Effects of Livestock Pressure and Vegetation Cover on the Spatial and Temporal Structure of Soil Microarthropod Communities in Iberian Rangelands

 ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Study Area
3. Materials and Methods
3.1. Determination of the Intensity of Livestock Pressure
3.2. Soil Sampling
3.3. Analysis of the Microarthropod Communities
3.4. Statistical Analyses
4. Results
4.1. Soil Parameters
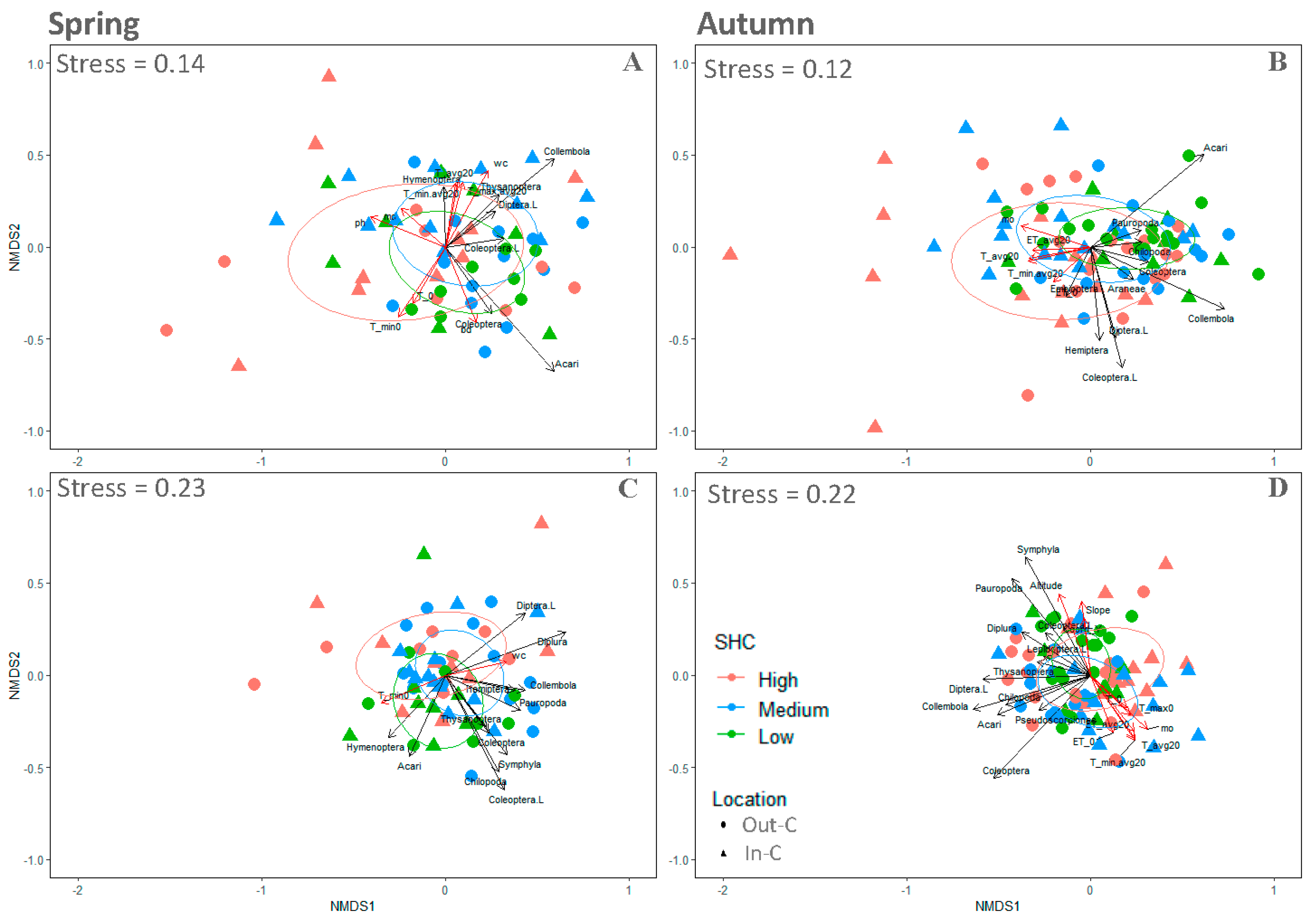
4.2. Seasonallity, Environmental Characteristics, and Community Structure
4.3. Spatial and Temporal Patterns of Abundances and QBS-ar
5. Discussion
5.1. The Response of Community Abundances
5.2. The Response of Biological Forms’ Evolutive Adaptation
5.3. Object-Based Image Analysis and SHC Classification
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Meteorological Variable | Units | Value | ||
|---|---|---|---|---|
| Annual solar radiation | W/m2 | 16.59 | ± | 6.76 |
| Net solar radiation | W/m2 | 7.52 | ± | 4.22 |
| Mean annual temperature | °C | 15.02 | ± | 6.30 |
| Maximum mean temperature of the coldest month | °C | 15.84 | ± | 0.62 |
| Minimum mean temperature of the coldest month | °C | 2.41 | ± | 0.27 |
| Maximum mean temperature of the warmest month | °C | 30.60 | ± | 6.46 |
| Minimum mean temperature of the warmest month | °C | 13.61 | ± | 4.22 |
| Mean annual rainfall | mm | 524.2 | ± | 28.4 |
| Mean annual effective precipitation | mm | 249.8 | ± | 14.4 |
| Mean annual evapotranspiration | mm | 1363.1 | ± | 75.5 |
| Taxa | EMI Score |
| Pseudoscorpiones | 20 |
| Opiliones | 10 |
| Araneae | 1–5 |
| Acari | 20 |
| Isopoda | 10 |
| Diplopoda | 10–20 |
| Pauropoda | 20 |
| Symphyla | 20 |
| Chilopoda | 10–20 |
| Protura | 20 |
| Diplura | 20 |
| Collembola | 1–20 |
| Psocoptera | 1 |
| Hemiptera | 1 |
| Thysanoptera | 1 |
| Zigentomi | 10 |
| Embioptera | 10 |
| Orthroptera | 1–20 |
| Coleoptera | 1–20 |
| Hymenoptera | 1–5 |
| Diptera | 1 |
| Lepidoptera | 1 |
| Coleoptera (larvae) | 10 |
| Diptera (larvae) | 10 |
| Hymenoptera (larvae) | 10 |
| Lepidoptera (larvae) | 10 |
| Holometabolans (adults) | 1 |
| Metrics | Factors | Pairs Comparison | Difference | p |
|---|---|---|---|---|
| Ln abundances | Livestock pressure | Low − High | 0.568 | 0.045 |
| Location | In-C − Out-C | –0.07 | <0.001 | |
| Season | Spring − Autumn | –0.051 | 0.007 | |
| Livestock pressure × Location | High × In-C − Low × Out-C | –1.108 | 0.009 | |
| High × In-C − Medium × Out-C | –1.147 | 0.004 | ||
| Medium × In-C − Medium × Out-C | –0.9 | 0.03 | ||
| Livestock pressure × Season | Low × Autumn − High × Autumn | 0.882 | 0.038 | |
| High × Spring − Low × Autumn | –1.093 | 0.032 | ||
| Medium × Spring − Low × Autumn | –0.924 | 0.036 | ||
| Location × Season | In-C × Autumn − Out-C × Autumn | –0.651 | 0.036 | |
| In-C × Spring − Out-C × Autumn | –1.199 | <0.001 | ||
| Livestock pressure × Location × Season | High × In-C × Spring − Low × Out-C × Autumn | –1.526 | 0.048 | |
| High × In-C × Autumn − Medium × Out-C × Autumn | –1.427 | 0.043 | ||
| High × In-C × Spring − Medium × Out-C × Autumn | –1.733 | 0.019 | ||
| High × In-C × Spring − Low × In-C × Autumn | –1.781 | 0.035 | ||
| QBS-ar | Location | In-C − Out-C | –19.693 | <0.001 |
| Livestock pressure × Location | High × In-C − High × Out-C | –24.222 | 0.037 | |
| High × In-C − Low × Out-C | –28.101 | 0.009 | ||
| High × In-C − Medium × Out-C | –33.791 | <0.001 | ||
| Medium × In-C − Medium × Out-C | –23.922 | 0.027 | ||
| Livestock pressure × Season | High × Spring − Low × Autumn | –23.295 | 0.09 | |
| Location × Season | In-C × Autumn − Out-C × Autumn | –27.661 | <0.001 | |
| In-C × Spring − Out-C × Autumn | –24.243 | 0.002 | ||
| Livestock pressure × Location × Season | High × In-C × Autumn − High × Out-C × Autumn | –39.419 | 0.08 | |
| High × In-C × Autumn − Low × Out-C × Autumn | –34.004 | 0.051 | ||
| H′ | Location × Season | In-C × Spring − Out-C × Spring | 0.197 | 0.063 |
| Spring | Autumn | |||
|---|---|---|---|---|
| Livestock Pressure | Out-C | In-C | Out-C | In-C |
| SHChigh | 66.9 ± 28.3 | 72.1 ± 27.2 | 104.4 ± 34.2 | 64.9 ± 23.2 |
| SHCmedium | 97.1 ± 39.4 | 80.3 ± 22.2 | 105.9 ± 27.8 | 76.8 ± 32.3 |
| SHClow | 91.3 ± 22.4 | 84.8 ± 22.7 | 98.9 ± 18.4 | 89.0 ± 31.5 |
| Spring | Autumn | |||
|---|---|---|---|---|
| Livestock Pressure | Out-C | In-C | Out-C | In-C |
| SHChigh | 0.79 ± 0.36 | 0.78 ± 0.33 | 0.97 ± 0.27 | 0.89 ± 0.23 |
| SHCmedium | 0.76 ± 0.20 | 1.04 ± 0.41 | 0.86 ± 0.34 | 0.83 ± 0.29 |
| SHClow | 0.69 ± 0.29 | 1.00 ± 0.48 | 0.76 ± 0.26 | 0.68 ± 0.24 |
| Spring | ||||||
|---|---|---|---|---|---|---|
| Out-C | In-C | |||||
| Taxa | SHChigh | SHCmedium | SHClow | SHChigh | SHCmedium | SHClow |
| Pseudoscorpiones | - | 3 | - | - | - | 3 |
| Opiliones | - | - | - | - | - | - |
| Araneae | 1 | - | 3 | - | 2 | 2 |
| Acari | 3410 | 3928 | 2882 | 902 | 1116 | 2030 |
| Isopoda | - | - | - | - | - | - |
| Diplopoda | - | - | - | - | - | 2 |
| Pauropoda | 1 | 8 | 1 | 3 | - | - |
| Symphyla | - | 10 | 3 | - | 3 | - |
| Chilopoda | 2 | 17 | 18 | 4 | 19 | 8 |
| Protura | - | 1 | - | - | - | 3 |
| Diplura | 30 | 29 | 24 | 3 | 7 | - |
| Collembola | 995 | 4368 | 1046 | 3424 | 5584 | 936 |
| Psocoptera | - | - | - | 5 | 2 | 7 |
| Hemiptera | 1 | 51 | 30 | 14 | 34 | 13 |
| Thysanoptera | 1 | 24 | 7 | 6 | 20 | 8 |
| Zigentomi | - | - | - | - | - | - |
| Embioptera | 1 | 2 | - | 6 | 4 | 2 |
| Orthroptera | - | - | - | 1 | - | - |
| Coleoptera | 46 | 180 | 28 | 25 | 18 | 12 |
| Hymenoptera | 5 | 26 | 64 | 17 | 153 | 31 |
| Diptera | - | 1 | 2 | 2 | 7 | 6 |
| Lepidoptera | - | - | 1 | 1 | 1 | - |
| Coleoptera (larvae) | 36 | 31 | 30 | 14 | 31 | 34 |
| Diptera (larvae) | 29 | 218 | 63 | 64 | 120 | 135 |
| Hymenoptera (larvae) | - | - | - | 4 | - | - |
| Lepidoptera (larvae) | 1 | 3 | 1 | - | 2 | 2 |
| Holometabolans | - | - | - | - | - | - |
| Total | 4559 | 8900 | 4203 | 4495 | 7123 | 3234 |
| Autumn | ||||||
|---|---|---|---|---|---|---|
| Out-C | In-C | |||||
| Taxa | SHChigh | SHCmedium | SHClow | SHChigh | SHCmedium | SHClow |
| Pseudoscorpiones | 1 | 6 | 1 | - | 1 | - |
| Opiliones | - | - | - | - | - | - |
| Araneae | 1 | 5 | 12 | 4 | 3 | 1 |
| Acari | 4003 | 5375 | 10370 | 1321 | 2599 | 2498 |
| Isopoda | - | - | - | - | 1 | - |
| Diplopoda | - | - | - | - | - | - |
| Pauropoda | 16 | 383 | 56 | 1 | 29 | 97 |
| Symphyla | 18 | 9 | 20 | 7 | 1 | 2 |
| Chilopoda | 5 | 11 | 2 | - | 11 | 1 |
| Protura | 2 | 2 | 7 | - | 1 | 1 |
| Diplura | 8 | 1 | - | - | 3 | 1 |
| Collembola | 7079 | 9787 | 13472 | 4708 | 4892 | 10,574 |
| Psocoptera | - | - | - | - | - | 2 |
| Hemiptera | 102 | 22 | 18 | 243 | 12 | 17 |
| Thysanoptera | 34 | 29 | 11 | 7 | 36 | 13 |
| Zigentomi | - | - | 3 | 1 | - | - |
| Embioptera | - | - | - | 7 | 2 | 5 |
| Orthroptera | - | - | - | - | - | - |
| Coleoptera | 96 | 85 | 29 | 10 | 36 | 32 |
| Hymenoptera | 3 | 17 | 17 | 80 | 58 | 10 |
| Diptera | 31 | 15 | 16 | 7 | 25 | 13 |
| Lepidoptera | - | - | - | - | - | - |
| Coleoptera (larvae) | 152 | 101 | 92 | 137 | 52 | 69 |
| Diptera (larvae) | 1374 | 712 | 226 | 238 | 84 | 71 |
| Hymenoptera (larvae) | - | - | - | - | - | - |
| Lepidoptera (larvae) | 70 | 16 | 21 | 21 | 4 | 2 |
| Holometabolans | 3 | 6 | 1 | 1 | 2 | 1 |
| Total | 12,424 | 16,382 | 24,310 | 6700 | 7839 | 13,410 |
References
- Jeffery, S.; Gardi, C.; Jones, A.; Montanarella, L.; Marmo, L.; Miko, L.; Ritz, K.; Peres, G.; Römbke, J.; Van der Putten, W.H. European Atlas of Soil Biodiversity; Publications Office of the European Union: Luxembourg, 2010. [Google Scholar]
- Adhikari, K.; Hartemink, A.E. Linking soils to ecosystem services—A global review. Geoderma 2016, 262, 101–111. [Google Scholar] [CrossRef]
- Lal, R. Soil carbon sequestration to mitigate climate change. Geoderma 2004, 123, 1–22. [Google Scholar] [CrossRef]
- Wagg, C.; Bender, S.F.; Widmer, F.; van der Heijden, M.G.A. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, D.C.; Wall, D.H. Chapter 5: Soil Fauna: Occurrence, Biodiversity, and Roles in Ecosystem Function. In Soil Microbiology Ecology and Biochemistry; Eldor, P.A., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 111–149. ISBN 9780124159556. [Google Scholar]
- Bardgett, R.D. Causes and consequences of biological diversity in soil. Zoology 2006, 105, 367–374. [Google Scholar] [CrossRef] [PubMed]
- IPCC. 2014 Climate Change 2014: Mitigation of Climate Change. Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Edenhofer, O., Pichs-Madruga, R., Sokona, Y., Farahani, E., Kadner, S., Seyboth, K., Adler, A., Eds.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Smith, P.; Nkem, J.; Calvin, K.; Campbell, D.; Cherubini, F.; Grassi, G.; Korotkov, V.; Le Hoang, A.; Lwasa, S.; McElwee, P.; et al. Interlinkages between Desertification, Land Degradation, Food Security and GHG fluxes: Synergies, trade-offs and Integrated Response Options. In Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Shukla, P.R., Skea, J., Calvo Buendía, E., Masson-Delmotte, V., Pörtner, H.O., Roberts, D.C., Zhai, P., Slade, R., Connors, S., van Diemen, R., Eds.; IPCC: Geneva, Switzerland, 2019; in press. [Google Scholar]
- Mulder, C.; Boit, A.; Bonkowski, M.; De Ruiter, P.C.; Mancinelli, G.; Van der Heijden, M.G.A.; Van Wijnen, H.J.; Vonk, J.A.; Rutgers, M. A Belowground Perspective on Dutch Agroecosystems: How Soil Organisms Interact to Support Ecosystem Services. In Advances in Ecological Research; Elsevier Ltd.: Amsterdam, The Netherlands, 2011; Volume 44, pp. 277–357. ISBN 9780123747945. [Google Scholar]
- IPCC. Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Shukla, P.R., Skea, J., Calvo Buendía, E., Masson-Delmotte, V., Pörtner, H.O., Roberts, D.C., Zhai, P., Slade, R., Connors, S., van Diemen, R., Eds.; IPCC: Geneva, Switzerland, 2019; in press. [Google Scholar]
- Parisi, V.; Menta, C.; Gardi, C.; Jacomini, C.; Mozzanica, E. Microarthropod communities as a tool to assess soil quality and biodiversity: A new approach in Italy. Agric. Ecosyst. Environ. 2005, 105, 323–333. [Google Scholar] [CrossRef]
- Menta, C.; Conti, F.D.; Pinto, S.; Bodini, A. Soil Biological Quality index (QBS-ar): 15 years of application at global scale. Ecol. Indic. 2018, 85, 773–780. [Google Scholar] [CrossRef]
- Menta, C.; Conti, F.D.; Pinto, S.; Leoni, A.; Lozano-Fondón, C. Monitoring soil restoration in an open-pit mine in northern Italy. Appl. Soil Ecol. 2014, 83, 22–29. [Google Scholar] [CrossRef]
- Soong, J.L.; Nielsen, U.N. The role of microarthropods in emerging models of soil organic matter. Soil Biol. Biochem. 2016, 102, 37–39. [Google Scholar] [CrossRef] [Green Version]
- Andrés, P.; Moore, J.C.; Simpson, R.T.; Selby, G.; Cotrufo, F.; Denef, K.; Haddix, M.L.; Shaw, E.A.; de Tomasel, C.M.; Molowny-Horas, R.; et al. Soil food web stability in response to grazing in a semi-arid prairie: The importance of soil textural heterogeneity. Soil Biol. Biochem. 2016, 97, 131–143. [Google Scholar] [CrossRef]
- Van Dam, N.M.; Heil, M. Multitrophic interactions below and above ground: En route to the next level. J. Ecol. 2011, 99, 77–88. [Google Scholar] [CrossRef]
- Menta, C.; Leoni, A.; Gardi, C.; Delia Conti, F. Are grasslands important habitats for soil microarthropod conservation? Biodivers. Conserv. 2011, 20, 1073–1087. [Google Scholar] [CrossRef]
- Wardle, D.A.; Yeates, G.W.; Williamson, W.; Bonner, K.I. The response of a three trophic level soil food web to the identity and diversity of plant species and functional groups. Oikos 2003, 102, 45–56. [Google Scholar] [CrossRef]
- Lindo, Z.; Whiteley, J.; Gonzalez, A. Traits explain community disassembly and trophic contraction following experimental environmental change. Glob. Chang. Biol. 2012, 18, 2448–2457. [Google Scholar] [CrossRef]
- Lacetera, N.; Segnalini, M.; Bernabucci, U.; Ronchi, B.; Vitali, A.; Tran, A.; Guis, H.; Caminade, C.; Calvete, C.; Morse, A.; et al. Climate Induced Effects on Livestock Population and Productivity in the Mediterranean Area. In Regional Assessment of Climate Change in the Mediterranean. Volume 2: Agriculture, Forests and Ecosystem Services and People; Navarra, A., Tubiana, L., Eds.; Advances in Global Change Research; Springer: Dordrecht, The Netherlands, 2013; pp. 135–156. [Google Scholar]
- Moreno, G.; Pulido, F.J. The Functioning, Management and Persistence of Dehesas. In Agroforestry in Europe; Springer: Dordrecht, The Netherlands, 2009; Volume 10600, pp. 127–160. ISBN 9781402082719. [Google Scholar]
- Fernández, M.P.; Contador, J.F.L.; Schnabel, S.; Gutiérrez, Á.G.; Lozano-Parra, J. Changes in Land Management of Iberian Rangelands and Grasslands in the Last 60 Years and their Effect on Vegetation. In Vegetation; Sebata, A., Ed.; IntechOpen: London, UK, 2018. [Google Scholar]
- Lozano-Parra, J.; Maneta, M.P.; Schnabel, S. Climate and topographic controls on simulated pasture production in a semiarid Mediterranean watershed with scattered tree cover. Hydrol. Earth Syst. Sci. 2014, 18, 1439–1456. [Google Scholar] [CrossRef] [Green Version]
- Schanbel, S.; Dahlgren, R.A.; Moreno-Marcos, G. Soil and water dymamics. In Mediiterranean Oak Woodland Working Landscapes: Dehesas of Spain and Ranchlands of California; Campos, P., Starrs, P.F., Huntsinger, L., Montero, G., Díaz, M., Eds.; Springer-Verlag: New York, NY, USA, 2013; pp. 91–121. ISBN 978-94-007-6706-5. [Google Scholar]
- Pulido, M.; Schnabel, S.; Lavado-Contador, F.; Lozano-Parra, J.; Gómez-Gutiérrez, Á. Selecting indicators for assessing soil quality and degradation in rangelands of Extremadura (SW Spain). Ecol. Indic. 2017, 74, 49–61. [Google Scholar] [CrossRef]
- Lozano-Parra, J.; Schnabel, S.; Ceballos-Barbancho, A. The role of vegetation covers on soil wetting processes at rainfall event scale in scattered tree woodland of Mediterranean climate. J. Hydrol. 2015, 529, 951–961. [Google Scholar] [CrossRef]
- Moreno, G.; Gonzalez-Bornay, G.; Pulido, F.; Lopez-Diaz, M.L.; Bertomeu, M.; Juárez, E.; Diaz, M. Exploring the causes of high biodiversity of Iberian dehesas: The importance of wood pastures and marginal habitats. Agrofor. Syst. 2015, 90, 87–105. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; King, J.S.; Burton, A.J.; Brown, S.E. Responses of tree fine roots to temperature.pdf. New Phytol. 2000, 147, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Lozano-Parra, J.; Pulido, M.; Lozano-Fondón, C.; Schnabel, S. How do soil moisture and vegetation covers influence soil temperature in drylands of Mediterranean regions? Water (Switzerland) 2018, 10, 1747. [Google Scholar] [CrossRef] [Green Version]
- Pulido, M.; Schnabel, S.; Francisco, J.; Contador, L.; Lozano-parra, J.; González, F. The impact of heavy grazing on soil quality and pasture production in rangelands of SW Spain. Land Degrad. Dev. 2016, 29, 219–230. [Google Scholar] [CrossRef]
- Jiménez-Chacón, A.; Homet, P.; Matías, L.; Gómez-Aparicio, L.; Godoy, O. Fine Scale Determinants of Soil Litter Fauna on a Mediterranean Mixed Oak Forest Invaded by the Exotic Soil-Borne Pathogen Phytophthora cinnamomi. Forests 2018, 9, 218. [Google Scholar] [CrossRef] [Green Version]
- Fuls, E.R. A technique for objective habitat condition assessments in rangelands. J. Arid Environ. 1992, 22, 195–198. [Google Scholar] [CrossRef]
- FAO. FAO-UNESCO Soil Map of the World; FAO: Rome, Italy, 2006. [Google Scholar]
- Ministerio de Fomento Centro Nacional de Información Geográfica. Available online: https://www.cnig.es/home (accessed on 31 May 2020).
- Drǎguţ, L.; Tiede, D.; Levick, S.R. ESP: A tool to estimate scale parameter for multiresolution image segmentation of remotely sensed data. Int. J. Geogr. Inf. Sci. 2010, 24, 859–871. [Google Scholar] [CrossRef]
- Ma, L.; Li, M.; Ma, X.; Cheng, L.; Du, P.; Liu, Y. A review of supervised object-based land-cover image classification. ISPRS J. Photogramm. Remote Sens. 2017, 130, 277–293. [Google Scholar] [CrossRef]
- Blaschke, T.; Lang, S.; Lorup, E.; Strobl, J.; Zeil, P. Object-oriented image processing in an integrated GIS/remote sensing environment and perspectives for environmental applications. In Environmental Information for Planning. Politics and the Public, Vol. 2; Cremers, A., Greve, K., Eds.; dc.Publisher: Marburg, Germany, 2000; pp. 555–570. [Google Scholar]
- Myint, S.W.; Gober, P.; Brazel, A.; Grossman-Clarke, S.; Weng, Q. Per-pixel vs. object-based classification of urban land cover extraction using high spatial resolution imagery. Remote Sens. Environ. 2011, 115, 1145–1161. [Google Scholar] [CrossRef]
- Meloni, F.; Civieta, B.F.; Zaragoza, J.A.; Bautista, S. Vegetation Pattern Modulates Ground Arthropod Diversity in Semi-Arid Mediterranean Steppes. Insects 2020, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- LI-COR Inc. LI-8100A Automated Soil CO2 Flux System Instruction Manual; LI-COR Inc.: Lincoln, NE, USA, 2010. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Borcard, D.; Gillet, F.; Legrende, P. Numerical Ecology with R.; Springer Science & Business Media: New York, NY, USA, 2011; ISBN 978-0-387-78170-9. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- McArdle, B.; Anderson, M.J. Fitting Multivariate Models To Community Data . Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Oksanen, J.; Guillaume Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, H.H.; Wagner, H. Vegan: Community Ecology Package, R Package Version 1.15-1; R Foundation for Statistical Computing: Vienna, Austria, 2019.
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: New York, NY, USA, 2009; ISBN 9781351414234. [Google Scholar]
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. (B) 2011, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Gao, M.; Liu, D.; Lin, L.; Wu, D. The small-scale structure of a soil mite metacommunity. Eur. J. Soil Biol. 2016, 74, 69–75. [Google Scholar] [CrossRef]
- Tilman, D. Niche tradeoffs, neutrality, and community structure: A stochastic theory of resource competition, invasion, and community assembly. Proc. Natl. Acad. Sci. USA 2004, 101, 10854–10861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, M.; He, P.; Zhang, X.; Liu, D.; Wu, D. Relative roles of spatial factors, environmental filtering and biotic interactions in fine-scale structuring of a soil mite community. Soil Biol. Biochem. 2014, 79, 68–77. [Google Scholar] [CrossRef]
- Sha, D.; Gao, M.; Sun, X.; Wu, D.; Zhang, X. Relative Contributions of Spatial and Environmental Processes and Biotic Interactions in a Soil Collembolan Community. Chin. Geogr. Sci. 2015, 25, 582–590. [Google Scholar] [CrossRef]
- Gao, M.; Sun, X.; Qiao, Z.; Hou, H.; Lu, T.; Wu, D. Distinct patterns suggest that assembly processes di ff er for dominant arthropods in above-ground and below-ground ecosystems. Pedobiol. J. Soil Ecol. 2018, 69, 17–28. [Google Scholar] [CrossRef]
- Caruso, T.; Trokhymets, V.; Bargagli, R.; Convey, P. Biotic interactions as a structuring force in soil communities: Evidence from the micro-arthropods of an Antarctic moss model system. Oecologia 2013, 172, 495–503. [Google Scholar] [CrossRef]
- Caruso, T.; Taormina, M.; Migliorini, M. Relative role of deterministic and stochastic determinants of soil animal community: A spatially explicit analysis of oribatid mites. J. Anim. Ecol. 2012, 81, 214–221. [Google Scholar] [CrossRef]
- Dong, C.; Gao, M.; Guo, C.; Lin, L.; Wu, D. The underlying processes of a soil mite metacommunity on a small scale. PLoS ONE 2017, 12, e0176828. [Google Scholar] [CrossRef]
- Mulder, C.; Den Hollander, H.A.; Hendriks, A.J. Aboveground Herbivory Shapes the Biomass Distribution and Flux of Soil Invertebrates. PLoS ONE 2008, 3, e3573. [Google Scholar] [CrossRef]
- Parisi, V.; Menta, C. Microarthropods of the soil: Convergence phenomena and evaluation of soil quality using QBS-ar and QBS-C. Fresenius Environ. Bull. 2008, 17, 1170–1174. [Google Scholar]
- Schon, N.L.; Mackay, A.D.; Minor, M.A.; Yeates, G.W.; Hedley, M.J. Soil fauna in grazed New Zealand hill country pastures at two management intensities. Appl. Soil Ecol. 2008, 40, 218–228. [Google Scholar] [CrossRef]
- Peco, B.; Navarro, E.; Carmona, C.P.; Medina, N.G.; Marques, M.J. Effects of grazing abandonment on soil multifunctionality: The role of plant functional traits. Agric. Ecosyst. Environ. 2017, 249, 215–225. [Google Scholar] [CrossRef]
- Schnabel, S.; Gómez Gutiérrez, A.; Lavado Contador, J.F. Grazing and soil erosion in dehesas of SW Spain. In Advances in Studies on Desertification; International Nuclear Information System: Vienna, Austria, 2009; pp. 725–728, 732. ISBN 978-84-8371-888-9. [Google Scholar]
- Junta de Extremadura REDAREX: Red de Asesoramiento al Regante de Extremadura. Available online: http://redarexplus.gobex.es/RedarexPlus/ (accessed on 15 May 2019).





| SHChigh | SHCmedium | SHClow | |||||
|---|---|---|---|---|---|---|---|
| Units | Out-C | In-C | Out-C | In-C | Out-C | In-C | |
| Bulk density | g cm−1 | 1.5 ± 0.1 | 1.2 ± 0.2 | 1.5 ± 0.1 | 1.2 ± 0.2 | 1.5 ± 0.2 | 1.2 ± 0.1 |
| Organic matter | % | 5.3 ± 2.5 | 10.3 ± 4.6 | 1.9 ± 1.8 | 9.1 ± 4.6 | 3.8 ± 1.4 | 9.5 ± 4.1 |
| pH | - | 5.8 ± 0.7 | 6.0 ± 0.8 | 5.6 ± 0.4 | 5.9 ± 0.4 | 5.8 ± 0.2 | 5.7 ± 0.7 |
| Soil CO2 efflux | µmol m−2 s−1 | 4.6 ± 2.8 | 4.9 ± 2.7 | 4.5 ± 1.4 | 5.2 ± 1.9 | 5.2 ± 1.9 | 5.0 ± 2.2 |
| Water content | % | 18.4 ± 5.9 | 20.8 ± 8.3 | 22.2 ± 8.8 | 23.2 ± 8.0 | 21.8 ± 8.4 | 23.7 ± 10.5 |
| Ln Abundances | QBS-ar | H′ | ||||
|---|---|---|---|---|---|---|
| Suorce of Variation | F Test | p-Value | F Test | p-Value | F Test | p-Value |
| Livestock pressure | 2.911 | 0.058 | 2.451 | 0.090 | 1.532 | 0.220 |
| Location | 14.655 | <0.001 *** | 17.464 | <0.001 *** | 0.913 | 0.341 |
| Season | 7.644 | 0.007 ** | 2.057 | 0.154 | 0.007 | 0.932 |
| Livestock pressure × Location | 1.233 | 0.295 | 1.059 | 0.350 | 1.209 | 0.302 |
| Livestock pressure × Season | 1.355 | 0.262 | 0.511 | 0.601 | 2.247 | 0.110 |
| Location × Season | 0.015 | 0.902 | 4.490 | 0.036 * | 6.232 | 0.014 * |
| Livestock pressure × Location × Season | 1.718 | 0.184 | 1.581 | 0.210 | 0.634 | 0.532 |
| Community Matrix | Season | Source of Dissimilarity | Df | F | R2 |
|---|---|---|---|---|---|
| Log-transformed | Spring | Location | 1 | 2.674 | 0.041 * |
| Abundances | Water content | 1 | 3.444 | 0.052 ** | |
| pH | 1 | 2.480 | 0.037 * | ||
| Soil CO2 efflux | 1 | 2.263 | 0.034 * | ||
| Residuals | 54 | 0.836 | |||
| Log-transformed | Autumn | Location | 1 | 6.217 | 0.062 *** |
| Abundances | SHC | 2 | 2.184 | 0.044 * | |
| Slope | 1 | 3.173 | 0.032 * | ||
| OM content | 1 | 3.530 | 0.035 ** | ||
| T (−20) | 1 | 2.720 | 0.027 * | ||
| EP (−20) | 1 | 2.156 | 0.022 * | ||
| Residuals | 78 | 0.779 | |||
| EMIs | Spring | Location | 1 | 2.097 | 0.034 |
| Water content | 1 | 3.155 | 0.051 * | ||
| Residuals | 57 | 0.916 | |||
| EMIs | Autumn | Location | 1 | 11.329 | 0.111 *** |
| Slope | 1 | 5.055 | 0.050 ** | ||
| T (−20) | 1 | 3.456 | 0.036 ** | ||
| Residuals | 82 | 0.803 |
| Metrics | Season | Parameter | F | p | R2 |
|---|---|---|---|---|---|
| Log-transformed | Spring | s(NMDS1, NDMS2) | 21.750 | <0.001 | 0.874 |
| Abundances | Location-out-C | 7.920 | <0.001 | ||
| Location-In-C | −0.948 | 0.348 | |||
| SHClow | −1.705 | 0.095 | |||
| SHCmedium | −1.444 | 0.156 | |||
| pH | −1.416 | 0.164 | |||
| OM content | −1.529 | 0.134 | |||
| Bulk density | −1.536 | 0.132 | |||
| Autumn | s(NMDS1, NDMS2) | 23.880 | <0.001 | 0.891 | |
| Location-out-C | 8.923 | <0.001 | |||
| Location-In-C | 1.742 | 0.087 | |||
| SHClow | 1.899 | 0.062 | |||
| SHCmedium | 1.066 | 0.291 | |||
| T (−20) | −1.605 | 0.114 | |||
| EP (−20) | 2.539 | 0.014 | |||
| QBS-ar | Spring | s(NMDS1, NDMS2) | 12.870 | <0.001 | 0.855 |
| Location-out-C | 35.507 | <0.001 | |||
| Location-In-C | −0.873 | 0.389 | |||
| Autumn | s(NMDS1, NDMS2) | 37.430 | <0.001 | 0.918 | |
| Location-out-C | 15.892 | <0.001 | |||
| Location-In-C | 0.107 | 0.915 | |||
| OM content | −2.214 | 0.030 | |||
| EP (−20) | −1.888 | 0.063 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lozano Fondón, C.; González, J.B.; Pulido Fernández, M.; Remelli, S.; Lozano-Parra, J.; Menta, C. Effects of Livestock Pressure and Vegetation Cover on the Spatial and Temporal Structure of Soil Microarthropod Communities in Iberian Rangelands. Forests 2020, 11, 628. https://doi.org/10.3390/f11060628
Lozano Fondón C, González JB, Pulido Fernández M, Remelli S, Lozano-Parra J, Menta C. Effects of Livestock Pressure and Vegetation Cover on the Spatial and Temporal Structure of Soil Microarthropod Communities in Iberian Rangelands. Forests. 2020; 11(6):628. https://doi.org/10.3390/f11060628
Chicago/Turabian StyleLozano Fondón, Carlos, Jesús Barrena González, Manuel Pulido Fernández, Sara Remelli, Javier Lozano-Parra, and Cristina Menta. 2020. "Effects of Livestock Pressure and Vegetation Cover on the Spatial and Temporal Structure of Soil Microarthropod Communities in Iberian Rangelands" Forests 11, no. 6: 628. https://doi.org/10.3390/f11060628