Assessing Differences in Competitive Effects Among Tree Species in Central British Columbia, Canada


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Data
2.2. Choice of a Competition Index
2.3. Basal Area Growth Models
2.4. Analyses
3. Results
3.1. Change in Stand Characteristics
3.2. Models for Predicting PABAI
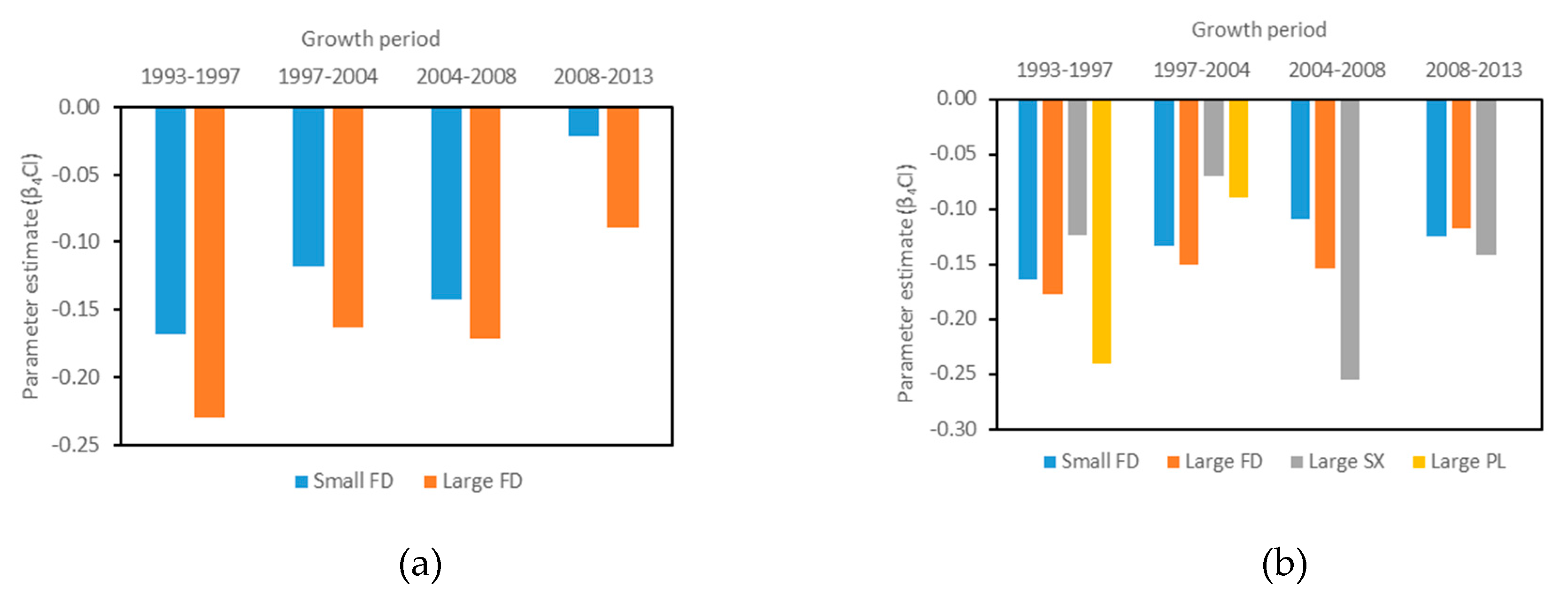
3.3. Effect of Competition on the Growth of the Subject Trees
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A




References
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed]
- Forrester, D.I.; Bauhus, J. A review of processes behind diversity—Productivity relationships in forests. Curr. For. Reports 2016, 2, 45–61. [Google Scholar] [CrossRef] [Green Version]
- Heym, M.; Bielak, K.; Wellhausen, K.; Uhl, E.; Biber, P.; Perkins, D.; Steckel, M.; Thurm, E.A.; Rais, A.; Pretzsch, H. A new method to reconstruct recent tree and stand attributes of temporary research plots: New opportunity to analyse mixed forest stands. In Conifers; IntechOpen: London, UK, 2018; p. 44. [Google Scholar] [CrossRef] [Green Version]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics: Update Edition; FES Other Publications; Yale University: New Haven, CT, USA, 1996; pp. 9–39. [Google Scholar]
- Mina, M.; del Río, M.; Huber, M.O.; Thürig, E.; Rohner, B. The symmetry of competitive interactions in mixed Norway spruce, silver fir and European beech forests. J. Veg. Sci. 2018, 29, 775–787. [Google Scholar] [CrossRef]
- Callaway, R.M. Positive Interactions and Interdependence in Plant Communities; Springer: Dordrecht, The Netherlands, 2007; pp. 179–254. [Google Scholar]
- Bertness, M.D.; Callaway, R.M. Positive interactions in communities. Trends. Ecol. Evol. 1994, 9, 191–193. [Google Scholar] [CrossRef]
- Zhao, D.; Borders, B.; Wilson, M.; Rathbun, S.L. Modeling neighborhood effects on the growth and survival of individual trees in a natural temperate species-rich forest. Ecol. Model. 2006, 196, 90–102. [Google Scholar] [CrossRef]
- Hui, G.; Wang, Y.; Zhang, G.; Zhao, Z.; Bai, C.; Liu, W. A novel approach for assessing the neighborhood competition in two different aged forests. For. Ecol. Manag. 2018, 422, 49–58. [Google Scholar] [CrossRef]
- Burton, P.J. Some limitations inherent to static indices of plant competition. Can. J. For. Res. 1993, 23, 2141–2152. [Google Scholar] [CrossRef]
- Li, Q.; Liang, Y.; Tong, B.; Du, X.; Ma, K. Compensatory effects between Pinus massoniana and broadleaved tree species. J. Plant Ecol. 2010, 3, 183–189. [Google Scholar] [CrossRef]
- Hubbell, S.P.; Ahumada, J.A.; Condit, R.; Foster, R.B. Local neighborhood effects on long-term survival of individual trees in a neotropical forest. Ecol. Res. 2001, 16, 859–875. [Google Scholar] [CrossRef]
- von Oheimb, G.; Lang, A.C.; Bruelheide, H.; Forrester, D.I.; Wäsche, I.; Yu, M.; Härdtle, W. Individual-tree radial growth in a subtropical broad-leaved forest: The role of local neighbourhood competition. For. Ecol. Manag. 2011, 261, 499–507. [Google Scholar] [CrossRef]
- Stoll, P.; Newbery, D.M. Evidence of species-specific neighborhood effects in the Dipterocarpaceae of a Bornean rain forest. Ecology 2005, 86, 3048–3062. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; Schütze, G. Transgressive overyielding in mixed compared with pure stands of Norway spruce and European beech in Central Europe: Evidence on stand level and explanation on individual tree level. Eur. J. For. Res. 2009, 128, 183–204. [Google Scholar] [CrossRef]
- Schwinning, S.; Weiner, J. Mechanisms determining the degree of size asymmetry in competition among plants. Oecologia 1998, 113, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Binkley, D. A hypothesis about the interaction of tree dominance and stand production through stand development. For. Ecol. Manag. 2004, 190, 265–271. [Google Scholar] [CrossRef]
- Weiner, J.; Thomas, S.C. Size variability and competition in plant monocultures. Oikos 1986, 47, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Dale, V.H.; Doyle, T.W.; Shugart, H.H. A comparison of tree growth models. Ecol. Model. 1985, 29, 145–169. [Google Scholar] [CrossRef]
- Uriarte, M.; Condit, R.; Canham, C.D.; Hubbell, S.P. A spatially explicit model of sapling growth in a tropical forest: Does the identity of neighbours matter? J. Ecol. 2004, 92, 348–360. [Google Scholar] [CrossRef]
- Marshall, P.L. Response of uneven-aged Douglas-fir to alternative spacing regimes: Description of the project and analysis of the initial impact of the spacing regimes. Canada-British Columbia Economic and Regional Development Agreement. FRDA Report 242. 1996. Available online: http://203.187.160.132:9011/www.llbc.leg.bc.ca/c3pr90ntc0td/public/pubdocs/bcdocs2012_2/243652/frr242.pdf (accessed on 1 February 2020).
- Vyse, A.; Hollstedt, C.; Huggard, D.J. Managing the Dry Douglas-Fir Forests of the Southern Interior: Workshop Proceedings: April 29–30, 1997, Kamloops, British Columbia, Canada; Vyse, A., Hollstedt, C., Huggard, D.J., Eds.; Citeseer: Kamloops, BC, Canada, 1998. [Google Scholar]
- Hope, G.D.; Mitchell, W.R.; Lloyd, D.A.; Erickson, W.R.; Harper, W.L.; Wilkeem, B.M. Interior Douglas-Fir Zone. Ecosystems of British Columbia; Meidinger, D., Pojar, J., Eds.; Research Branch: Victoria, BC, USA, 1991; pp. 153–166. [Google Scholar]
- Day, K. Interior Douglas-fir and selection management at the UBC/Alex Fraser Research Forest. In Proceedings of the IUFRO Uneven-Aged Silviculture Symposium, Corvallis, OR, USA, 15–19 September 1997; pp. 542–552. [Google Scholar]
- Day, K. UBC Alex Fraser Research Forest Management and Working Plan# 3; UBC Alex Fraser Research Forest: Williams Lake, BC, Canada, 2007; 156p, Available online: https://afrf-forestry.sites.olt.ubc.ca/files/2012/03/MWP3WebQuality.pdf (accessed on 1 February 2020).
- Marshall, P.L.; Wang, Y. Growth of Uneven-Aged Interior Douglas-Fir stands as Influenced by Different Stand Structures; Canada-British Columbia Economic and Regional Development Agreement. FRDA Report 267; Canadian Forest Service; B.C. Ministry of Forests: Victoria, BC, Canada, 1996; 20p. Available online: https://www.for.gov.bc.ca/hfd/pubs/Docs/Frr/FRR267.pdf (accessed on 1 February 2020).
- Rivas, J.J.C.; González, J.G.Á.; Aguirre, O.; Hernández, F.J. The effect of competition on individual tree basal area growth in mature stands of Pinus cooperi Blanco in Durango (Mexico). Eur. J. For. Res. 2005, 124, 133–142. [Google Scholar] [CrossRef]
- Kahriman, A.; Şahin, A.; Sönmez, T.; Yavuz, M. A novel approach to selecting a competition index: The effect of competition on individual-tree diameter growth of Calabrian pine. Can. J. For. Res. 2018, 48, 1217–1226. [Google Scholar] [CrossRef]
- Contreras, M.A.M.A.; Affleck, D.; Chung, W. Evaluating tree competition indices as predictors of basal area increment in western Montana forests. For. Ecol. Manag. 2011, 262, 1939–1949. [Google Scholar] [CrossRef]
- Martin, G.L.; Ek, A.R. A comparison of competition measures and growth models for predicting plantation red pine diameter and height growth. For. Sci. 1984, 30, 731–743. [Google Scholar] [CrossRef]
- Steneker, G.A.; Jarvis, J.M. A preliminary study to assess competition in a white spruce-trembling aspen stand. For. Chron. 1963, 39, 334–336. [Google Scholar] [CrossRef] [Green Version]
- Lorimer, C.G. Tests of age-independent competition indices for individual trees in natural hardwood stands. For. Ecol. Manag. 1983, 6, 343–360. [Google Scholar] [CrossRef]
- Glover, G.R.; Hool, J.N. A basal area ratio predictor of loblolly pine plantation mortality. For. Sci. 1979, 25, 275–282. [Google Scholar] [CrossRef]
- Krajicek, J.E.; Brinkman, K.A.; Gingrich, S.F. Crown competition—A measure of density. For. Sci. 1961, 7, 35–42. [Google Scholar]
- Wykoff, W.R.; Crookston, N.L.; Stage, A.R. User’s guide to the stand prognosis model; USDA Forest Service General Technical Report INT-133; United States Department of Agriculture Forest Service: Ogden, UT, USA, 1982; 112p. Available online: https://www.fs.fed.us/rm/pubs_int/int_gtr133.pdf (accessed on 1 February 2020).
- Hegyi, F. A simulation model for managing jack-pine stands. In Proceedings of the Growth Models for Tree and Stand Simulation IUFRO S4.01–4; Fries, J., Ed.; Department of Forest Yield, Royal College of Forestry: Stockholm, Sweden, 1974; pp. 74–90. [Google Scholar]
- Rouvinen, S.; Kuuluvainen, T. Structure and asymmetry of tree crowns in relation to local competition in a natural mature Scots pine forest. Can. J. For. Res. 1997, 27, 890–902. [Google Scholar] [CrossRef]
- Braathe, P. Height increment of young single trees in relation to height and distance of neighbouring trees. Mitteilungen der Forstl. Bundesversuchsanstalt 1980, 130, 43–48. [Google Scholar]
- Opie, J. Predictability of individual tree growth using various definitions of competing basal area. For. Sci. 1968, 14, 314–323. [Google Scholar] [CrossRef]
- Bella, I. A new competition model for individual trees. For. Sci. 1971, 17, 364–372. [Google Scholar] [CrossRef]
- Monserud, R.A.; Ek, A.R. Prediction of understory tree height growth in northern hardwood stands. For. Sci. 1977, 23, 391–400. [Google Scholar] [CrossRef]
- Pretzsch, H. Forest Dynamics, Growth and Yield. From Measurement to Model; Springer: Berlin/Heidelberg, Germany, 2009; p. 300. ISBN 978-3-540-88306-7. [Google Scholar]
- Schröder, J.; Soalleiro, R.R.; Alonso, G.V. An age-independent basal area increment model for maritime pine trees in northwestern Spain. For. Ecol. Manag. 2002, 157, 55–64. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, S.; Meng, S.X.; Trincado, G.; Vanderschaaf, C.L. A multilevel individual tree basal area increment model for aspen in boreal mixedwood stands. Can. J. For. Res. 2009, 39, 2203–2214. [Google Scholar] [CrossRef]
- Del Río, M.; Condés, S.; Pretzsch, H. Analyzing size-symmetric vs. size-asymmetric and intra- vs. inter-specific competition in beech (Fagus sylvatica L.) mixed stands. For. Ecol. Manag. 2014, 325, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Chambers, J., Eddy, W., Sheather, S., Tierney, L., Eds.; Springer-Verlag: New York, NY, USA, 2000; pp. 94, 306–408. ISBN 978-1-4757-8144-1. [Google Scholar]
- R Core Team. R: A Language And Environment For Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://CRAN.R-project.org/package=nlme (accessed on 1 February 2020).
- Vonesh, E.; Chinchilli, V.M. Linear and Nonlinear Models for the Analysis of Repeated Measurements; CRC Press; Marcel Dekker: New York, NY, USA, 1997; p. 546. ISBN 1482293277. [Google Scholar]
- Wang, Y.; LeMay, V.M.; Baker, T.G. Modelling and prediction of dominant height and site index of Eucalyptus globulus plantations using a nonlinear mixed-effects model approach. Can. J. For. Res. 2007, 37, 1390–1403. [Google Scholar] [CrossRef]
- Weiner, J. Asymmetric competition in plant populations. Trends Ecol. Evol. 1990, 5, 360–364. [Google Scholar] [CrossRef]
- Chen, H.Y.H.; Klinka, K.; Kayahara, G.J. Effects of light on growth, crown architecture, and specific leaf area for naturally established Pinus contorta var. latifolia and Pseudotsuga menziesii var. glauca saplings. Can. J. For. Res. 1996, 26, 1149–1157. [Google Scholar] [CrossRef]
- Kayahara, G.J.; Chen, H.Y.H.; Klinka, K.; Coates, K.D. Relations of Terminal Growth and Specific Leaf Area to Available Light in Naturally Regenerated Seedlings of Lodgepole Pine and Interior Spruce in Central British Columbia; British Columbia Minstry of Forests Research Report 09; Ministry of Forests Research Program: Victoria, BC, Canada, 1996; 32p. Available online: https://pdfs.semanticscholar.org/6881/8c2a91e483a53a3252fe01a71a89a10cf5b3.pdf (accessed on 1 February 2020).
- Riofrío, J.; Del Río, M.; Bravo, F. Mixing effects on growth efficiency in mixed pine forests. Forestry 2017, 90, 381–392. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Mohren, G.M.J.; del Río, M.; Schelhaas, M.J.; Bouwman, M.; Sterck, F.J. Species mixing effects on forest productivity: A case study at stand-, species- and tree-level in the Netherlands. Forests 2018, 9, 713. [Google Scholar] [CrossRef] [Green Version]
- Fichtner, A.; Härdtle, W.; Bruelheide, H.; Kunz, M.; Li, Y.; von Oheimb, G. Neighbourhood interactions drive overyielding in mixed-species tree communities. Nat. Commun. 2018, 9, 1144. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, H.C.; Astrup, R.; Trowbridge, A.; Coates, K.D. Competition and tree crowns: A neighborhood analysis of three boreal tree species. For. Ecol. Manag. 2010, 259, 1586–1596. [Google Scholar] [CrossRef]
- Canham, C.D.; Papaik, M.J.; Uriarte, M.; McWilliams, W.H.; Jenkins, J.C.; Twery, M.J. Neighborhood analysis of canopy tree competition along environmental gradients in New England forests. Ecol. Appl. 2006, 16, 540–554. [Google Scholar] [CrossRef] [Green Version]
- Coates, K.D.; Canham, C.D.; LePage, P.T. Above- versus below-ground competitive effects and responses of a guild of temperate tree species. J. Ecol. 2009, 97, 118–130. [Google Scholar] [CrossRef]
- LeMay, V.M.; Pommerening, A.; Marshall, P. Spatio-temporal structure of multi-storied, multi-aged interior Douglas fir (Pseudotsuga menziesii var. glauca) stands. J. Ecol. 2009, 97, 1062–1074. [Google Scholar] [CrossRef]
- Druckenbrod, D.L.; Shugart, H.H.; Davies, I. Spatial pattern and process in forest stands within the Virginia piedmont. J. Veg. Sci. 2005, 16, 37–48. [Google Scholar] [CrossRef]
- Manso, R.; Morneau, F.; Ningre, F.; Fortin, M. Effect of climate and intra-and inter-specific competition on diameter increment in beech and oak stands. Forestry 2015, 88, 540–551. [Google Scholar] [CrossRef]
- Adler, P.B.; Smull, D.; Beard, K.H.; Choi, R.T.; Furniss, T.; Kulmatiski, A.; Meiners, J.M.; Tredennick, A.T.; Veblen, K.E. Competition and coexistence in plant communities: Intraspecific competition is stronger than interspecific competition. Ecol. Lett. 2018, 21, 1319–1329. [Google Scholar] [CrossRef] [Green Version]
- Dietze, M.C.; Wolosin, M.S.; Clark, J.S. Capturing diversity and interspecific variability in allometries: A hierarchical approach. For. Ecol. Manag. 2008, 256, 1939–1948. [Google Scholar] [CrossRef]
- Mina, M.; Huber, M.O.; Forrester, D.I.; Thürig, E.; Rohner, B. Multiple factors modulate tree growth complementarity in Central European mixed forests. J. Ecol. 2018, 106, 1106–1119. [Google Scholar] [CrossRef]
- Eis, S.; Craigdallie, D.; Simmons, C. Growth of lodgepole pine and white spruce in the central interior of British Columbia. Can. J. For. Res. 1982, 12, 567–575. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Tree Species | Block C | Block D | ||||||
|---|---|---|---|---|---|---|---|---|---|
| DBH (cm) | DBH (cm) | ||||||||
| n | Mean | Min | Max | n | Mean | Min | Max | ||
| 1993 | Douglas-fir | 861 | 8.7 | 0.2 | 37.6 | 725 | 9.0 | 0.3 | 51.5 |
| Spruce | 17 | 12.4 | 2.0 | 23.5 | 237 | 10.1 | 1.0 | 27.5 | |
| Lodgepole pine | 80 | 11.7 | 0.6 | 31.1 | 69 | 15.0 | 5.5 | 41.1 | |
| Broadleaf (aspen and birch) | 140 | 6.5 | 0.3 | 20.2 | 74 | 8.4 | 0.3 | 46.1 | |
| 1997 | Douglas-fir | 862 | 9.2 | 0.2 | 38.5 | 716 | 9.6 | 0.6 | 52.5 |
| Spruce | 17 | 13.7 | 3.1 | 25.5 | 225 | 10.8 | 0.9 | 28.5 | |
| Lodgepole pine | 76 | 13.0 | 0.4 | 32.2 | 68 | 17.1 | 5.6 | 41.5 | |
| Broadleaf (aspen and birch) | 119 | 7.0 | 0.3 | 21.0 | 76 | 7.3 | 0.2 | 47.0 | |
| 2004 | Douglas-fir | 840 | 10.3 | 0.1 | 39.5 | 679 | 10.7 | 0.3 | 54.2 |
| Spruce | 17 | 15.5 | 4.8 | 27.3 | 196 | 12.2 | 1.4 | 30.0 | |
| Lodgepole pine | 65 | 11.9 | 0.4 | 30.9 | 64 | 13.4 | 5.7 | 32.4 | |
| Broadleaf (aspen and birch) | 86 | 8.4 | 0.1 | 22.5 | 50 | 9.6 | 0.1 | 49.0 | |
| 2008 | Douglas-fir | 829 | 10.6 | 0.2 | 40.9 | 650 | 11.6 | 0.6 | 54.6 |
| Spruce | 17 | 16.0 | 0.8 | 28.2 | 173 | 12.9 | 0.6 | 31.3 | |
| Lodgepole pine | 34 | 6.4 | 0.3 | 22.0 | 10 | 14.0 | 5.7 | 22.3 | |
| Broadleaf (aspen and birch) | 90 | 8.4 | 0.1 | 23.4 | 42 | 9.9 | 0.1 | 49.4 | |
| 2013 | Douglas-fir | 760 | 11.9 | 0.5 | 42.5 | 616 | 12.4 | 0.6 | 55.1 |
| Spruce | 17 | 17.2 | 1.4 | 29.3 | 149 | 14.3 | 0.9 | 32.8 | |
| Lodgepole pine | 21 | 7.5 | 0.3 | 17.0 | 9 | 15.1 | 5.6 | 23.4 | |
| Broadleaf (aspen and birch) | 72 | 10.3 | 0.2 | 23.6 | 33 | 11.4 | 0.1 | 50.5 | |
| Source | Equation |
|---|---|
| Non-spatially explicit competition indices | |
| Steneker and Jarvis [31] | |
| Lorimer [32] | |
| Glover and Hool [33] | |
| Krajicek et al. [34] | |
| Wykoff et al. [35] | |
| Spatially explicit competition indices | |
| Hegyi [36] | |
| Martin and Ek [30] | |
| Rouvinen and Kuuluvainen [37] | |
| Rouvinen and Kuuluvainen [37] | |
| Braathe [38] | |
| Opie [39] | |
| Bella [40] | |
| Monserud and Ek [41] |
| Measurement Period | Block C | Block D | |||||
|---|---|---|---|---|---|---|---|
| Variable | FD | FD | SX | PL | |||
| n = 381 | n = 280 | n = 325 | n = 263 | n = 92 | n =58 | ||
| small trees | large trees | small trees | large trees | large trees | large trees | ||
| 1993 | DBH | 6.4 (2.1) | 14.9 (5.0) | 6.4 (2.3) | 14.4 (4.9) | 15.6 (3.6) | 16.8 (3.7) |
| BA | 35.9 (20.4) | 192.8 (158.5) | 36.3 (22.8) | 182.5 (174.2) | 200.9 (100.6) | 231.1 (111.3) | |
| 1997 | DBH | 7.0 (2.4) | 16.2 (5.0) | 6.8 (2.5) | 15.4 (5.1) | 16.3 (3.8) | 17.8 (3.9) |
| BA | 42.7 (25.5) | 224.7 (167.4) | 41.5 (26.9) | 206.9 (184.6) | 219.2 (110.0) | 261.5 (122.9) | |
| PABAI | 1.7 (1.7) | 8.0 (4.1) | 1.3 (1.4) | 6.1 (3.7) | 4.6 (3.4) | 7.6 (4.8) | |
| 2004 | DBH | 7.6 (2.7) | 17.9 (5.2) | 7.4 (2.9) | 16.9 (5.4) | 17.3 (4.2) | 19.1 (4.2) |
| BA | 51.7 (33.4) | 274.1 (186.7) | 49.3 (34.5) | 247.0 (205.0) | 248.4 (127.3) | 300.1 (138.9) | |
| PABAI | 1.3 (1.4) | 7.1 (4.1) | 1.1 (1.4) | 5.7 (3.8) | 4.2 (3.1) | 5.5 (3.5) | |
| 2008 | DBH | 8.0 (3.0) | 18.9 (5.4) | 7.7 (3.2) | 17.8 (5.6) | 17.9 (4.5) | |
| BA | 56.5 (39.0) | 303.3 (198.4) | 54.3 (41.2) | 274.2 (215.9) | 266.8 (140.3) | ||
| PABAI | 1.2 (1.8) | 7.3 (4.7) | 1.3 (2.2) | 6.8 (4.8) | 4.6 (5.2) | ||
| 2013 | DBH | 8.3 (3.3) | 20.1 (5.7) | 8.1 (3.5) | 19.0 (5.8) | 18.5 (4.8) | |
| BA | 63.0 (46.3) | 343.2 (218.4) | 60.7 (51.2) | 309.1 (229.5) | 286 (156.4) | ||
| PABAI | 1.1 (1.5) | 6.7 (4.5) | 1.1 (2.1) | 5.8 (4.3) | 3.2 (4.0) | ||
| Blocks | Species | Tree Size | Equation (7) | Equation (8) | Equation (9) | |
|---|---|---|---|---|---|---|
| Block C | FD | small | RMSE | 0.048 | 0.046 | 0.047 |
| AIC | 3050.8 | 2893.2 | 2837.1 | |||
| LRT | 165.6* | 80.1* | ||||
| Marginal | 0.56 | 0.71 | 0.75 | |||
| Conditional | 0.72 | 0.74 | 0.76 | |||
| large | RMSE | 0.108 | 0.076 | 0.074 | ||
| AIC | 5417.0 | 5215.2 | 5218.5 | |||
| LRT | 209.9* | 20.6 | ||||
| Marginal | 0.43 | 0.59 | 0.60 | |||
| Conditional | 0.55 | 0.60 | 0.61 | |||
| Block D | FD | small | RMSE | 0.023 | 0.024 | 0.024 |
| AIC | 2207.9 | 2140.9 | 2149.8 | |||
| LRT | 75.03* | 15.05 | ||||
| Marginal | 0.55 | 0.66 | 0.67 | |||
| Conditional | 0.78 | 0.80 | 0.80 | |||
| large | RMSE | 0.053 | 0.034 | 0.038 | ||
| AIC | 4580.1 | 4421.7 | 4415.24 | |||
| LRT | 166.37 * | 30.49* | ||||
| Marginal | 0.54 | 0.61 | 0.62 | |||
| Conditional | 0.65 | 0.69 | 0.71 | |||
| SX | large | RMSE | 0.003 | 0.003 | 0.002 | |
| AIC | 1487.2 | 1468.4 | 1475.0 | |||
| LRT | 26.7* | 17.4 | ||||
| Marginal | 0.42 | 0.50 | 0.48 | |||
| Conditional | 0.73 | 0.76 | 0.77 | |||
| PL | large | RMSE | 0.030 | 0.031 | 0.063 | |
| AIC | 592.69 | 587.97 | 589.56 | |||
| LRT | 8.72* | 10.41 | ||||
| Marginal | 0.37 | 0.50 | 0.54 | |||
| Conditional | 0.53 | 0.56 | 0.61 | |||
| Block C | Block D | |||||
|---|---|---|---|---|---|---|
| Parameter | Small FD | Large FD | Small FD | Large FD | Large SX | Large PL |
| Fixed effects | ||||||
| (Ref) | 0.133 (0.038) | 0.244 (0.099) | 0.063 (0.027) | 0.066 (0.028) | 0.001 (0.002) | 0.017 (0.040) |
| −0.011 (0.065) | −0.230 (0.099) | 0.053 (0.061) | −0.047 (0.030) | 0.000 (0.003) | −0.017 (0.040) | |
| −0.097 (0.048) | −0.234 (0.099) | 0.003 (0.049) | −0.064 (0.028) | −0.001 (0.002) | ||
| −0.126 (0.039) | −0.240 (0.099) | 0.129 (0.111) | −0.066 (0.028) | −0.001 (0.003) | ||
| (Ref) | 1.650 (0.207) | 1.607 (0.176) | 1.900 (0.287) | 1.950 (0.180) | 3.358 (0.763) | 2.468 (0.953) |
| −0.413 (0.349) | 0.968 (0.318) | −0.790 (0.410) | 0.380 (0.287) | −0.143 (1.184) | 1.587 (1.997) | |
| 0.161 (0.486) | 1.034 (0.369) | −0.729 (0.447) | 1.316 (0.350) | 0.710 (1.286) | ||
| 0.721 (0.527) | 1.165 (0.409) | −1.444 (0.398) | 1.602 (0.429) | −0.231 (1.582) | ||
| (Ref) | −0.213 (0.234) | 0.124 (0.029) | 0.012 (0.310) | 0.079 (0.028) | 0.252 (0.134) | 0.158 (0.139) |
| −0.480 (0.342) | 0.056 (0.046) | −0.892 (0.424) | 0.014 (0.042) | −0.090 (0.194) | 0.167 (0.268) | |
| −0.210 (0.336) | 0.046 (0.047) | −1.079 (0.379) | 0.127 (0.048) | 0.011 (0.188) | ||
| 0.049 (0.329) | −0.004 (0.045) | −1.211 (0.349) | 0.147 (0.054) | −0.182 (0.205) | ||
| (Ref) | −0.217 (0.015) | −0.250 (0.018) | −0.164 (0.028) | −0.153 (0.025) | −0.031 (0.060) | −0.126 (0.079) |
| 0.049 (0.022) | 0.068 (0.026) | 0.040 (0.040) | 0.012 (0.038) | 0.006 (0.086) | 0.079 (0.117) | |
| 0.006 (0.031) | 0.043 (0.029) | 0.033 (0.044) | −0.005 (0.037) | −0.142 (0.084) | ||
| 0.153 (0.032) | 0.144 (0.031) | 0.034 (0.048) | −0.001 (0.042) | −0.058 (0.118) | ||
| (Ref) | 0.003 (0.064) | −0.020 (0.113) | −0.182 (0.037) | −0.216 (0.044) | −0.128 (0.048) | −0.338 (0.253) |
| −0.037 (0.112) | −0.180 (0.193) | −0.001 (0.054) | 0.002 (0.058) | 0.056 (0.070) | 0.301 (0.338) | |
| −0.157 (0.122) | −0.015 (0.180) | 0.038 (0.061) | 0.063 (0.055) | −0.126 (0.076) | ||
| −0.127 (0.157) | −0.236 (0.227) | 0.057 (0.063) | 0.196 (0.057) | 0.011 (0.117) | ||
| (Ref) | −0.065 (0.027) | −0.166 (0.047) | −0.222 (0.049) | −0.298 (0.050) | −0.378 (0.154) | −0.507 (0.163) |
| 0.049 (0.044) | 0.066 (0.074) | 0.063 (0.074) | 0.088 (0.071) | −0.156 (0.241) | 0.094 (0.282) | |
| 0.029 (0.058) | 0.229 (0.096) | 0.260 (0.073) | 0.248 (0.066) | 0.221 (0.185) | ||
| 0.434 (0.158) | 0.249 (0.251) | −0.102 (0.333) | 0.007 (0.296) | −0.939 (2.114) | ||
| (Ref) | 0.023 (0.032) | −0.205 (0.060) | −0.164 (0.060) | −0.195 (0.057) | −0.274 (0.133) | −1.096 (0.674) |
| 0.067 (0.060) | 0.083 (0.099) | −0.023 (0.106) | 0.079 (0.081) | 0.007 (0.217) | 0.809 (0.909) | |
| 0.032 (0.078) | 0.193 (0.109) | −0.160 (0.179) | 0.038 (0.087) | 0.006 (0.254) | ||
| 0.066 (0.087) | 0.184 (0.127) | −0.048 (0.193) | 0.120 (0.101) | −0.105 (0.451) | ||
| Variance component | ||||||
| σ2 | 0.0022 | 0.0055 | 0.0007 | 0.0014 | 0.0000 | 0.0040 |
| 0.0054 | 0.0005 | 0.0116 | 0.0054 | 0.0164 | 0.0029 | |
| Power | 1.3278 | 1.2473 | 1.6035 | 1.4291 | 2.3180 | 1.3126 |
| Block C | Block D | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Small FD | Large FD | Small FD | Large FD | Large SX | Large PL | |||||||
| F-Value | P-Value | F-Value | P-Value | F-Value | P-Value | F-Value | P-Value | F-Value | P-Value | F-Value | P-Value | |
| 3353.8 | <0.0001 | 39966.7 | <0.0001 | 1545.80 | <0.0001 | 20993.31 | <0.0001 | 1489.95 | <0.0001 | 6005.46 | <0.0001 | |
| 1293.3 | <0.0001 | 15448.4 | <0.0001 | 144.10 | <0.0001 | 4128.48 | <0.0001 | 863.38 | <0.0001 | 6266.03 | <0.0001 | |
| 1494.6 | <0.0001 | 757.85 | <0.0001 | 1285.22 | <0.0001 | 1266.56 | <0.0001 | 313.76 | <0.0001 | 73.84 | <0.0001 | |
| 12.9 | <0.0001 | 14.12 | <0.0001 | 9.38 | <0.0001 | 3.66 | 0.0122 | 2.08 | 0.1027 | 0.06 | 0.8113 | |
| 8.0 | 0.0049 | 79.35 | <0.0001 | 32.49 | <0.0001 | 68.21 | <0.0001 | 10.71 | 0.0012 | 2.91 | 0.0912 | |
| 1.9 | 0.1329 | 1.53 | 0.2062 | 2.46 | 0.0615 | 4.38 | 0.0045 | 0.95 | 0.4186 | 0.33 | 0.5676 | |
| 221.6 | <0.0001 | 228.07 | <0.0001 | 27.09 | <0.0001 | 100.36 | <0.0001 | 0.19 | 0.6636 | 0.00 | 0.9657 | |
| 7.6 | <0.0001 | 8.33 | <0.0001 | 0.58 | 0.625 | 1.08 | 0.3565 | 0.86 | 0.4628 | 0.63 | 0.4306 | |
| 0.9 | 0.3344 | 1.34 | 0.2464 | 23.13 | <0.0001 | 16.85 | <0.0001 | 10.62 | 0.0012 | 2.62 | 0.1091 | |
| 1.0 | 0.4128 | 0.62 | 0.6033 | 0.10 | 0.9624 | 2.44 | 0.0629 | 3.02 | 0.0299 | 1.32 | 0.2542 | |
| 4.9 | 0.0276 | 7.61 | 0.0059 | 12.95 | 0.0003 | 28.79 | <0.0001 | 8.68 | 0.0034 | 11.47 | 0.0010 | |
| 3.2 | 0.0227 | 2.14 | 0.0938 | 4.42 | 0.0042 | 4.87 | 0.0023 | 1.13 | 0.3369 | 0.15 | 0.6996 | |
| 3.0 | 0.0835 | 8.95 | 0.0028 | 11.36 | 0.0008 | 14.63 | 0.0001 | 7.83 | 0.0054 | 1.88 | 0.1732 | |
| 0.5 | 0.6632 | 1.42 | 0.24 | 0.33 | 0.8012 | 0.60 | 0.6145 | 0.02 | 0.9961 | 0.79 | 0.3757 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acquah, S.B.; Marshall, P.L. Assessing Differences in Competitive Effects Among Tree Species in Central British Columbia, Canada. Forests 2020, 11, 167. https://doi.org/10.3390/f11020167
Acquah SB, Marshall PL. Assessing Differences in Competitive Effects Among Tree Species in Central British Columbia, Canada. Forests. 2020; 11(2):167. https://doi.org/10.3390/f11020167
Chicago/Turabian StyleAcquah, Stella Britwum, and Peter L. Marshall. 2020. "Assessing Differences in Competitive Effects Among Tree Species in Central British Columbia, Canada" Forests 11, no. 2: 167. https://doi.org/10.3390/f11020167