Demonstrating the Effect of Height Variation on Stand-Level Optimization with Diameter-Structured Matrix Model



Abstract
:1. Introduction
2. Material and Methods
2.1. The Optimization Problem
2.2. Model Estimation and Data
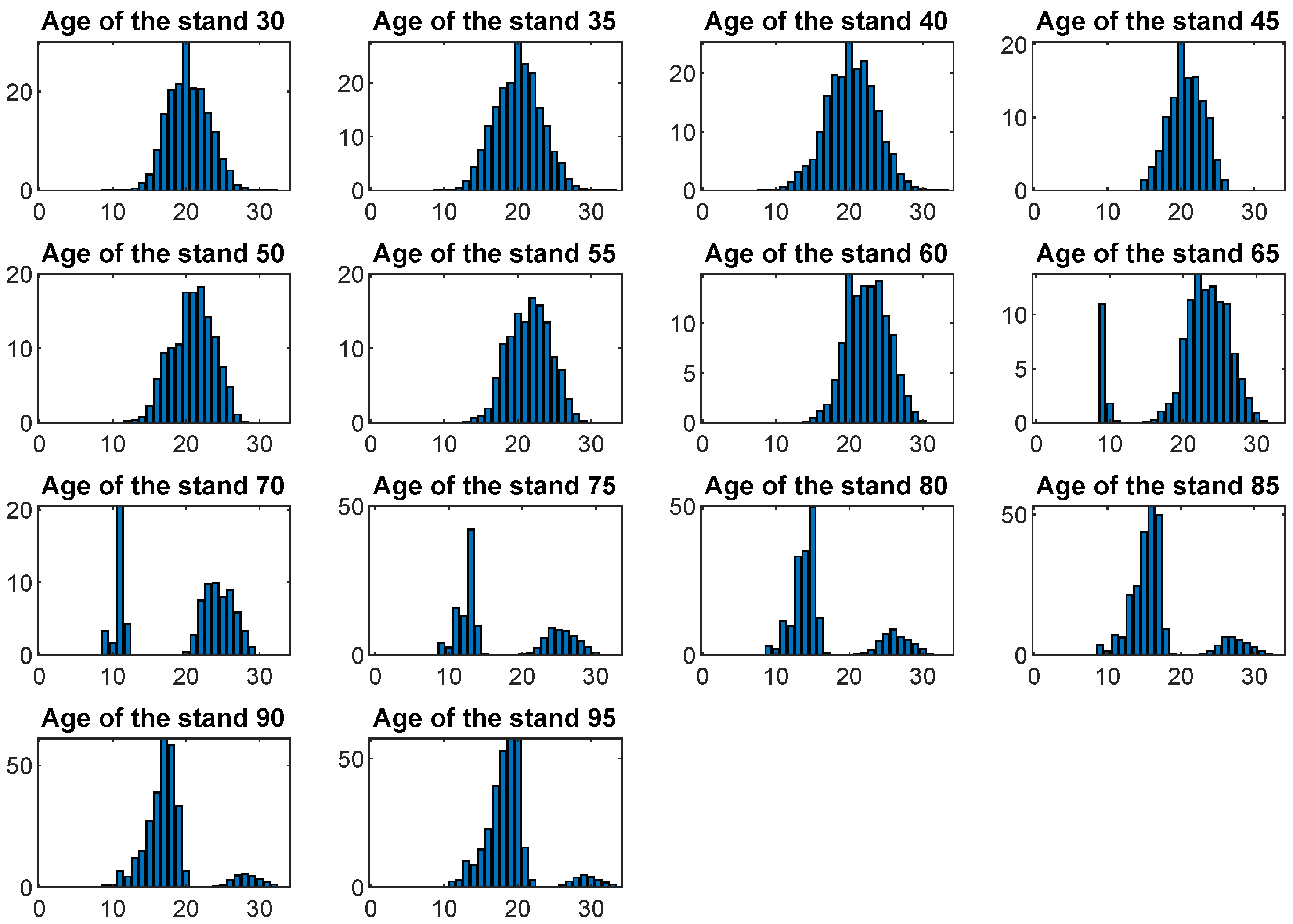
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Xie, Q.; He, Z.-R.; Wandg, X. Optimal harvesting in diffusive population models with size random growth and distributed recruitment. Electron. J. Differ. Eq. 2016, 214, 1–13. [Google Scholar]
- Goetz, R.U.; Xabadia, A.; Calvo, E. Optimal forest management in the presence of intraspecific competition. Math. Popul. Stud. 2016, 18, 151–171. [Google Scholar] [CrossRef]
- Lefkovitch, L. The study of population growth in organisms grouped by stages. Biometrics 1965, 21, 1–18. [Google Scholar] [CrossRef]
- Caswell, H. Matrix Population Models: Construction, Analysis and Interpretation, 2nd ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2001. [Google Scholar]
- Picard, N.; Mortier, F.; Chagneau, P. Influence of estimations of the vital rates in the stock recovery rate using matrix models for tropical rainforests. Ecol. Model. 2008, 214, 349–360. [Google Scholar] [CrossRef]
- Liang, J.; Picard, N. Matrix model of forest dynamics: An overview and Outlook. For. Sci. 2013, 59, 359–378. [Google Scholar] [CrossRef]
- Picard, N.; Liang, J. Matrix models for size-structured populations: Unrealistic fast growth or simply diffusion? PLoS ONE 2014, 9, e98254. [Google Scholar] [CrossRef]
- Liang, J.; Zhou, M. A geospatial model of forest dynamics with controlled trend surface. Ecol. Model. 2010, 221, 2339–2352. [Google Scholar] [CrossRef]
- Liang, J.; Zhou, M.; Verbyla, D.; Zhang, L.; Springsteen, A.L.; Malone, T. Mapping forest dynamics under climate change: A matrix model. For. Ecol. Manag. 2011, 262, 2250–2262. [Google Scholar] [CrossRef]
- Hu, H.; Wang, S.; Guo, Z.; Xu, B.; Fang, J. The stage-classified matrix models project a significant increase in biomass carbon stocks in China’s forests between 2005 and 2050. Sci. Rep. 2015, 5, 11203. [Google Scholar] [CrossRef] [Green Version]
- Pihlainen, S.; Tahvonen, O.; Niinimäki, S. The economics of timber and bioenergy production and carbon storage in Scots pine stands. Can. J. For. Res. 2014, 44, 1091–1102. [Google Scholar] [CrossRef]
- Assmuth, A.; Rämö, J.; Tahvonen, O. Economics of size-structured forestry with carbon storage. Can. J. For. Res. 2018, 48, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Pyy, J.; Ahtikoski, A.; Laitinen, E.; Siipilehto, J. Introducing a Non-Stationary Matrix Model for Stand-Level Optimization, an Even-Aged Pine (Pinus sylvestris L.) Stand in Finland. Forests 2017, 8, 163. [Google Scholar] [CrossRef] [Green Version]
- Pyy, J.; Ahtikoski, A.; Lapin, A.; Laitinen, E. Solution of Optimal Harvesting Problem by Finite Difference Approximations of Size-Structured Population Model. Math. Comp. Appl. 2018, 23, 22. [Google Scholar] [CrossRef] [Green Version]
- Cao, T.; Valsta, L.; Mäkelä, A. A comparison of carbon assessment methods for optimizing timber production and carbon sequestration in Scots pine stands. For. Ecol. Manag. 2010, 260, 1726–1734. [Google Scholar] [CrossRef]
- Niinimäki, S.; Tahvonen, O.; Mäkelä, A. Applying a process-based model in Norway spruce management. For. Ecol. Manag. 2012, 265, 102–115. [Google Scholar] [CrossRef]
- Tahvonen, O.; Pihlainen, S.; Niinimäki, S. On the economics of optimal timber production in boreal Scots pine stands. Can. J. For. Res. 2013, 43, 719–730. [Google Scholar] [CrossRef]
- Hurttala, H.; Cao, T.; Valsta, L. Optimization of Scots pine (Pinus sylvestris) management with the total net return from the value chain. J. For. Econ. 2017, 28, 1–11. [Google Scholar] [CrossRef]
- Pohjola, J.; Valsta, L. Carbon credits and management of Scots pine and Norway spruce stands in Finland. For. Policy Econ. 2007, 9, 789–798. [Google Scholar] [CrossRef]
- Ahtikoski, A.; Salminen, H.; Ojansuu, R.; Hynynen, J.; Kärkkäinen, K.; Haapanen, M. Optimizing stand management involving the effect of genetic gain: Preliminary results for Scots pine in Finland. Can. J. For. Res. 2013, 43, 1–7. [Google Scholar] [CrossRef]
- Juutinen, A.; Ahtikoski, A.; Lehtonen, M.; Mäkipää, R.; Ollikainen, M. The impact of a short-term carbon payment scheme on forest management. For. Policy Econ. 2018, 90, 115–127. [Google Scholar] [CrossRef]
- Grimm, V. Ten years of individual-based modelling in ecology: What have we learned and what could we learn in the future? Ecol. Model. 1999, 115, 129–148. [Google Scholar] [CrossRef]
- Zuidema, P.A.; Jonsegas, E.; Chien, P.D.; During, H.J.; Schieving, F. Integral projection models for trees: A new parametrization method and a validation of model output. J. Ecol. 2010, 98, 345–355. [Google Scholar] [CrossRef]
- Sable, S.E.; Rose, K.A. A comparison of individual-based and matrix projection models for simulating yellow perch population dynamics in Oneida Lake, New York, USA. Ecol. Model. 2008, 215, 105–121. [Google Scholar] [CrossRef]
- Haapanen, M.; Hynynen, J.; Ruotsalainen, S.; Siipilehto, J.; Kilpeläinen, M.-L. Realised and projected gains in growth, quality and simulated yield of genetically improved Scots pine in southern Finland. Eur. J. For. Res. 2016, 135, 997–1009. [Google Scholar] [CrossRef]
- Ahtikoski, A.; Haapanen, M.; Hynynen, J.; Karhu, J.; Kärkkäinen, K. Genetically improved reforestation stock provides simultaneous benefits for growers and a sawmill, a case study in Finland. Scand. J. For. Res. 2018, 33, 484–492. [Google Scholar] [CrossRef]
- Lyhykäinen, H.T.; Mäkinen, H.; Mäkelä, A.; Pastila, S.; Heikkilä, A.; Usenius, A. Predicting lumber grade and by-product yields for Scots pine trees. For. Ecol. Manag. 2009, 258, 146–158. [Google Scholar] [CrossRef]
- Ojansuu, R.; Mäkinen, H.; Heinonen, J. Including variation in branch and tree properties improves timber grade estimates in Scots pine stands. Can. J. For. Res. 2018, 48, 542–553. [Google Scholar] [CrossRef]
- Mäkinen, H. Effect of intertree competition on branch characteristics of Pinus sylvestris families. Scand. J. For. Res. 1996, 11, 129–136. [Google Scholar] [CrossRef]
- Gort, J.; Zubizarret-Gerendiain, A.; Peltola, H.; Kilpeläinen, A.; Pulkkinen, P.; Jaatinen, R.; Kellomäki, S. Differences in branch characteristics of Scots pine (Pinus sylvestris L.) genetic entries grown at different spacing. Ann. For. Sci. 2010, 67, 70. [Google Scholar] [CrossRef] [Green Version]
- Malinen, J.; Kilpeläinen, H.; Verkasalo, E. Validating the predicted saw log and pulpwood proportions and gross value of Scots pine and Norway spruce harvest at stand level by Most Similar Neighbour analyses and a stem quality database. Silva Fenn. 2018, 52. [Google Scholar] [CrossRef]
- Mäkinen, H.; Isomäki, A. Thinning intensity and growth of Scots pine stands in Finland. For. Ecol. Manag. 2004, 201, 311–325. [Google Scholar] [CrossRef]
- Siipilehto, J.; Kangas, A. Näslundin pituuskäyrä ja siihen perustuvia malleja läpimitan ja pituuden välisestä riippuvuudesta suomalaisissa talousmetsissä. MetsäTieteen Aikakauskirja 2015, 4, 215–236. [Google Scholar]
- Laasasenaho, J. Taper curve and volume functions for pine, spruce and birch. Commun. Lnst. For. Fenn. 1982, 108, 1–74. [Google Scholar]
- Hynynen, J.; Ojansuu, R.; Hökkä, H.; Siipilehto, J.; Salminen, H.; Haapala, P. Models for predicting stand development in MELA System. Metsäntutkimus laitoksen tiedonantoja. Finn. For. Res. Inst. Res. Pap. 2002, 835, 31–35. [Google Scholar]
- Xue, H.; Mäkelä, A.; Valsta, L.; Vanclay, J.K.; Cao, T. Comparison of population-based algorithms for optimizing thinnings and rotation using a process-based growth model. Scand. J. For. Res. 2019. [Google Scholar] [CrossRef] [Green Version]
- Mäkelä, A.; del Rìo, M.; Hynynen, J.; Hawkins, M.J.; Reyer, C.; Soares, P.; van Oijen, M.; Tomé, M. Using stand-scale forest models for estimating indicators of sustainable forest management. For. Ecol. Manag. 2012, 285, 164–178. [Google Scholar] [CrossRef] [Green Version]
- Pukkala, T. Population-based methods in the optimization of stand management. Silva Fenn. 2009, 43, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Anita, S.; Arnautu, V.; Stefanescu, R. Numerical optimal harvesting for a periodic age-structured population dynamics with logistic term. Numer. Funct. Anal. Optim. 2009, 30, 183–198. [Google Scholar] [CrossRef]
- Ikonen, V.-P.; Kellomäki, S.; Peltola, H. Sawn timber properties of Scots pine as affected by initial stand density, thinning and pruning: A simulation based approach. Silva Fenn. 2009, 43, 411–431. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Torres, I.; Perez, S.O.; Robredo, F.G.; Belda, C.F. Optimizing the management of uneven-aged Pinus nigra stands between two stable positions. iFor. Biosci. For. 2016, 9, 599–607. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
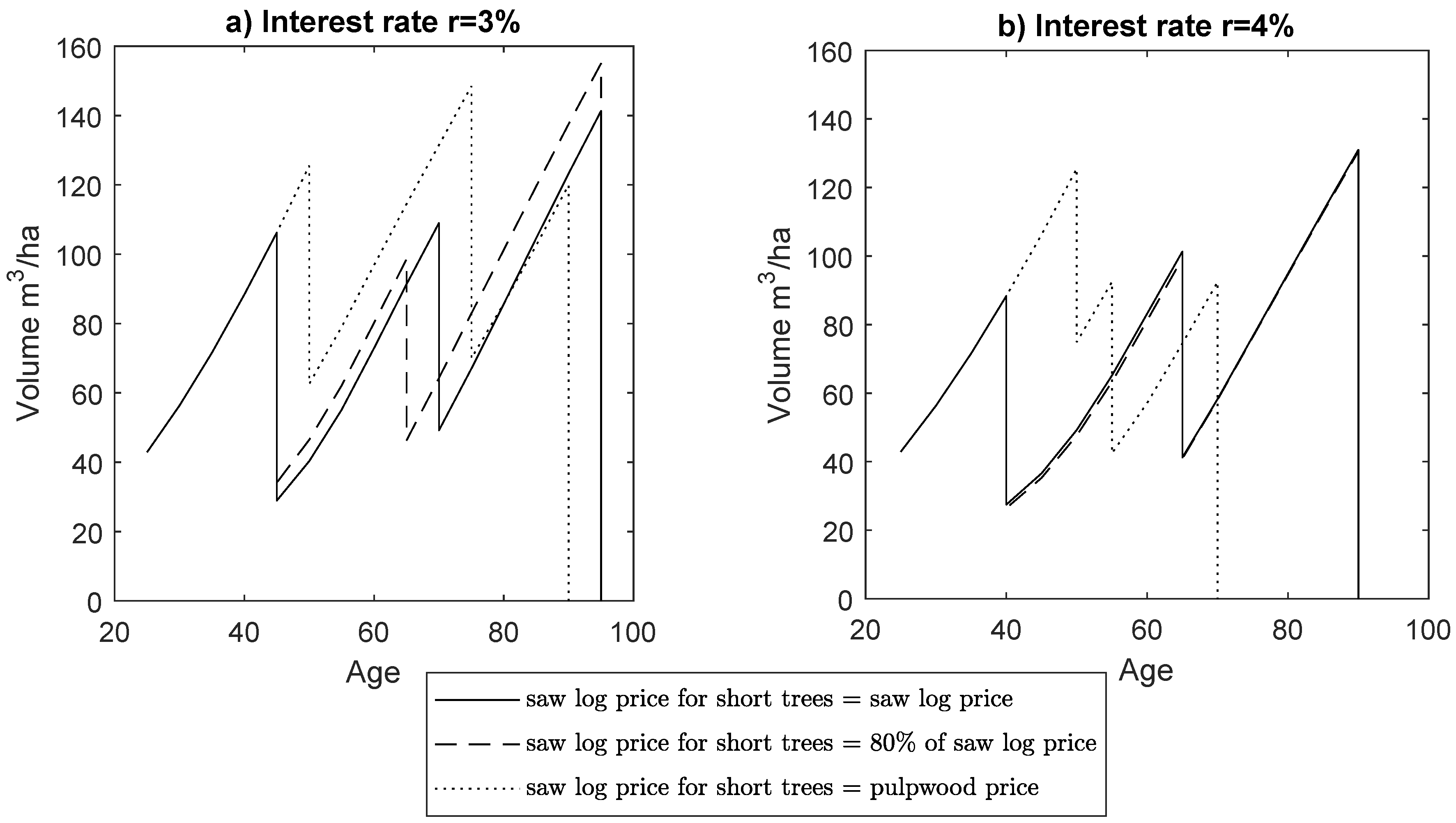
| Thinning | Stand Age (a) | Removal (m ha) | Thinning Intensity (% of Basal Area Removed) | Saw Log Proportion (%) | MaxNPV (€ha) | MAI (m ha a) |
|---|---|---|---|---|---|---|
| // | // | // | c/ / | // | // | |
| Interest rate | ||||||
| 1st | 45/45/50 | 77.3/72.1/62.9 | 54/50/35 | 60/61/67 | 3059/2872/2582 | 2.93/2.94/2.90 |
| 2nd | 70/65/75 | 59.8/52.3/78.5 | 33/27/42 | 64/64/72 | ||
| clearcut | 95/95/90 | 141.4/155.1/120.0 | 100/100/100 | 39/42/50 | ||
| total | 278.5/279.5/261.4 | 50/51/61 | ||||
| Interest rate | ||||||
| 1st | 40/40/50 | 60.9/61.9/50.6 | 61/62/28 | 53/52/71 | 2169/2036/1760 | 2.80/2.79/2.76 |
| 2nd | 65/65/55 | 60.1/58.4/50.0 | 32/31/32 | 64/64/60 | ||
| clearcut | 90/90/70 | 131.0/130.5/92.4 | 100/100/100 | 35/35/38 | ||
| total | 252.0/250.9/193.0 | 46/46/52 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pyy, J.; Laitinen, E.; Ahtikoski, A. Demonstrating the Effect of Height Variation on Stand-Level Optimization with Diameter-Structured Matrix Model. Forests 2020, 11, 226. https://doi.org/10.3390/f11020226
Pyy J, Laitinen E, Ahtikoski A. Demonstrating the Effect of Height Variation on Stand-Level Optimization with Diameter-Structured Matrix Model. Forests. 2020; 11(2):226. https://doi.org/10.3390/f11020226
Chicago/Turabian StylePyy, Johanna, Erkki Laitinen, and Anssi Ahtikoski. 2020. "Demonstrating the Effect of Height Variation on Stand-Level Optimization with Diameter-Structured Matrix Model" Forests 11, no. 2: 226. https://doi.org/10.3390/f11020226
APA StylePyy, J., Laitinen, E., & Ahtikoski, A. (2020). Demonstrating the Effect of Height Variation on Stand-Level Optimization with Diameter-Structured Matrix Model. Forests, 11(2), 226. https://doi.org/10.3390/f11020226