Presence of Root Rot Reduces Stability of Norway Spruce (Picea abies): Results of Static Pulling Tests in Latvia

 , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Sample Trees
2.2. Pulling Tests
2.3. Soil and Root Measurements
2.4. Data Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nabuurs, G.J.; Lindner, M.; Verkerk, P.J.; Gunia, K.; Deda, P.; Michalak, R.; Grassi, G. First sign of carbon sink saturation in European forest biomass. Nat. Clim. Change 2013, 3, 792–796. [Google Scholar] [CrossRef]
- Gardiner, B.; Schuck, A.R.T.; Schelhaas, M.J.; Orazio, C.; Blennow, K.; Nicoll, B. Living with Storm Damage to Forests; European Forest Institute: Joensuu, Finland, 2013; pp. 1–132. [Google Scholar]
- Seidl, R.; Rammer, W. Climate change amplifies the interactions between wind and bark beetle disturbances in forest landscapes. Landsc. Ecol. 2017, 32, 1485–1498. [Google Scholar] [CrossRef] [PubMed]
- Schelhaas, M.J.; Nabuurs, G.J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Change Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Laapas, M.; Lehtonen, I.; Venäläinen, A.; Peltola, H.M. The 10-Year Return Levels of Maximum Wind Speeds under Frozen and Unfrozen Soil Forest Conditions in Finland. Climate 2019, 7, 62. [Google Scholar] [CrossRef]
- Suvanto, S.; Henttonen, H.M.; Nöjd, P.; Mäkinen, H. Forest susceptibility to storm damage is affected by similar factors regardless of storm type: Comparison of thunder storms and autumn extra-tropical cyclones in Finland. For. Ecol. Manag. 2016, 381, 17–28. [Google Scholar] [CrossRef]
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schelhaas, M.J.; Lasch, P.; Eggers, J.; van der Maaten-Theunissen, M.; et al. Climate change and European forests: What do we know, what are the uncertainties, and what are the implications for forest management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef]
- Suvanto, S.; Peltoniemi, M.; Tuominen, S.; Strandström, M.; Lehtonen, A. High-resolution mapping of forest vulnerability to wind for disturbance-aware forestry. For. Ecol. Manag. 2019, 453, 117619. [Google Scholar] [CrossRef]
- Díaz-Yáñez, O.; Mola-Yudego, B.; González-Olabarria, J.R.; Pukkala, T. How does forest composition and structure affect the stability against wind and snow? For. Ecol. Manag. 2017, 401, 215–222. [Google Scholar] [CrossRef]
- Honkaniemi, J.; Lehtonen, M.; Väisänen, H.; Peltola, H. Effects of wood decay by Heterobasidion annosum on the vulnerability of Norway spruce stands to wind damage: A mechanistic modelling approach. Can. J. For. Res. 2017, 47, 777–787. [Google Scholar] [CrossRef]
- Gardiner, B.A.; Stacey, G.R.; Belcher, R.E.; Wood, C.J. Field and wind tunnel assessments of the implications of respacing and thinning for tree stability. Forestry 1997, 70, 233–252. [Google Scholar] [CrossRef]
- Hale, S.E.; Gardiner, B.A.; Wellpott, A.; Nicoll, B.C.; Achim, A. Wind loading of trees: Influence of tree size and competition. Eur. J. For. Res. 2012, 131, 203–217. [Google Scholar] [CrossRef]
- Stenlid, J.; Redfern, D.B. Spread within the Tree and Stand. In Heterobasidion Annosum. Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttemann, A., Eds.; CAB International: Wallingford, CT, USA, 1998; pp. 125–143. [Google Scholar]
- Zeng, H.; Pukkala, T.; Peltola, H. The use of heuristic optimisation in risk management of wind damage in forest planning. For. Ecol. Manag. 2007, 241, 189–199. [Google Scholar] [CrossRef]
- Heinonen, T.; Pukkala, T.; Ikonen, V.P.; Peltola, H.; Venäläinen, A.; Dupont, S. Integrating the risk of wind damage into forest planning. For. Ecol. Manag. 2009, 258, 1567–1577. [Google Scholar] [CrossRef]
- Heinonen, T.; Pukkala, T.; Ikonen, V.P.; Peltola, H.; Gregow, H.; Venäläinen, A. Consideration of strong winds, their directional distribution and snow loading in wind risk assessment related to landscape level forest planning. For. Ecol. Manag. 2011, 261, 710–719. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 2014, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Päivänen, J.; Hånell, B. Peatland Ecology and Forestry—A Sound Approach; Department of Forest Ecology, University of Helsinki: Helsinki, Finland, 2012; pp. 1–267. [Google Scholar]
- Albrecht, A.; Hanewinkel, M.; Bauhus, J.; Kohnle, U. How does silviculture affect storm damage in forests of south-western Germany? Results from empirical modeling based on long-term observations. Eur. J. For. Res. 2012, 131, 229–247. [Google Scholar] [CrossRef]
- Richter, C. Wood Characteristics: Description, Causes, Prevention, Impact on Use and Technological Adaptation. Springer Internationale Publishing: Basel, Switzerland, 2015; p. 222. [Google Scholar]
- Lodin, I. Choice of Tree Species in the Aftermath of Two Major Storms—A Qualitative Study of Private Forest Owners in Southern Sweden. Master’s Thesis, Swedish University of Agricultural Sciences, Alnarp, Sweden, January 2016. [Google Scholar]
- Woodward, S.; Stenlid, J.; Karjalainen, R.; Hüttemann, A. Heterobasidion Annosum. Biology, Ecology, Impact and Control; CAB International: Wallingford, CT, USA, 1998; p. 589. [Google Scholar]
- Nicoll, B.C.; Gardiner, B.A.; Rayner, B.; Peace, A.J. Anchorage of coniferous trees in relation to species, soil type, and rooting depth. Can. J. For. Res. 2006, 36, 1871–1883. [Google Scholar] [CrossRef]
- Lundström, T.; Jonas, T.; Stöckli, S.; Ammann, W. Anchorage of mature conifers: Resistive turning moment, root–soil plate geometry and root growth orientation. Tree Physiol. 2007, 27, 1217–1227. [Google Scholar]
- Bergeron, C.; Ruel, J.C.; Èlie, J.G.; Mitchell, S.J. Root anchorage and stem strength of black spruce (Picea mariana) trees in regular and irregular stands. Forestry 2009, 82, 29–41. [Google Scholar] [CrossRef]
- Giordano, L.; Lione, G.; Nicolotti, G.; Gonthier, P. Effect of Heterobasidion annosum s.l. root and butt rots on the stability of Norway spruce: An. uprooting test. In Proceedings of the XIII International Conference on Root and Butt Root of Forest Trees, Firenze (FI), S. Martino di Castrozza (TN), Italy, 4–10 September 2012; Capretti, P., Comparini, P., Garbelotto, M., La Porta, N., Santini, A., Eds.; University Press: Firenze, Italy, 2012; pp. 247–250. [Google Scholar]
- Göcke, L.; Rust, S.; Ruhl, F. Assessing the Anchorage and Critical Wind Speed of Urban Trees using Root Plate Inclination in high Winds. Arboric. Urban. For. 2018, 44, 1–11. [Google Scholar]
- Detter, A.; van Wassenaer, P.; Rust, S. Stability recovery in London Plane trees 8 years after primary anchorage failure. Arboric. Urban. For. 2019, 45, 279–288. [Google Scholar]
- Detter, A.; Richter, K.; Rust, C.; Rust, S. Aktuelle Untersuchungen zum Primärversagen von grünem Holz-Current studies on primary failure in green wood. In Proceedings of the Conference Deutsche Baumpflegetage, Augsburg, Germany, 5–7 May 2015; pp. 156–167. [Google Scholar]
- Detter, A.; Rust, S.; Rust, C.; Maybaum, G. Determining strength limits for standing tree stems from bending tests. In Proceedings of the 18th International Nondestructive Testing and Evaluation of Wood Symposium, Madison, WI, USA, 4–27 September 2013; Ross, R.J., Wang, X., Eds.; U.S. Department of Agriculture, Forest Service, Forest Products Laboratory: Madison, WI, USA, 2013; p. 226. [Google Scholar]
- Rogers, M.; Casey, A.; McMenamin, C. An experimental investigation of the effects of dynamic loading on coniferous trees planted on wet mineral soils. In Wind and Trees; Coutts, M.P., Grace, J., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 204–219. [Google Scholar]
- Bakeman, R. Recommended effect size statistics for repeated measures designs. Behav. Res. Methods 2005, 37, 379–384. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 5 December 2019).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Lawrence, M.A. EZ: Easy analysis and visualization of factorial experiments [Software]. 2011. (R package version 3.0-0). Available online: https://CRAN.R-project.org/package=ez (accessed on 2 November 2016).
- Liepa, I. Pieauguma mācība; [Increment theory]; Latvia University of Agriculture: Jelgava, Latvia, 1996; p. 123. [Google Scholar]
- Peltola, H.; Kellomäki, S.; Hassinen, A.; Granander, M. Mechanical stability of Scots pine, Norway spruce and birch: An analysis of tree-pulling experiments in Finland. For. Ecol. Manag. 2000, 135, 143–153. [Google Scholar] [CrossRef]
- Orth, R.; Seneviratne, S.I. Analysis of soil moisture memory from observations in Europe. J. Geophys. Res. 2012, 117, D15115. [Google Scholar] [CrossRef]
- Ray, D.; Nicoll, B.C. The effect of soil water-table depth on root-plate development and stability of Sitka spruce. Forestry 1998, 71, 169–182. [Google Scholar] [CrossRef]
- Kamimura, K.; Kitagawa, K.; Saito, S.; Mizunaga, H. Root anchorage of hinoki (Chamaecyparis obtuse (Sieb. Et Zucc.) Endl.) under the combined loading of wind and rapidly supplied water on soil: Analyses based on tree-pulling experiments. Eur J. Forest Res. 2012, 131, 219–227. [Google Scholar] [CrossRef]
- Detter, A.; Rust, S.; Böttcher, J.; Bouillon, J. Ambient influences on the results of non-destructive pulling tests. In Proceedings of the 21st International Nondestructive Testing and Evaluation of Wood Symposium, Freiburg, Germany, 24–27 September 2019; Forest Products Laboratory, General Technical Report: Madison, WI, USA, 2019. [Google Scholar]
- Pollet, C.; Henin, J.M.; Hébert, J.; Jourez, B. Effect of growth rate on the physical and mechanical properties of Douglas-fir in western Europe. Can. J. For. Res. 2017, 47, 1056–1065. [Google Scholar] [CrossRef]
- Mayr, S.; Bertel, C.; Dämon, B.; Beikircher, B. Static and dynamic bending has minor effects on xylem hydraulics of conifer branches (Picea abies, Pinus sylvestris). Plant. Cell Environ. 2014, 37, 2151–2157. [Google Scholar] [CrossRef]
- Seidl, R.; Blennow, K. Pervasive growth reduction in Norway Spruce forests following wind disturbance. PLoS ONE 2012, 7, e33301. [Google Scholar] [CrossRef]
- Honkaniemi, J.; Ojansuu, R.; Kasanen, R.; Heliövaara, K. Interaction of disturbance agents on Norway spruce: A mechanistic model of bark beetle dynamics integrated in simulation framework WINDROT. Ecol. Model. 2018, 388, 45–60. [Google Scholar] [CrossRef]
- Krisans, O.; Saleniece, R.; Rust, S.; Elferts, D.; Kapostins, R.; Jansons, A.; Matisons, R. Effect of Bark-Stripping on Mechanical Stability of Norway Spruce. Forests 2020, 11, 357. [Google Scholar] [CrossRef]


{kind=link}
| Stand | N | Soil Type | Tree Species (%) | Min DBH (cm) | Max DBH (cm) | Mean DBH (cm) |
|---|---|---|---|---|---|---|
| 1 | 20 | Fine sand | Norway spruce (80), birch (10), Scots pine (10) | 18.3 | 40 | 29 |
| 2 | 17 | Fine sand | Norway spruce (100) | 27 | 43.6 | 32.9 |
| 3 | 20 | Peat | Norway spruce (70), birch (20), black alder (10) | 21.7 | 46 | 32.2 |
| 4 | 20 | Peat | Norway spruce (100) | 23.7 | 44.2 | 34.7 |
| Variable | Root Rot | Control | ||||||
|---|---|---|---|---|---|---|---|---|
| Soil | Failure | Soil | Failure | |||||
| Peat | Fine Sand | Uproot | Fracture | Peat | Fine Sand | Uproot | Fracture | |
| DBH (cm) | 33 ± 6.3 | 29.2 ± 7 | 31.6 ± 6.5 | 28.4 ± 9 | 32 ± 4.8 | 30.5 ± 7.2 | 32.1 ± 5.4 | 21.5 ± 3.5 |
| Vsrp (m3) | 3.92 ± 2.04 | 3.1 ± 2.02 | 3.51 ± 2.01 | - | 4.12 ± 1.04 | 3.9 ± 1.59 | 4.03 ± 1.25 | - |
| SM (%) | 64.6 ± 20 | 39.5 ± 21.3 | 55.9 ± 23.3 | 31.7 ± 17.8 | 67.5 ± 17.4 | 38.2 ± 21.9 | 55.4 ± 23.1 | 37 ± 38.2 |
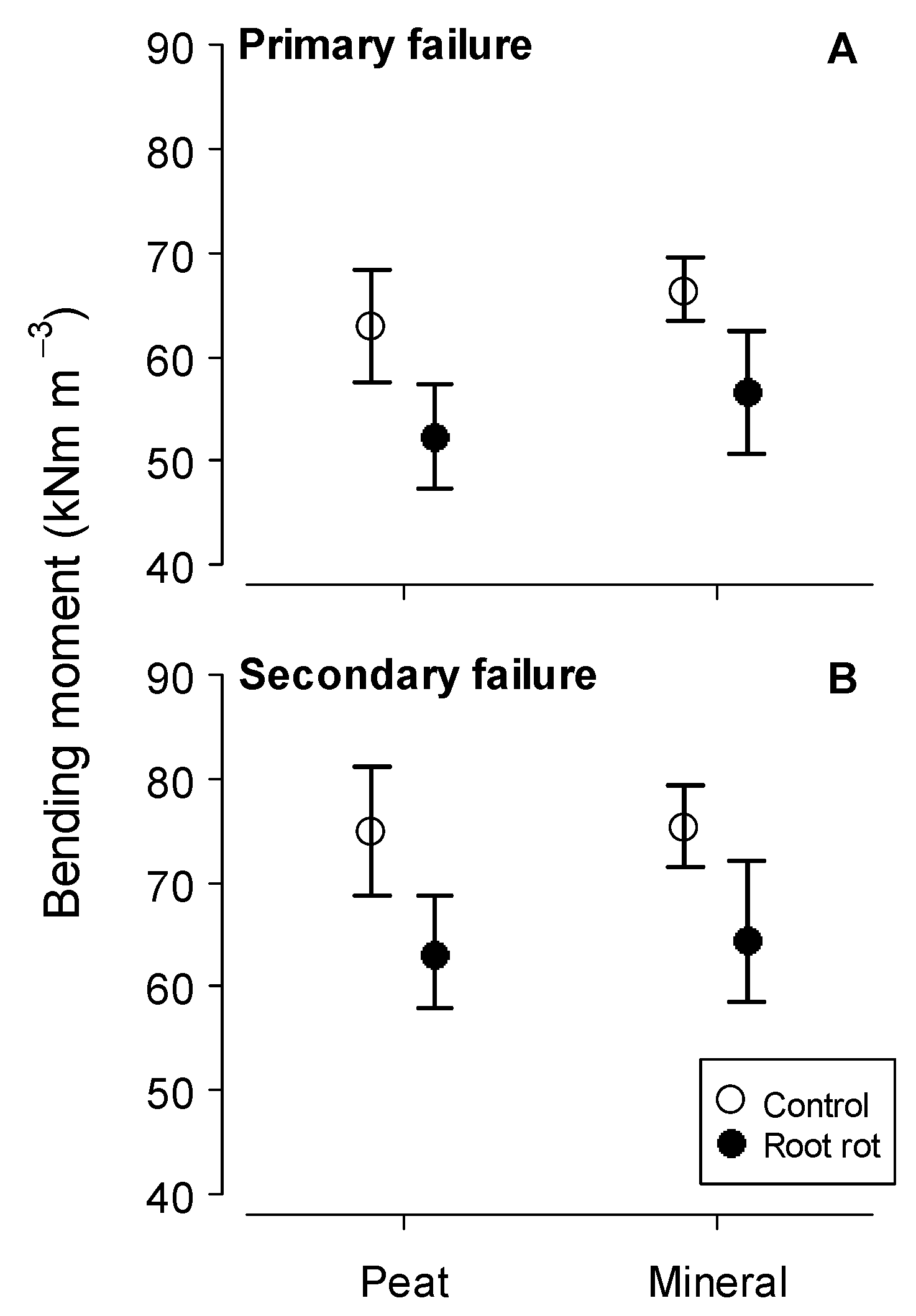
| BBMprim (kNm) | 56.8 ± 25.2 | 46.9 ± 26.7 | 53.5 ± 27.1 | 42.9 ± 18.9 | 60.1 ± 21.4 | 63.3 ± 26.9 | 64.2 ± 22.8 | 29.55 ± 5.8 |
| BBMsec (kNm) | 67.9 ± 29.6 | 53.2 ± 30 | 62.4 ± 31.4 | 50.8 ± 23.8 | 72 ± 25.9 | 71.3 ± 29.1 | 74.4 ± 26.1 | 38.7 ± 11.5 |
| MOE (GPa) | 38.4 ± 19.9 | 30 ± 15.4 | 34.8 ± 18.1 | 32.8 ± 20.8 | 28.2 ± 9 | 29.1 ± 9.6 | 28.1 ± 9.3 | 34.2 ± 2.7 |
| BBMprim (kNm) | F-Value | Ges * | p-Value |
|---|---|---|---|
| Soil | 0.15 | 0.07 | 0.73 |
| Root rot | 224.83 | 0.34 | <0.01 |
| Soil: root rot | 0.36 | <0.001 | 0.60 |
| BBMsec (kNm) | F-Value | Ges * | p-Value |
| Soil | 0.01 | 0.004 | 0.92 |
| Root rot | 39.69 | 0.44 | 0.02 |
| Soil: root rot | 0.05 | 0.001 | 0.83 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krisans, O.; Matisons, R.; Rust, S.; Burnevica, N.; Bruna, L.; Elferts, D.; Kalvane, L.; Jansons, A. Presence of Root Rot Reduces Stability of Norway Spruce (Picea abies): Results of Static Pulling Tests in Latvia. Forests 2020, 11, 416. https://doi.org/10.3390/f11040416
Krisans O, Matisons R, Rust S, Burnevica N, Bruna L, Elferts D, Kalvane L, Jansons A. Presence of Root Rot Reduces Stability of Norway Spruce (Picea abies): Results of Static Pulling Tests in Latvia. Forests. 2020; 11(4):416. https://doi.org/10.3390/f11040416
Chicago/Turabian StyleKrisans, Oskars, Roberts Matisons, Steffen Rust, Natalija Burnevica, Lauma Bruna, Didzis Elferts, Liene Kalvane, and Aris Jansons. 2020. "Presence of Root Rot Reduces Stability of Norway Spruce (Picea abies): Results of Static Pulling Tests in Latvia" Forests 11, no. 4: 416. https://doi.org/10.3390/f11040416
APA StyleKrisans, O., Matisons, R., Rust, S., Burnevica, N., Bruna, L., Elferts, D., Kalvane, L., & Jansons, A. (2020). Presence of Root Rot Reduces Stability of Norway Spruce (Picea abies): Results of Static Pulling Tests in Latvia. Forests, 11(4), 416. https://doi.org/10.3390/f11040416