Model SH-9 |
|---|---|---|
| 1-h Dead Fuel Load | 2.5 t/ha | 10 t/ha |
| 10-h Dead Fuel Load | 0.34 t/ha | 5.5 t/ha |
| 100-h Dead Fuel Load | 0.56 t/ha | 0 t/ha |
| Live Herbaceous Fuel Load | 1.5 t/ha | 3.5 t/ha |
| Live Woody Fuel Load | 2.5 t/ha | 16 t/ha |
| 1-h SAV Ratio | 59.05 cm2/cm3 | 24.60 cm2/cm3 |
| Live Herbaceous SAV Ratio | 52.49 cm2/cm3 | 59.05 cm2/cm3 |
| Live Woody SAV Ratio | 45.93 cm2/cm3 | 49.21 cm2/cm3 |
| Fuel Bed Depth | 40 cm | 134 cm |
| Dead Fuel Moisture of Extinction | 30% | 40% |
| Dead Fuel Heat Content | 18,622.3 kJ/kg | 18,622.3 kJ/kg |
| Live Fuel Heat Content | 18,622.3 kJ/kg | 18,622.3 kJ/kg |
| FOREST (TU-3) | Wind Speed (km/h) | A | B | C | D |
| Rate of Spread (m/min) | 0 | 0.3 | 0.5 | 0.4 | 0.3 |
| 10 | 0.9 | 1.2 | 1.1 | 0.9 | |
| 20 | 1.7 | 5.1 | 2.6 | 1.7 | |
| 30 | 5.8 | 14.6 | 6.9 | 3.9 | |
| Fire Line Intensity (kW/m) | 0 | 48 | 74 | 69 | 45 |
| 10 | 130 | 200 | 183 | 121 | |
| 20 | 259 | 1384 | 462 | 240 | |
| 30 | 1393 | 5924 | 1585 | 653 | |
| Flame Length (m) | 0 | 0.5 | 0.6 | 0.5 | 0.4 |
| 10 | 0.7 | 0.9 | 0.9 | 0.7 | |
| 20 | 1 | 3.3 | 1.6 | 1 | |
| 30 | 3.3 | 8.7 | 3.6 | 2 | |
| Crown Fraction Burned | 0 | 0 | 0 | 0 | 0 |
| 10 | 0 | 0 | 0 | 0 | |
| 20 | 0 | 0.35 | 0.06 | 0 | |
| 30 | 0.30 | 0.81 | 0.32 | 0.13 | |
| SHRUB (SH-9) | Wind Speed (km/h) | A | B | C | D |
| Rate of Spread (m/min) | 0 | 0.7 | 1 | 0.8 | 0.7 |
| 10 | 2.1 | 3.1 | 2.2 | 1.8 | |
| 20 | 5.7 | 8.6 | 5.5 | 4.4 | |
| 30 | 12.6 | 17.7 | 11.6 | 9.1 | |
| Fire Line Intensity (kW/m) | 0 | 560 | 765 | 586 | 490 |
| 10 | 1752 | 2615 | 1679 | 1330 | |
| 20 | 5208 | 8228 | 4510 | 3402 | |
| 30 | 12,562 | 17,179 | 10,074 | 7372 | |
| Flame Length (m) | 0 | 1.8 | 2.2 | 1.9 | 1.7 |
| 10 | 3.9 | 5.1 | 3.8 | 3.2 | |
| 20 | 8.0 | 10.9 | 7.3 | 6 | |
| 30 | 14.4 | 17.7 | 12.4 | 10.1 | |
| Crown Fraction Burned | 0 | 0.13 | 0.19 | 0.1 | 0.08 |
| 10 | 0.34 | 0.44 | 0.27 | 0.23 | |
| 20 | 0.63 | 0.79 | 0.49 | 0.43 | |
| 30 | 0.95 | 1 | 0.75 | 0.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balaguer-Romano, R.; Díaz-Sierra, R.; Madrigal, J.; Voltas, J.; Resco de Dios, V. Needle Senescence Affects Fire Behavior in Aleppo Pine (Pinus halepensis Mill.) Stands: A Simulation Study. Forests 2020, 11, 1054. https://doi.org/10.3390/f11101054
Balaguer-Romano R, Díaz-Sierra R, Madrigal J, Voltas J, Resco de Dios V. Needle Senescence Affects Fire Behavior in Aleppo Pine (Pinus halepensis Mill.) Stands: A Simulation Study. Forests. 2020; 11(10):1054. https://doi.org/10.3390/f11101054
Chicago/Turabian StyleBalaguer-Romano, Rodrigo, Rubén Díaz-Sierra, Javier Madrigal, Jordi Voltas, and Víctor Resco de Dios. 2020. "Needle Senescence Affects Fire Behavior in Aleppo Pine (Pinus halepensis Mill.) Stands: A Simulation Study" Forests 11, no. 10: 1054. https://doi.org/10.3390/f11101054
APA StyleBalaguer-Romano, R., Díaz-Sierra, R., Madrigal, J., Voltas, J., & Resco de Dios, V. (2020). Needle Senescence Affects Fire Behavior in Aleppo Pine (Pinus halepensis Mill.) Stands: A Simulation Study. Forests, 11(10), 1054. https://doi.org/10.3390/f11101054