Ease of Access to An Alternative Food Source Enables Wallabies to Strip Bark in Tasmanian Pinus radiata Plantations


Abstract
:1. Introduction
2. Materials and Methods
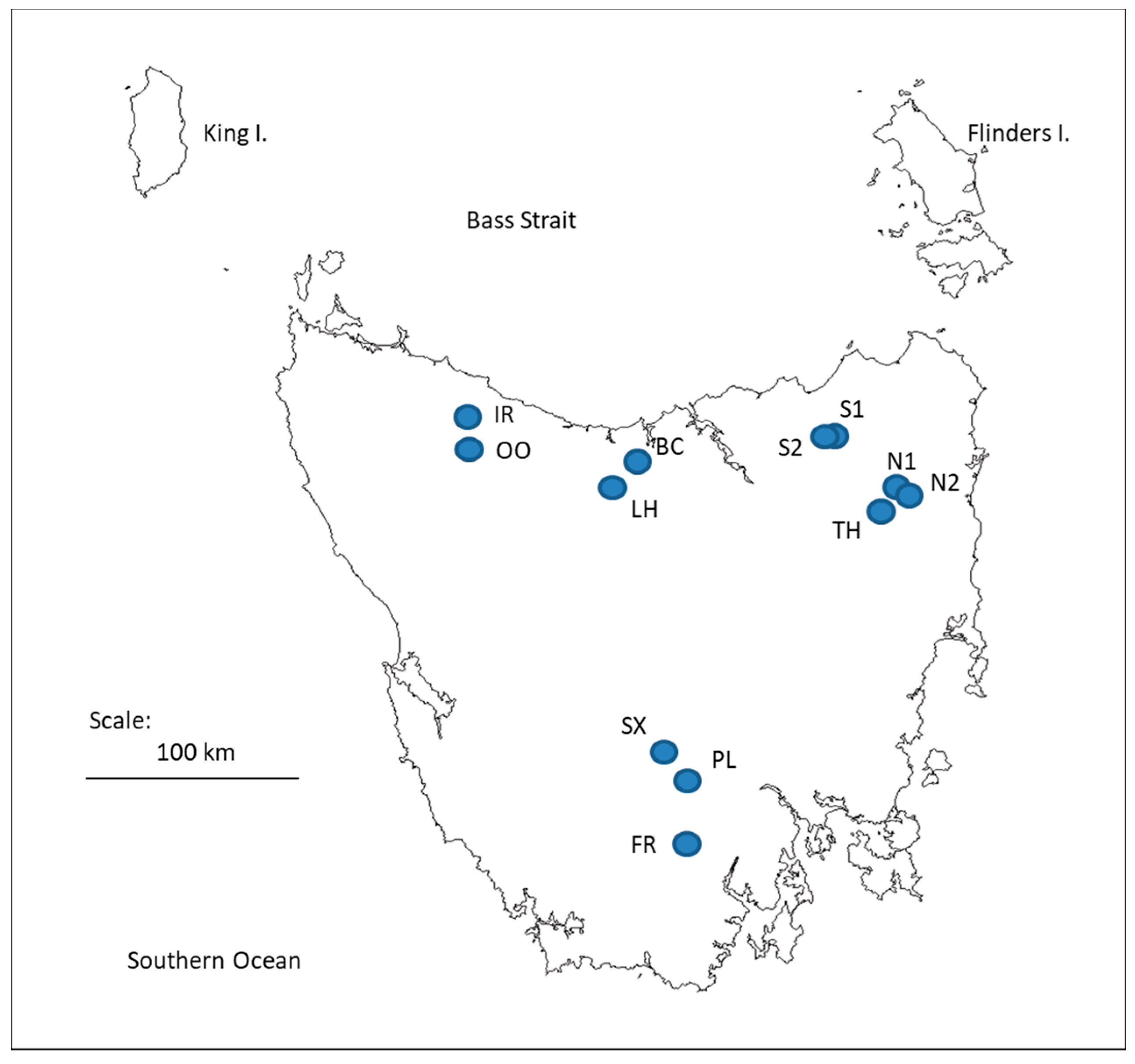
2.1. Study Sites
2.2. Response Variables
2.3. Explanatory Variables
2.4. Statistical Analysis
3. Results
3.1. Percentage Girdling at the Plot Level (n = 60)
3.2. Percentage Girdling at the Site Level (n = 12)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mead, D.J. Sustainable Management of Pinus radiata Plantations; FAO Forestry Paper No. 170; FAO: Rome, Italy, 2013. [Google Scholar]
- McNally, J. Damage to Victorian exotic pine plantations by native animals. Aust. For. 1955, 19, 87–99. [Google Scholar] [CrossRef]
- Wotherspoon, K. An Analysis of the Factors Influencing Bark Stripping by Browsing Mammals in Pinus radiata Plantations; Division of Forestry Research and Development, Forestry Tasmania: Hobart, Australia, 2004; p. 28, unpublished confidential client report. [Google Scholar]
- Department of Primary Industries, Parks, Water and the Environment, Tasmanian Government. 2014. Available online: http://dpipwe.tas.gov.au/wildlife-management/living-with-wildlife/living-with-kangaroos-and-wallabies (accessed on 26 May 2017).
- Calaby, J.H. Marsupials. In The Complete Book of Australian Mammals; Strahan, R., Ed.; Angus & Robertson: Sydney, Australia, 1983; pp. 239–241. [Google Scholar]
- Wardlaw, T. A Manual of Health Problems Affecting Pinus radiata in Tasmania; Technical Report 12/2003; Forestry Tasmania, Division of Forest Research and Development: Hobart, Australia, 2003. [Google Scholar]
- Sprent, J.; McArthur, C. Diet and diet selection of two species in the macropodid browser-grazer continuum—Do they eat what they ‘should’? Aust. J. Zool. 2002, 50, 183–192. [Google Scholar] [CrossRef]
- Pietrzykowski, E.; McArthur, C.; Fitzgerald, H.; Goodwin, A.N. Influence of patch characteristics on browsing of tree seedlings by mammalian herbivores. J. Appl. Ecol. 2003, 40, 458–469. [Google Scholar] [CrossRef]
- Le Mar, K.; McArthur, C. Comparison of habitat selection by two sympatric macropods, Thylogale billardierii and Macropus rufogriseus, in a patchy eucalypt-forestry environment. Austral Ecol. 2005, 30, 674–683. [Google Scholar] [CrossRef]
- Gill, R.M.A. A review of damage by mammals in North Temperate Forests: 2. Small mammals. Forestry 1992, 65, 281–308. [Google Scholar] [CrossRef]
- Díaz-Yáñez, O.; Mola-Yudego, B.; González-Olabarria, J.R. What variables make a forest stand vulnerable to browsing damage occurrence? Silva Fenn. 2017, 51. [Google Scholar] [CrossRef] [Green Version]
- Senn, J.; Suter, W. Ungulate browsing on silver fir (Abies alba) in the Swiss Alps: Beliefs in search of supporting data. For. Ecol. Manage. 2003, 181, 151–164. [Google Scholar] [CrossRef]
- Di Stephano, J. Mammalian browsing damage in the Mt. Cole State forest, southeastern Australia: Analysis of browsing patterns, spatial relationships and browse selection. New For. 2005, 29, 43–61. [Google Scholar] [CrossRef]
- Hazeltine, A.; Kirkpatrick, J.B. Practical and theoretical implications of a browsing cascade in Tasmanian forest and woodland. Aust. J. Bot. 2015, 63, 435–443. [Google Scholar] [CrossRef]
- Stutz, R.S.; Banks, R.B.; Dexter, N.; McArthur, C. Herbivore search behaviour drives associational plant refuge. Acta Oecol. 2015, 67, 1–7. [Google Scholar] [CrossRef]
- Wardlaw, T. The evolution of forest health surveillance in Tasmania. Tasforests 2008, 17, 57–66. [Google Scholar]
- Queensland Government. SILO Climate Data. Department of Science, Information Technology, Innovation and the Arts, The State of Queensland, Australia. Available online: http://www.longpaddock.qld.gov.au/ (accessed on 2 January 2014).
- Kobe, R.K. Carbohydrate allocation to storage as a basis of interspecific variation in sapling survivorship and growth. Oikos 1997, 80, 226–233. [Google Scholar] [CrossRef]
- Kaelke, C.M.; Dawson, J.O. The accretion of nonstructural carbohydrates changes seasonally in Alnus incana ssp. rugosa in accord with tissue type, growth, N allocation, and root hypoxia. Symbiosis 2005, 39, 61–66. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Multimodel inference: Understanding AIC and BIC in model selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Ball, J.P.; Dahlgren, J. Browsing damage on pine (Pinus sylvestris and Pinus contorta) by a migrating moose (Alces alces) population in winter: Relation to habitat composition and road barriers. Scand. J. For. Res. 2002, 17, 427–435. [Google Scholar] [CrossRef]
- Page, D.E.; Close, D.; Beadle, C.L.; Wardlaw, T.J.; Mohammed, C.L. Seasonal dynamics in understorey abundance and carbohydrate concentration in relation to browsing and bark stripping of Tasmanian Pinus radiata plantations. For. Ecol. Manage. 2013, 296, 98–107. [Google Scholar] [CrossRef]
- Levitt, J. Response of Plants to Environmental Stresses. Volume I. Chilling, Freezing and High Temperature Stresses; Academic Press: New York, NY, USA, 1980; 497p. [Google Scholar]
- Fernández, M.; Royo, A.; Gil, L.; Pardos, J.A. Effects of temperature on growth and stress hardening development of phytotron-grown seedlings of Aleppo pine (Pinus halepensis Mill.). Ann. For. Sci. 2003, 60, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Ögren, E. Effects of climatic warming on cold hardiness of some northern woody plants assessed from simulation experiments. Physiol. Plant. 2001, 112, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Ögren, E.; Nilsson, T.; Sundblad, L.G. Relationship between respiratory depletion of sugars and loss of cold hardiness in coniferous seedlings over-wintering at raised temperatures: Indications of different sensitivities of spruce and pine. Plant Cell Environ. 1997, 20, 247–253. [Google Scholar] [CrossRef]
- Repo, T.; Leinonen, I.; Ryyppö, A.; Finér, L. The effect of soil temperature on the bud phenology, chlorophyll fluorescence, carbohydrate content and cold hardiness of Norway spruce seedlings. Physiol. Plant. 2004, 121, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Milligan, H.T.; Koricheva, J. Effects of tree species richness and composition on moose winter browsing damage and foraging selectivity: An experimental study. J. Anim. Ecol. 2013, 82, 739–748. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Site Name | Lat. (N) | Long. (E) | Alt. (m) | Ave. Annual Rainfall (mm) | Ave. Annual Air Temp. (°C) | Ave. Min. Soil Temp. (Spring) (°C) | Ave. Min. Air Temp. (Spring) (°C) | Tdiff (Ave. Diff. of Min. Soil and Air Temps.) (°C) | Average Damage Score (%girdling) |
|---|---|---|---|---|---|---|---|---|---|
| Branchs Creek | −41.27 | 146.66 | 131 | 744 | 12.9 | 9.4 | 8.0 | 1.4 | 0.0 |
| Franklin | −43.06 | 146.88 | 293 | 1123 | 9.7 | 10.9 | 4.7 | 6.2 | 25.5 |
| Inglis River | −41.11 | 145.60 | 111 | 1353 | 11.3 | 8.8 | 6.2 | 2.6 | 22.9 |
| Longhill | −41.34 | 146.49 | 120 | 988 | 11.6 | 7.3 | 6.2 | 1.1 | 2.5 |
| Nicholas 1 | −41.45 | 147.97 | 338 | 915 | 10.6 | 8.1 | 5.1 | 3.0 | 16.1 |
| Nicholas 2 | −41.47 | 147.98 | 324 | 915 | 10.6 | 8.6 | 5.1 | 3.5 | 4.7 |
| Oonah | −41.23 | 145.62 | 454 | 1439 | 11.2 | 7.5 | 6.1 | 1.4 | 0.4 |
| Plenty | −42.87 | 146.89 | 427 | 876 | 9.2 | 8.0 | 4.0 | 4.0 | 16.7 |
| Springfield 1 | −41.21 | 147.63 | 311 | 785 | 13.0 | 8.4 | 7.3 | 1.1 | 8.7 |
| Springfield 2 | −41.21 | 147.61 | 294 | 785 | 13.0 | 9.9 | 7.3 | 2.6 | 21.8 |
| Styx | −42.77 | 146.83 | 539 | 714 | 11.6 | 8.0 | 5.6 | 2.4 | 1.3 |
| Tower Hill | −41.53 | 147.91 | 512 | 716 | 11.5 | 6.2 | 5.6 | 0.6 | 41.6 |
| Variable | Description | Units |
|---|---|---|
| Plot-specific: | ||
| %Gird | Cumulative damage score, percentage of bark removed | % |
| ang%Gird | Angular transformation of cumulative damage score %Gird | |
| BareGrd | Percentage of area as bare ground | % |
| BBM | Composite variable, = BareGrd+Bracken+Moss | % |
| BBMG | Composite variable, = BareGrd+Bracken+Moss+Grass | % |
| Bracken | Percentage of area as Pteridium esculentum | % |
| Grass | Percentage of area as grass | % |
| height | Average height of the trees in the plot | m |
| inter_1 | Length of first internode of tree | mm |
| inter_2 | Length of second internode of tree | mm |
| LiveMat | Percentage of area as live material (grasses, herbs, forbs, etc.) | % |
| Moss | Percentage of area as mosses and liverworts | % |
| P_radiata | Percentage of area containing wilding Pinus radiata | % |
| Rock | Percentage of area occupied by rock | % |
| RockWood | Composite variable, = Rock+WoodDeb | % |
| SoilTmin | Minimum soil temperature in spring months, iButton | °C |
| whorl_1 | No. of branches in first whorl of tree | integer |
| whorl_2 | No. of branches in second whorl of tree | integer |
| WoodDeb | Percentage of area as woody debris | % |
| Site-specific: | ||
| Tdiff | Difference between mean minimum soil and air temperatures in spring months, = SoilTmin-TminSpr | °C |
| TminSpr | Minimum air temperature in spring months (SILO) | °C |
| Regression Relationship | AIC | BIC | adj R2 |
|---|---|---|---|
| ang%Gird = 0.106 + 0.00326(BareGrd) + 0.0136(Bracken) + 0.00947(Moss) | −187.9 | −185.4 | 0.282 |
| ang%Gird = 0.09743 + 0.00500(BBM) | −187.8 | −185.6 | 0.257 |
| AIC | BIC | adjR2 | |
|---|---|---|---|
| Model: ang%Gird = −0.6288 + 0.01493(BBM) + 0.01034(Grass) + 0.06646(Tdiff) | −55.14 | −49.64 | 0.834 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, A.H.; Ratkowsky, D.A.; Wardlaw, T.J.; Mohammed, C.L. Ease of Access to An Alternative Food Source Enables Wallabies to Strip Bark in Tasmanian Pinus radiata Plantations. Forests 2020, 11, 387. https://doi.org/10.3390/f11040387
Smith AH, Ratkowsky DA, Wardlaw TJ, Mohammed CL. Ease of Access to An Alternative Food Source Enables Wallabies to Strip Bark in Tasmanian Pinus radiata Plantations. Forests. 2020; 11(4):387. https://doi.org/10.3390/f11040387
Chicago/Turabian StyleSmith, Anna H., David A. Ratkowsky, Timothy J. Wardlaw, and Caroline L. Mohammed. 2020. "Ease of Access to An Alternative Food Source Enables Wallabies to Strip Bark in Tasmanian Pinus radiata Plantations" Forests 11, no. 4: 387. https://doi.org/10.3390/f11040387
APA StyleSmith, A. H., Ratkowsky, D. A., Wardlaw, T. J., & Mohammed, C. L. (2020). Ease of Access to An Alternative Food Source Enables Wallabies to Strip Bark in Tasmanian Pinus radiata Plantations. Forests, 11(4), 387. https://doi.org/10.3390/f11040387