Regional Climate, Edaphic Conditions and Establishment Substrates Interact to Influence Initial Growth of Black Spruce and Jack Pine Planted in the Boreal Forest




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Area and Data Collection
2.2. Statistical Analyses
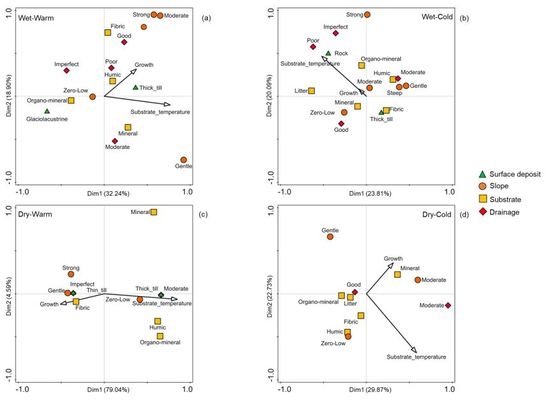
3. Results
3.1. Black Spruce
3.2. Jack Pine
4. Discussion
4.1. Black Spruce
4.2. Jack Pine
5. Conclusions and Implications for Forest Management
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Grandpré, L.; Bergeron, Y.; Nguyen-Xuan, T.-V.; Boudreault, C.; Grondin, P. Composition and dynamics of the understory vegetation in the boreal forests of Quebec. In The Herbaceous Layer of Forests of Eastern North America; Gilliam, F.S., Roberts, M.R., Eds.; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Zhang, S.Y.; Koubaa, A.; FPInnovations (Institut). Les résineux de l’Est du Canada: Écologie Forestière, Caractéristiques, Transformation et Usages; FPInnovations: Pointe-Claire, QC, Canada, 2009; ISBN 978-0-86488-538-8. [Google Scholar]
- Natural Resources Canada. The State of Canada’s Forests. Available online: https://www.nrcan.gc.ca/our-natural-resources/forests-forestry/state-canadas-forests-report/16496 (accessed on 4 September 2019).
- National Forestry Database Area Planted and Seeded, and Number of Seedlings Planted. Terms and Conditions Apply. Available online: https://public.tableau.com/views/6_RegenLandingPage/6_EN?:embed=y&:showVizHome=no&:host_url=https%3A%2F%2Fpublic.tableau.com%2F&:embed_code_version=3&:tabs=no&:toolbar=yes&:animate_transition=yes&:display_static_image=no&:display_spinner=no&:display_overlay=yes&:display_count=yes&:loadOrderID=0 (accessed on 25 November 2019).
- Salmon, D. Ressources et Industries Forestières—Portrait Statistique 2017; Ministère des Forêts, de la Faune et des Parcs: Québec, QC, Canada, 2018; p. 133. [Google Scholar]
- Reich, P.B.; Tjoelker, M.G.; Walters, M.B.; Vanderklein, D.W.; Buschena, C. Close association of RGR, leaf and root morphology, seed mass and shade tolerance in seedlings of nine boreal tree species grown in high and low light. Funct. Ecol. 1998, 12, 327–338. [Google Scholar] [CrossRef]
- Prévost, M. Effets du scarifiage sur les propriétés du sol, la croissance des semis et la compétition: Revue des connaissances actuelles et perspectives de recherches au Québec. Ann. Sci. For. 1992, 49, 277–296. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.G.; Macdonald, S.E.; Rothwell, R.L. Impacts of harvesting and mechanical site preparation on soil chemical properties of mixed-wood boreal forest sites in Alberta. Can. J. Soil Sci. 1996, 76, 531–540. [Google Scholar] [CrossRef] [Green Version]
- Prévost, M. Predicting Soil Properties from Organic Matter Content following Mechanical Site Preparation of Forest Soils. Soil Sci. Soc. Am. J. 2004, 68, 943–949. [Google Scholar] [CrossRef]
- Henneb, M.; Valeria, O.; Thiffault, N.; Fenton, N.J.; Bergeron, Y. Effects of Mechanical Site Preparation on Microsite Availability and Growth of Planted Black Spruce in Canadian Paludified Forests. Forests 2019, 10, 670. [Google Scholar] [CrossRef] [Green Version]
- Henneb, M.; Valeria, O.; Thiffault, N.; Fenton, N.J. Black spruce seedling growth response to organic and organic-mineral substrates. Silva Fenn. 2019. [Google Scholar] [CrossRef]
- Margolis, H.A.; Brand, D.G. An ecophysiological basis for understanding plantation establishment. Can. J. For. Res. 1990, 20, 375–390. [Google Scholar] [CrossRef]
- Spittlehouse, D.L.; Stathers, R.J. Seedling Microclimate; Land Management Report—Ministry of Forests, British Columbia; Ministry of Forests: Victoria, BC, Canada, 1990. [Google Scholar]
- Sutherland, B.J.; Foreman, F.F. Guide to the Use of Mechanical Site Preparation Equipment in Northwestern Ontario; Natural Resources Canada, Canadian Forest Service, Great Lakes Forestry Centre, Sault Ste. Marie, Ontario, Ontario Ministry of Natural Resources: Toronto, ON, Canada, 1995. [Google Scholar]
- Barras, N.; Kellman, M. The supply of regeneration micro-sites and segregation of tree species in a hardwood/boreal forest transition zone. J. Biogeogr. 1998, 25, 871–881. [Google Scholar] [CrossRef]
- Bergeron, Y.; Drapeau, P.; Gauthier, S.; Lecomte, N. Using knowledge of natural disturbances to support sustainable forest management in the northern Clay Belt. For. Chron. 2007, 83, 326–337. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, M.; Paré, D.; Bergeron, Y. Relationships between microsite type and the growth and nutrition of young black spruce on post-disturbed lowland black spruce sites in eastern Canada. Can. J. For. Res. 2007, 37, 62–73. [Google Scholar] [CrossRef]
- Lafleur, B.; Paré, D.; Fenton, N.J.; Bergeron, Y. Growth and nutrition of black spruce seedlings in response to disruption of Pleurozium and Sphagnum moss carpets in boreal forested peatlands. Plant Soil 2011, 345, 141–153. [Google Scholar] [CrossRef]
- Simon, A.; Gratzer, G.; Sieghardt, M. The influence of windthrow microsites on tree regeneration and establishment in an old growth mountain forest. For. Ecol. Manag. 2011, 262, 1289–1297. [Google Scholar] [CrossRef]
- Henneb, M.; Valeria, O.; Fenton, N.J.; Thiffault, N.; Bergeron, Y. Mechanical site preparation: Key to microsite creation success on Clay Belt paludified sites. For. Chron. 2015, 91, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Hansen, A.J.; Neilson, R.P.; Dale, V.H.; Flather, C.H.; Iverson, L.R.; Currie, D.J.; Shafer, S.; Cook, R.; Bartlein, P.J. Global Change in Forests: Responses of Species, Communities, and Biomes: Interactions between climate change and land use are projected to cause large shifts in biodiversity. BioScience 2001, 51, 765–779. [Google Scholar] [CrossRef]
- Walker, K.V.; Davis, M.B.; Sugita, S. Climate Change and Shifts in Potential Tree Species Range Limits in the Great Lakes Region. J. Gt. Lakes Res. 2002, 28, 555–567. [Google Scholar] [CrossRef]
- Shafer, S.L.; Bartlein, P.J.; Thompson, R.S. Potential Changes in the Distributions of Western North America Tree and Shrub Taxa under Future Climate Scenarios. Ecosystems 2001, 4, 200–215. [Google Scholar] [CrossRef] [Green Version]
- Goldblum, D.; Rigg, L.S. Tree growth response to climate change at the deciduous–boreal forest ecotone, Ontario, Canada. Can. J. For. Res. 2005, 35, 2709–2718. [Google Scholar] [CrossRef]
- Boucher, D.; Gauthier, S.; Thiffault, N.; Marchand, W.; Girardin, M.; Urli, M. How climate change might affect tree regeneration following fire at northern latitudes: A review. New For. 2019. [Google Scholar] [CrossRef] [Green Version]
- Saucier, J.P.; Robitaille, A.; Grondin, P. Cadre bioclimatique du Québec. In Manuel de Foresterie; Editions Multimondes Inc.: Montreal, QC, Canada, 2009; pp. 186–205. [Google Scholar]
- MFFP (Ministère des Forêts, de la Faune et des Parcs). Qualité des Plantations—Guide de l’évaluateur; Gouvernement du Québec, Direction de l’aménagement et de l’environnement Forestiers. 2016. Available online: https://mffp.gouv.qc.ca/publications/forets/connaissances/guide-evaluateur-qualite-plantations.pdf (accessed on 9 April 2019).
- MFFP (Ministère des Forêts, de la Faune et des Parcs). Inventaire écoforestier du Québec Méridional (IÉQM). Available online: https://mffp.gouv.qc.ca/les-forets/inventaire-ecoforestier/ (accessed on 25 November 2019).
- R Core Team. R: A Language and Environment for Statistical Computing; R Fondation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 24 January 2020).
- Avery, T.E.; Burkhart, H.E. Forest Measurements, 5th ed.; Waveland Press: Long Grove, IL, USA, 2015; ISBN 978-1-4786-2974-0. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Leys, C.; Ley, C.; Klein, O.; Bernard, P.; Licata, L. Detecting outliers: Do not use standard deviation around the mean, use absolute deviation around the median. J. Exp. Soc. Psychol. 2013, 49, 764–766. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-S.; Wessel, P. Directional median filtering for regional-residual separation of bathymetry. Geochem. Geophys. Geosystems 2008, 9. [Google Scholar] [CrossRef] [Green Version]
- Werner, C.; Davis, K.; Bakwin, P.; Yi, C.; Hurst, D.; Lock, L. Regional-scale measurements of CH4 exchange from a tall tower over a mixed temperate/boreal lowland and wetland forest. Glob. Change Biol. 2003, 9, 1251–1261. [Google Scholar] [CrossRef]
- Diaci, J. Regeneration dynamics in a Norway spruce plantation on a silver fir-beech forest site in the Slovenian Alps. For. Ecol. Manag. 2002, 161, 27–38. [Google Scholar] [CrossRef]
- Wickham, J.D.; O’Neill, R.V.; Jones, K.B. Forest fragmentation as an economic indicator. Landsc. Ecol. 2000, 15, 171–179. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Smilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012. [Google Scholar]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014; ISBN 978-1-107-69440-8. [Google Scholar]
- Herr, D.G.; Duchesne, L.C. Jack pine (Pinus banksiana) seedling emergence is affected by organic horizon removal, ashes, soil, water and shade. Water. Air. Soil Pollut. 1995, 82, 147–154. [Google Scholar] [CrossRef]
- Hamilton, W.N.; Krause, H.H. Relationship between jack pine growth and site variables in New Brunswick plantations. Can. J. For. Res. 1985, 15, 922–926. [Google Scholar] [CrossRef]
- Walker, X.; Johnstone, J.F. Widespread negative correlations between black spruce growth and temperature across topographic moisture gradients in the boreal forest. Environ. Res. Lett. 2014, 9, 064016. [Google Scholar] [CrossRef]
- Hamel, B.; Bélanger, N.; Paré, D. Productivity of black spruce and Jack pine stands in Quebec as related to climate, site biological features and soil properties. For. Ecol. Manag. 2004, 191, 239–251. [Google Scholar] [CrossRef]
- Strong, W.L.; Roi, G.H.L. Rooting depths and successional development of selected boreal forest communities. Can. J. For. Res. 1983, 13, 577–588. [Google Scholar] [CrossRef]
- Steele, S.J.; Gower, S.T.; Vogel, J.G.; Norman, J.M. Root mass, net primary production and turnover in aspen, jack pine and black spruce forests in Saskatchewan and Manitoba, Canada. Tree Physiol. 1997, 17, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Barber, V.A.; Juday, G.P.; Finney, B.P. Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature 2000, 405, 668–673. [Google Scholar] [CrossRef] [PubMed]
- D’Arrigo, R.; Wilson, R.; Liepert, B.; Cherubini, P. On the ‘Divergence Problem’ in Northern Forests: A review of the tree-ring evidence and possible causes. Glob. Planet Chang. 2008, 60, 289–305. [Google Scholar] [CrossRef]
- Gower, S.T.; McMurtrie, R.E.; Murty, D. Aboveground net primary production decline with stand age: Potential causes. Trends Ecol. Evol. 1996, 11, 378–382. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Ecophysiology of Northern Spruce Species; NRC Research Press: Ottawa, ON, Canada, 2000; ISBN 978-0-660-17959-9. [Google Scholar]
- Prescott, C.E.; Maynard, D.G.; Laiho, R. Humus in northern forests: Friend or foe? For. Ecol. Manag. 2000, 133, 23–36. [Google Scholar] [CrossRef]
- Viereck, L.A.; Dyrness, C.T.; Cleve, K.V.; Foote, M.J. Vegetation, soils, and forest productivity in selected forest types in interior Alaska. Can. J. For. Res. 1983, 13, 703–720. [Google Scholar] [CrossRef]
- Thiffault, N.; Jobidon, R.; Munson, A. Performance and physiology of large containerized and bare-root spruce seedlings in relation to scarification and competition in Québec (Canada). Ann. For. Sci. 2003, 60, 645–655. [Google Scholar] [CrossRef] [Green Version]
- Bruand, A.; Tessier, D. Water retention properties of the clay in soils developed on clayey sediments: Significance of parent material and soil history. Eur. J. Soil Sci. 2000, 51, 679–688. [Google Scholar] [CrossRef]
- Boivin, P.; Garnier, P.; Tessier, D. Relationship between Clay Content, Clay Type, and Shrinkage Properties of Soil Samples. Soil Sci. Soc. Am. J. 2004, 68, 1145–1153. [Google Scholar] [CrossRef] [Green Version]
- Körner, C. Carbon limitation in trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Deslauriers, A.; Griçar, J.; Seo, J.-W.; Rathgeber, C.B.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; King, J.S.; Burton, A.J.; Brown, S.E. Responses of tree fine roots to temperature. New Phytol. 2000, 147, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Londo, A.J.; Mroz, G.D. Bucket Mounding as a Mechanical Site Preparation Technique in Wetlands. North. J. Appl. For. 2001, 18, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Kabrick, J.M.; Dey, D.C.; Sambeek, J.W.V.; Wallendorf, M.; Gold, M.A. Soil properties and growth of swamp white oak and pin oak on bedded soils in the lower Missouri River floodplain. For. Ecol. Manag. 2005, 204, 315–327. [Google Scholar] [CrossRef]
- Löf, M.; Birkedal, M. Direct seeding of Quercus robur L. for reforestation: The influence of mechanical site preparation and sowing date on early growth of seedlings. For. Ecol. Manag. 2009, 258, 704–711. [Google Scholar] [CrossRef]
- Dang, Q.-L.; Cheng, S. Effects of soil temperature on ecophysiological traits in seedlings of four boreal tree species. For. Ecol. Manag. 2004, 194, 379–387. [Google Scholar] [CrossRef]
- Way, D.A.; Sage, R.F. Elevated growth temperatures reduce the carbon gain of black spruce [Picea mariana (Mill.) B.S.P.]. Glob. Change Biol. 2008, 14, 624–636. [Google Scholar] [CrossRef]
- Sutherland, B.; Foreman, F.F. Black spruce and vegetation response to chemical and mechanical site preparation on a boreal mixedwood site. Can. J. For. Res. 2000, 30, 1561–1570. [Google Scholar] [CrossRef]
- Prévost, M.; Dumais, D. Croissance et statut nutritif de marcottes, de semis naturels et de plants d’épinette noire à la suite du scarifiage: Résultats de 10 ans. Can. J. For. Res. 2003, 33, 2097–2107. [Google Scholar] [CrossRef]
- Satoh, T. Organo-mineral complex status in soils. I. thermal analytical characteristics of humus in the soils. Soil Sci. Plant Nutr. 1984, 30, 1–12. [Google Scholar] [CrossRef]
- Moskal, T.D.; Leskiw, L.; Naeth, M.A.; Chanasyk, D.S. Effect of organic carbon (peat) on moisture retention of peat:mineral mixes. Can. J. Soil Sci. 2001, 81, 205–211. [Google Scholar] [CrossRef]
- Anderson, D.W.; Paul, E.A. Organo-Mineral Complexes and Their Study by Radiocarbon Dating 1. Soil Sci. Soc. Am. J. 1984, 48, 298–301. [Google Scholar] [CrossRef] [Green Version]
- DesRochers, A.; Gagnon, R. Erratum: Is ring count at ground level a good estimation of black spruce age? Can. J. For. Res. 1997, 27, 1703. [Google Scholar] [CrossRef]
- Krause, C.; Morin, H. Adventive-root development in mature black spruce and balsam fir in the boreal forests of Quebec, Canada. Can. J. For. Res. 2005, 35, 2642–2654. [Google Scholar] [CrossRef]
- Bergsten, U.; Goulet, F.; Lundmark, T.; Löfvenius, M.O. Frost heaving in a boreal soil in relation to soil scarification and snow cover. Can. J. For. Res. 2001, 31, 1084–1092. [Google Scholar] [CrossRef]
- De Chantal, M.; Leinonen, K.; Ilvesniemi, H.; Westman, C.J. Combined effects of site preparation, soil properties, and sowing date on the establishment of Pinus sylvestris and Picea abies from seeds. Can. J. For. Res. 2003, 33, 931–945. [Google Scholar] [CrossRef]
- Lavoie, M.; Paré, D.; Fenton, N.; Groot, A.; Taylor, K. Paludification and management of forested peatlands in Canada: A literature review. Environ. Rev. 2005, 13, 21–50. [Google Scholar] [CrossRef]
- Van Cleve, K.; Oliver, L.; Schlentner, R.; Viereck, L.A.; Dyrness, C.T. Productivity and nutrient cycling in taiga forest ecosystems. Can. J. For. Res. 1983, 13, 747–766. [Google Scholar] [CrossRef]
- Li, J.; Ziegler, S.; Lane, C.S.; Billings, S.A. Warming-enhanced preferential microbial mineralization of humified boreal forest soil organic matter: Interpretation of soil profiles along a climate transect using laboratory incubations. J. Geophys. Res. Biogeosci. 2012, 117. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.Y.; Dang, Q.-L. Effects of soil temperature on biomass production and allocation in seedlings of four boreal tree species. For. Ecol. Manag. 2003, 180, 1–9. [Google Scholar] [CrossRef]
- Zhang, S.; Dang, Q.-L. Interactive Effects of Soil Temperature and [CO2] on Morphological and Biomass Traits in Seedlings of Four Boreal Tree Species. For. Sci. 2007, 53, 453–460. [Google Scholar]
- Reich, P.B.; Sendall, K.M.; Rice, K.; Rich, R.L.; Stefanski, A.; Hobbie, S.E.; Montgomery, R.A. Geographic range predicts photosynthetic and growth response to warming in co-occurring tree species. Nat. Clim. Chang. 2015, 5, 148–152. [Google Scholar] [CrossRef]
- Bell, F.W.; Ter-Mikaelian, M.T.; Wagner, R.G. Relative competitiveness of nine early-successional boreal forest species associated with planted jack pine and black spruce seedlings. Can. J. For. Res. 2000, 30, 790–800. [Google Scholar] [CrossRef]
- Day, M.E.; Schedlbauer, J.L.; Livingston, W.H.; Greenwood, M.S.; White, A.S.; Brissette, J.C. Influence of seedbed, light environment, and elevated night temperature on growth and carbon allocation in pitch pine (Pinus rigida) and jack pine (Pinus banksiana) seedlings. For. Ecol. Manag. 2005, 205, 59–71. [Google Scholar] [CrossRef]
- Strimbu, V.C.; Bokalo, M.; Comeau, P.G. Deterministic Models of Growth and Mortality for Jack Pine in Boreal Forests of Western Canada. Forests 2017, 8, 410. [Google Scholar] [CrossRef] [Green Version]
- Sirois, L. Impact of fire on Picea mariana and Pinus banksiana seedlings in subarctic lichen woodlands. J. Veg. Sci. 1993, 4, 795–802. [Google Scholar] [CrossRef]
- Blake, T.J.; Li, J. Hydraulic adjustment in jack pine and black spruce seedlings under controlled cycles of dehydration and rehydration. Physiol. Plant. 2003, 117, 532–539. [Google Scholar] [CrossRef]
- Hébert, F.; Boucher, J.-F.; Bernier, P.Y.; Lord, D. Growth response and water relations of 3-year-old planted black spruce and jack pine seedlings in site prepared lichen woodlands. For. Ecol. Manag. 2006, 223, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Chesick, E.E.; Bergmann, B.A. Jack Pine (Pinus banksiana Lamb.). In Trees III.; Bajaj, Y.P.S., Ed.; Biotechnology in Agriculture and Forestry; Springer: Berlin/Heidelberg, Germany, 1991; pp. 241–253. ISBN 978-3-662-13231-9. [Google Scholar]
- Little, S.; Garrett, P.W. Pitch pine. In Silvics of North America; Burns, R.M., Honkala, B.H., Eds.; US Government Printing Office: Washington, DC, USA, 1990; pp. 456–462. [Google Scholar]
- Rudolph, T.D.; Laidly, P.R. Jack Pine (Pinus banksiana Lamb.). In Silvics of North America: 1. Conifers; Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; US Government Printing Office: Washington, DC, USA, 1990; p. 877. [Google Scholar]
- Béland, M.; Bergeron, Y. Height growth of jack pine (Pinus banksiana) in relation to site types in boreal forests of Abitibi, Quebec. Can. J. For. Res. 1996, 26, 2170–2179. [Google Scholar] [CrossRef]
- Béland, M.; Bergeron, Y.; Zarnovican, R. Natural regeneration of jack pine following harvesting and site preparation in the Clay Belt of northwestern Quebec. For. Chron. 1999, 75, 821–831. [Google Scholar] [CrossRef] [Green Version]
- Chrosciewicz, Z. Jack Pine Regeneration Following Postcut Burning under Seed Trees in Central Saskatchewan. For. Chron. 1988, 64, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Chrosciewicz, Z. Site Conditions for Jack Pine Seeding. For. Chron. 1990, 66, 579–584. [Google Scholar] [CrossRef] [Green Version]
- Boiffin, J.; Munson, A.D. Three large fire years threaten resilience of closed crown black spruce forests in eastern Canada. Ecosphere 2013, 4, art56. [Google Scholar] [CrossRef]
- Kenkel, N.C. Structure and dynamics of jack pine stands near Elk Lake, Ontario: A multivariate approach. Can. J. Bot. 1986, 64, 486–497. [Google Scholar] [CrossRef]
- Amiro, B.D.; Stocks, B.J.; Alexander, M.E.; Flannigan, M.D.; Wotton, B.M. Fire, climate change, carbon and fuel management in the Canadian boreal forest. Int. J. Wildland Fire 2001, 10, 405–413. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Region | Number of Seedlings | Ground Collar Diameter (mm) | Height (cm) |
|---|---|---|---|---|
| Black spruce | Wet–Warm | 1120 | 18.0 (±6.3) | 89.7 (±26.8) |
| Wet–Cold | 1289 | 12.9 (±5.2) | 61.8 (±27.0) | |
| Dry–Warm | 239 | 9.6 (±4.6) | 51.4 (±18.2) | |
| Dry–Cold | 348 | 9.6 (±6.6) | 47.1 (±21.9) | |
| Jack pine | Dry–Warm | 1034 | 15.2 (±6.7) | 69.3 (±25.1) |
| Dry–Cold | 462 | 10.2 (±2.5) | 44.1 (±9.6) |
| Explanatory Variables | Black Spruce (Model R2 = 0.61) | Jack Pine (Model R2 = 0.75) | |||
|---|---|---|---|---|---|
| F-Value | p-Value * | F-Value | p-Value * | ||
| Microsite scale | Humus thickness | 0.944 | 0.331 | 0.790 | 0.374 |
| Planting position | 1.055 | 0.384 | 0.357 | 0.840 | |
| Substrate temperature (°C) | 0.649 | 0.420 | 2.775 | 0.096 | |
| Substrate type | 1.429 | 0.199 | 1.661 | 0.127 | |
| Stand scale | Surface deposit | 0.598 | 0.621 | 5.627 | 0.001 |
| Drainage class | 1.608 | 0.206 | 0.291 | 0.884 | |
| Slope class | 3.928 | 0.007 | 2.773 | 0.040 | |
| Regional scale | Precipitation (mm) | 0.103 | 0.748 | 1.953 | 0.162 |
| Temperature (°C) | 8.067 | 0.009 | 1.425 | 0.233 | |
| Relative humidity (%) | 0.067 | 0.796 | 1.030 | 0.310 | |
| Interactions | Substrate type × Substrate temperature | 1.399 | 0.211 | 2.713 | 0.008 |
| Substrate type × Precipitation | 3.742 | 0.001 | 4.385 | 0.002 | |
| Substrate type × Relative humidity | 1.604 | 0.142 | 1.954 | 0.070 | |
| Substrate type × Planting position | 0.889 | 0.633 | 1.544 | 0.055 | |
| Substrate type × Surface deposit | 2.031 | 0.018 | 3.205 | 0.001 | |
| Substrate type × Drainage class | 1.050 | 0.399 | 2.069 | 0.006 | |
| Humus thickness × Surface deposit | 1.170 | 0.320 | 0.414 | 0.661 | |
| Planting position × Substrate temperature | 1.307 | 0.258 | 0.968 | 0.436 | |
| Planting position × Surface deposit | 0.859 | 0.603 | 1.309 | 0.219 | |
| Planting position × Drainage class | 1.443 | 0.139 | 0.807 | 0.671 | |
| Surface deposit × Substrate temperature | 2.781 | 0.043 | 0.300 | 0.825 | |
| Surface deposit × Drainage class | 2.329 | 0.034 | 1.039 | 0.385 | |
| Surface deposit × Slope class | 0.580 | 0.679 | 1.355 | 0.255 | |
| Drainage class × Slope class | 1.977 | 0.101 | 2.334 | 0.072 | |
| Substrate type × Substrate temperature × Surface deposit | 0.750 | 0.690 | 2.222 | 0.030 | |
| Substrate type × Planting position × Drainage class | 1.169 | 0.241 | 1.241 | 0.206 | |
| Substrate type × Surface deposit × Drainage class | 2.134 | 0.008 | 0.934 | 0.505 | |
| Substrate type × Drainage class × Slope class | 2.522 | 0.002 | 0.827 | 0.578 | |
| Planting position × Substrate type × Substrate temperature | 1.402 | 0.096 | 0.810 | 0.676 | |
| Planting position × Drainage class × Surface deposit | 0.920 | 0.565 | 0.926 | 0.508 | |
| Surface deposit × Drainage class × Slope class | 0.563 | 0.573 | 0.042 | 0.839 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henneb, M.; Thiffault, N.; Valeria, O. Regional Climate, Edaphic Conditions and Establishment Substrates Interact to Influence Initial Growth of Black Spruce and Jack Pine Planted in the Boreal Forest. Forests 2020, 11, 139. https://doi.org/10.3390/f11020139
Henneb M, Thiffault N, Valeria O. Regional Climate, Edaphic Conditions and Establishment Substrates Interact to Influence Initial Growth of Black Spruce and Jack Pine Planted in the Boreal Forest. Forests. 2020; 11(2):139. https://doi.org/10.3390/f11020139
Chicago/Turabian StyleHenneb, Mohammed, Nelson Thiffault, and Osvaldo Valeria. 2020. "Regional Climate, Edaphic Conditions and Establishment Substrates Interact to Influence Initial Growth of Black Spruce and Jack Pine Planted in the Boreal Forest" Forests 11, no. 2: 139. https://doi.org/10.3390/f11020139