Frugivory by Coyotes Decreases the Time to Germination and Increases the Growth of Netleaf Hackberry (Celtis reticulata) Seedlings



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
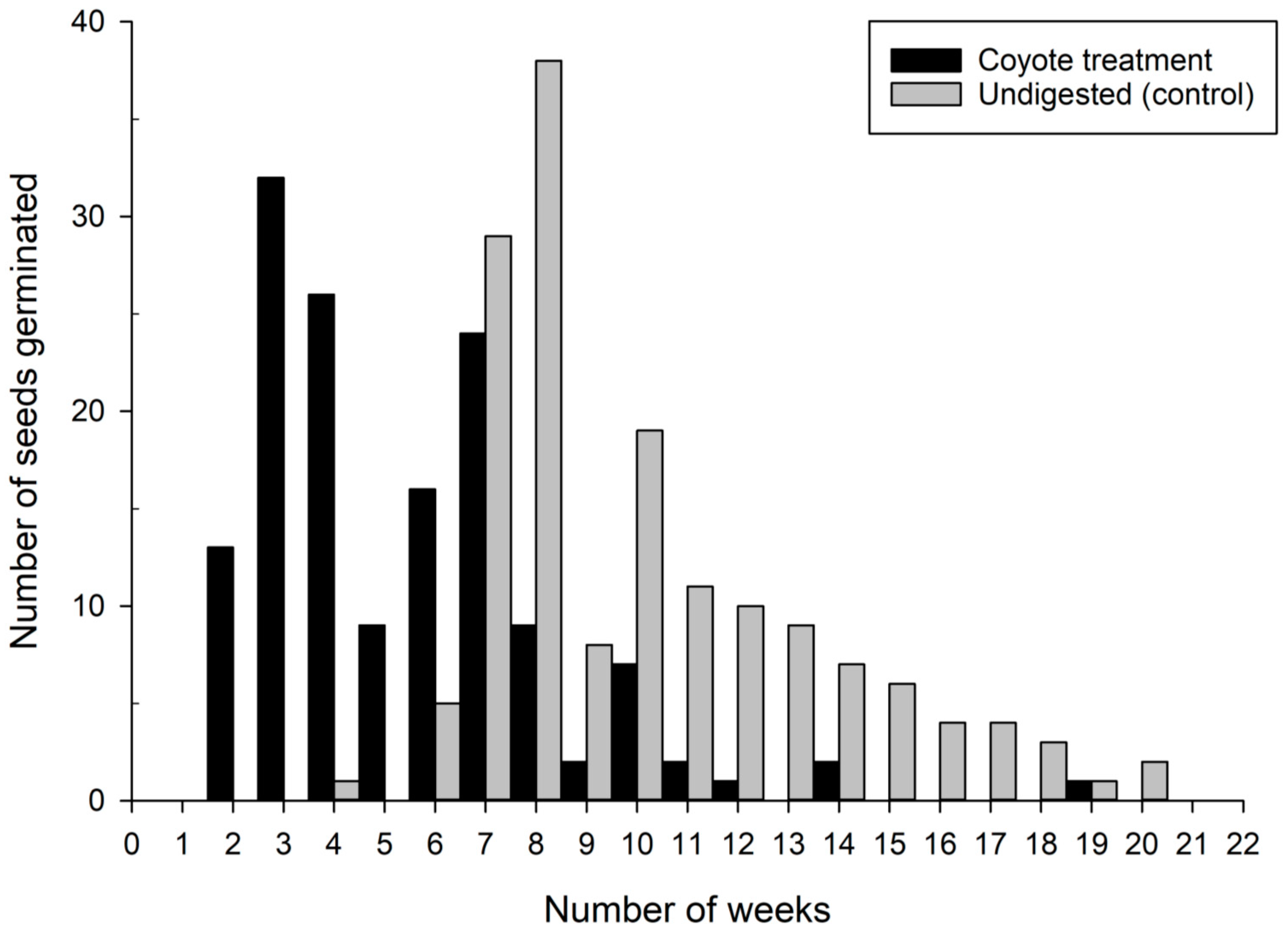
3. Results
4. Discussion
4.1. Germination Percentage
4.2. Germination Timing
4.3. Growth and Survival
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- van der Pijl, L. Principles of Dispersal in Higher Plants, 3rd ed.; Springer: Berlin, Germany, 1982. [Google Scholar]
- Howe, H.F.; Smallwood, J. Ecology of seed dispersal. Annu. Rev. Ecol. Syst. 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Willson, M.F. Mammals as seed-dispersal mutualists in North America. Oikos 1993, 67, 159–176. [Google Scholar] [CrossRef]
- Herrera, C.M. Plant-vertebrate seed dispersal systems in the Mediterranean: Ecological, evolutionary, and historical determinants. Annu. Rev. Ecol. Syst. 1995, 26, 705–727. [Google Scholar] [CrossRef]
- Traba, J.; Arrieta, S.; Herranz, J.; Clamagirand, M.C. Red fox (Vulpes vulpes L.) favour seed dispersal, germination and seedling survival of Mediterranean hackberry (Celtis australis L.). Acta Oecol. 2006, 30, 39–45. [Google Scholar]
- Fedriani, J.M.; Delibes, M. Functional diversity in fruit-frugivore interactions: A field experiment with Mediterranean mammals. Ecography 2009, 32, 983–992. [Google Scholar] [CrossRef]
- Izhaki, I.; Safriel, U. The effect of some Mediterranean scrubland frugivores upon germination patterns. J. Ecol. 1990, 78, 56–65. [Google Scholar] [CrossRef]
- Nilsson, P.; Fagerström, T.; Tuomi, J.; Åström, M. Does seed dormancy benefit the mother plant by reducing sib competition? Evol. Ecol. 1994, 8, 422–430. [Google Scholar] [CrossRef]
- Simons, A.M. Selection for increased allocation to offspring number under environmental unpredictability. J. Evol. Biol. 2007, 20, 813–817. [Google Scholar] [CrossRef]
- Murray, K.; Russell, S.; Picone, C.; Winnett-Murray, K.; Sherwood, W.; Kuhlmann, M. Fruit laxatives and seed passage rates in frugivores: Consequences for plant reproductive success. Ecology 1994, 75, 989–994. [Google Scholar] [CrossRef]
- DeBolt, A.M. Celtis reticulata Torr. In Wildland Shrubs of the United States and Its Territories: Thamnic Descriptions; Francis, J.K., Ed.; Gen. Tech. Rep. IITF-GTR-26; US Department of Agriculture, International Institute of Tropical Forestry, Rocky Mountain Research Station: San Juan, PR, USA; Fort Collins, CO, USA, 2004; Volume 1, pp. 167–169. [Google Scholar]
- Hayward, C.L. Biotic communities of the Wasatch chaparral, Utah. Ecol. Monogr. 1948, 18, 473–506. [Google Scholar] [CrossRef]
- DeBolt, A.M.; McCune, B. Is netleaf hackberry a viable rehabilitation species for Idaho rangelands? In Proceedings: Wildland Shrub and Arid Land Restoration Symposium; Roundy, B.A., McArthur, E.D., Haley, J.S., Mann, D.K., Eds.; Gen. Tech. Rep. INT-GTR-315; U.S. Department of Agriculture: Ogden, UT, USA, 1995; pp. 305–309. [Google Scholar]
- DeBolt, A.M.; McCune, B. Ecology of Celtis reticulata in Idaho. Great Basin Nat. 1995, 55, 237–248. [Google Scholar]
- Bonner, F.T. Celtis L. hackberry. In Seeds of Woody Plants in the United States, Handbook 450; Shopmeyer, C.S., Ed.; U.S. Department of Agriculture: Washington, DC, USA, 1974; pp. 298–300. [Google Scholar]
- DeBolt, A.M. The Ecology of Celtis reticulata Torr. (Netleaf Hackberry) in Idaho. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 1992. [Google Scholar]
- Stevens, M.T.; Holland, D.L.; Tanner, N.V. Netleaf hackberry seeds planted near boulders in the foothills of the Wasatch Mountains: Germination, survival, and patterns of establishment. West. N. Am. Nat. 2016, 76, 452–458. [Google Scholar] [CrossRef]
- Stevens, M.T.; Holland, D.L. Germination of netleaf hackberry seeds on the north sides of boulders: A shift away from patterns of mature hackberry distribution? West. N. Am. Nat. 2017, 77, 272–274. [Google Scholar] [CrossRef]
- Lewis Ivey, M.L.; Adkikari, A.; Graham, C.; Malekian, F.; Fontenot, K. Vegetable Seed Sanitation: Best Practices to Ensure On-farm Food Safety; Louisiana Department of Agriculture and Forestry: Baton Rouge, LA, USA, 2015; pp. 1–2. [Google Scholar]
- Traveset, A.; Robertson, A.; Rodríguez-Pérez, J. A review on the role of endozoochory in seed germination. In Seed Dispersal: Theory and Its Application in a Changing World; Dennis, A.J., Schupp, E.W., Green, R.J., Westcott, D.A., Eds.; CAB International: Wallingford, UK, 2007; pp. 78–103. [Google Scholar]
- Traveset, A.; Riera, N.; Mas, R.E. Ecology of fruit-colour polymorphism in Myrtus communis and differential effects of birds and mammals on seed germination and seedling growth. J. Ecol. 2001, 89, 749–760. [Google Scholar] [CrossRef] [Green Version]
- Traveset, A.; Verdú, M. A meta-analysis of the effect of gut treatment on seed germination. In Seed Dispersal and Frugivory: Ecology, Evolution, and Conservation; Levey, D.J., Silva, W.R., Galetti, M., Eds.; CAB International: Wallingford, UK, 2002; pp. 339–350. [Google Scholar]
- Cypher, B.L.; Cypher, E.A. Germination rates of tree seeds ingested by coyotes and raccoons. Am. Midl. Nat. 1999, 142, 71–76. [Google Scholar] [CrossRef]
- Harper, J.L. Population Biology of Plants; Academic Press: New York, NY, USA, 1977. [Google Scholar]
- Verdú, M.; Traveset, A. Early emergence enhances plant fitness: A phylogenetically controlled meta-analysis. Ecology 2005, 86, 1385–1394. [Google Scholar] [CrossRef]
- de Luis, M.; Verdú, M.; Raventós, J. Early to rise makes a plant healthy, wealthy, and wise. Ecology 2008, 89, 3061–3071. [Google Scholar] [CrossRef]
- Schupp, E.W. Quantity, quality and the effectiveness of seed dispersal by animals. Vegetatio 1993, 107, 15–29. [Google Scholar]
- Ross, M.A.; Harper, J.L. Occupation of biological space during seedling establishment. J. Ecol. 1972, 60, 77–88. [Google Scholar] [CrossRef]
- Seiwa, K. Advantages of early germination for growth and survival of seedlings of Acer mono under different overstorey phenologies in deciduous broad-leaved forests. J. Ecol. 1998, 86, 219–228. [Google Scholar] [CrossRef]
- Moreno, J.; Oechel, W. Factors controlling postfire seedling establishment in southern California chaparral. Oecologia 1992, 90, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Seiwa, K. Effects of seed size and emergence time on tree seedling establishment: Importance of developmental constraints. Oecologia 2000, 123, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Castro, J. Short delay in timing of emergence determines establishment success in Pinus sylvestris across microhabitats. Ann. Bot. 2006, 98, 1233–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathias, A.; Kisdi, E. Adaptive diversification of germination strategies. Proc. Roy. Soc. B-Biol. Sci. 2002, 269, 151–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlowski, T.T. Growth and Development of Trees; Seed Germination, Ontogeny and Shoot Growth; Academic Press: New York, NY, USA, 1971; Volume 1, pp. 164–206. [Google Scholar]
- Peattie, D.C. A Natural History of Western Trees; Houghton Mifflin Harcourt: Boston, MA, USA, 1991. [Google Scholar]
- Lanner, R.M. Trees of the Great Basin: A Natural History; University of Nevada Press: Reno, NV, USA, 1984. [Google Scholar]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stevens, M.T.; Houghton, S.; Veltkamp, H.A. Frugivory by Coyotes Decreases the Time to Germination and Increases the Growth of Netleaf Hackberry (Celtis reticulata) Seedlings. Forests 2020, 11, 727. https://doi.org/10.3390/f11070727
Stevens MT, Houghton S, Veltkamp HA. Frugivory by Coyotes Decreases the Time to Germination and Increases the Growth of Netleaf Hackberry (Celtis reticulata) Seedlings. Forests. 2020; 11(7):727. https://doi.org/10.3390/f11070727
Chicago/Turabian StyleStevens, Michael T., Sydney Houghton, and Hannah A. Veltkamp. 2020. "Frugivory by Coyotes Decreases the Time to Germination and Increases the Growth of Netleaf Hackberry (Celtis reticulata) Seedlings" Forests 11, no. 7: 727. https://doi.org/10.3390/f11070727
APA StyleStevens, M. T., Houghton, S., & Veltkamp, H. A. (2020). Frugivory by Coyotes Decreases the Time to Germination and Increases the Growth of Netleaf Hackberry (Celtis reticulata) Seedlings. Forests, 11(7), 727. https://doi.org/10.3390/f11070727