4.1. Regeneration Success after 15 Years
This study was first designed to evaluate the success of post-harvest regeneration within canopy openings of two different sizes, created in an operational context over a large territory (
Figure 1) through either group selection cutting or patch cutting, both followed by soil scarification. Most openings had a stocking for commercial species above 40% after 15 years, based on 6.25 m
2 quadrats, in both opening types and in all three soil scarification methods (
Figure 2g). This is easily above the 20% stocking threshold required by the Quebec management guidelines [
26]. A stocking of 40% obtained with sampling units of that size would mean that there is at least one commercial species located in 640 out of the 1600 possible sampling units per hectare (10,000 m
2 ÷ 6.25 m
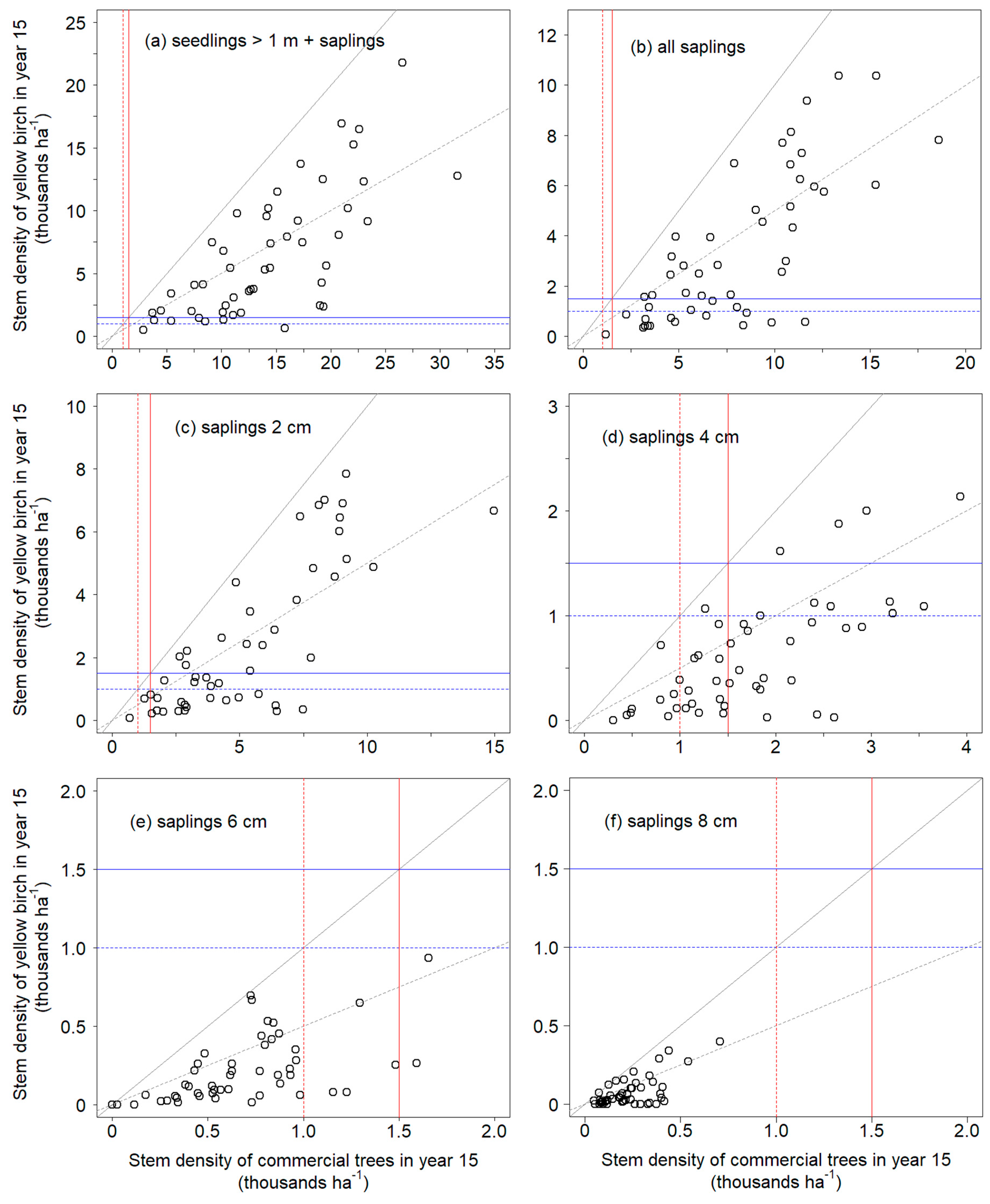
2 = 1600), thus resulting in a tree density of at least 640 stems properly spaced per hectare. In addition, every cluster had a density of commercial species of at least 2500 stems ≥ 1 m in height ha
−1, well above the recommended stocking of 1500 stems ha
−1 from Leak et al. [
34] (
Figure 5a). When considering saplings only, all clusters had densities above the minimum stocking of 1000 stems ha
−1 (
Figure 5b). As suggested by Elenitsky et al. [
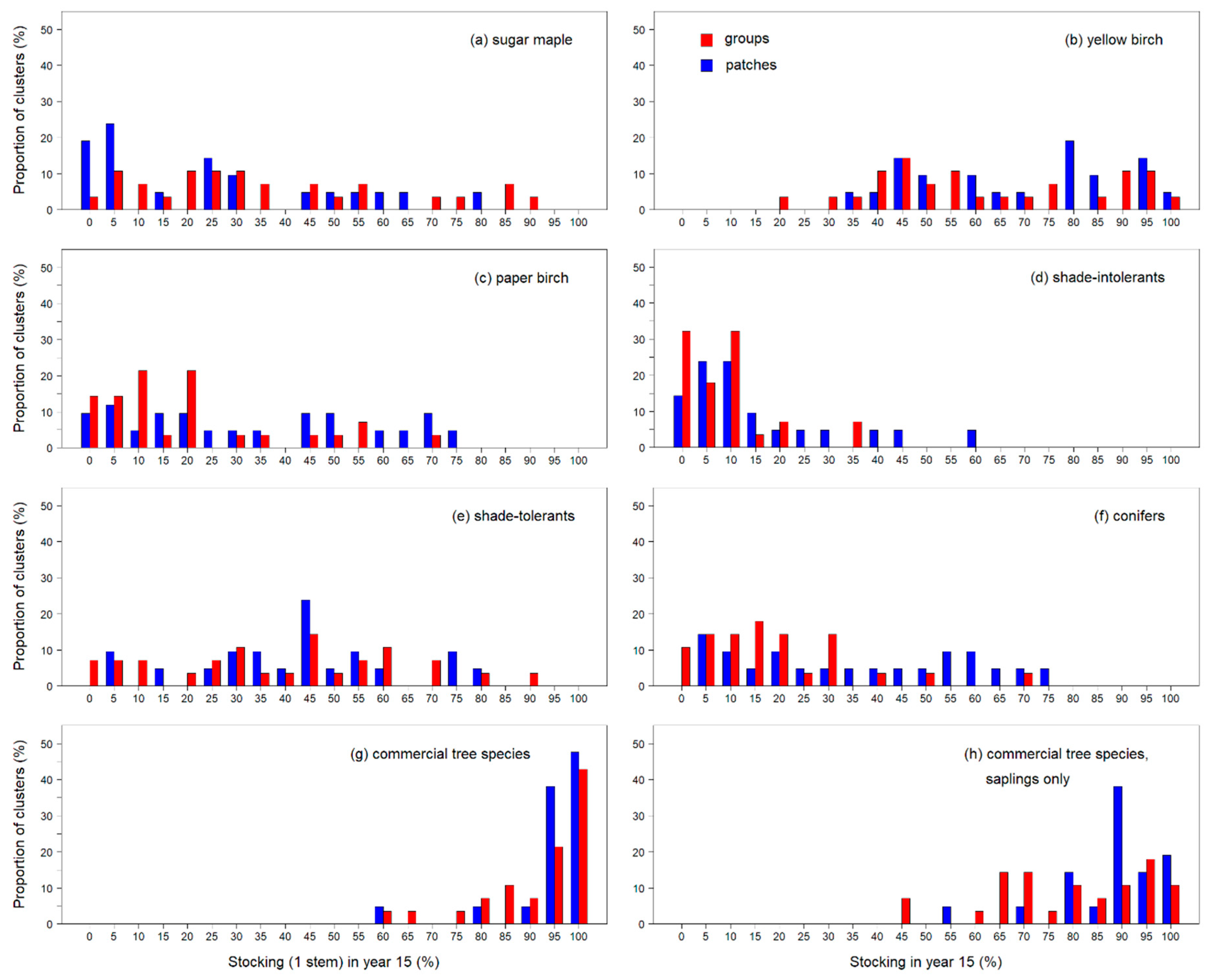
66], if we assume that (i) mortality is low as trees grow from saplings to 10-cm DBH, (ii) that saplings have transcended both deer browsing and shrub competition, and (iii) that a certain amount of seedlings ≥ 1 m in height will recruit into saplings in the near future, it would thus be sufficient to expect an eventual crown closure. Yellow birch, the main target species for natural regeneration and the most abundant commercial species in the current study, was present in all canopy openings, but with a highly variable distribution (mean Stock-1 = 62.5%, SD = 23, range = 15–100%,
Figure 2b), as was also the case for sugar maple and the other commercial species pools (
Figure 2a,c–f). Nevertheless, Stock-1 values for YB in all clusters were above the required thresholds from the Quebec management guidelines [
26]. Additionally, mean densities of YB were always above the threshold of 1500 stems ha
−1 recommended by Leak et al. [
34] for a successful regeneration (
Figure 5a).
The fact that these canopy openings showed satisfyingly high mean densities, but sometimes had lower stocking of commercial species, especially saplings, indicate that those trees are abundant but not always so well distributed within openings. As a matter of fact, this has been observed elsewhere: in 9-year-old canopy openings created through group selection cutting, Poznanovic et al. [
23] documented the spatial aggregation of YB, which they ascribed to spatial heterogeneity of suitable microsites. Six years later at these same sites, thus 15 years after harvest, Knapp et al. [
67] observed that over half of the canopy openings they studied, ranging in size from 267 to 1192 m
2, harbored 10- to 310-m
2 portions of their area where regeneration, regardless of species, was no taller than 1.4 m. In comparison, they reported that 86% of saplings (all species combined) showed heights between 0.5 and 5 m. Hence, they concluded that their openings exhibited ‘spatially uneven patterns of regeneration, with areas of persistent and dense shrubs’ [
67]. We observed this type of heterogeneity on the field in some of our canopy openings, often associated with a somewhat poorly drain soil, a very stony soil or intense cervid browsing, but these three hypotheses remain to be tested.
In our study, canopy openings created through harvest were of two very different sizes classes (1000 m
2 for groups vs. 15,000 m
2 for patches, on average). Yet, YB regeneration did not differ between the two size classes. Previous authors have similarly observed no effect of gap sizes on the regeneration success of YB [
21,
47,
48,
50,
68] despite establishing experimental gradients of gap sizes varying in surface area from 260 to 1600 m
2, and especially in studies with favorable microsites to YB germination covering on average 50% of the area [
21,
47,
48,
68]—a scarification result similar to that at our sites, see Beaudet et al. [
52]. Other studies nonetheless reported effects of gap size on YB regeneration success, although they used a gradient that reached down to very small sizes (<300 m
2) and were located in stands where shade-tolerant species such as sugar maple and American beech were abundant [
21,
67,
68,
69,
70] which was not often the case in this study. In addition, Bolton and D’Amato [
21] as well as Shields et al. [
68] acknowledged that the minimal soil disturbance that occurred in their gaps was not sufficient to create suitable germination microsites for YB. Incidentally, such factors (small gaps, abundance of shade-tolerants, and no soil scarification) are often reunited in studies where YB regeneration was the least successful, with relative stem densities of 5% or less [
67,
69,
70].
The stem densities of yellow birch reached in our study are comparable to those of Shabaga et al. [
71] in the neighboring Canadian province of Ontario. They reported after 10 years mean YB densities of 7000 stems ha
−1 for the ‘large regeneration’, which in their case comprised all individuals > 50 cm in height. Yet they observed large variation between ‘systematic gaps’ (≈5000 stems ha
−1) and ‘traditional gaps’ (≈14,000 stems ha
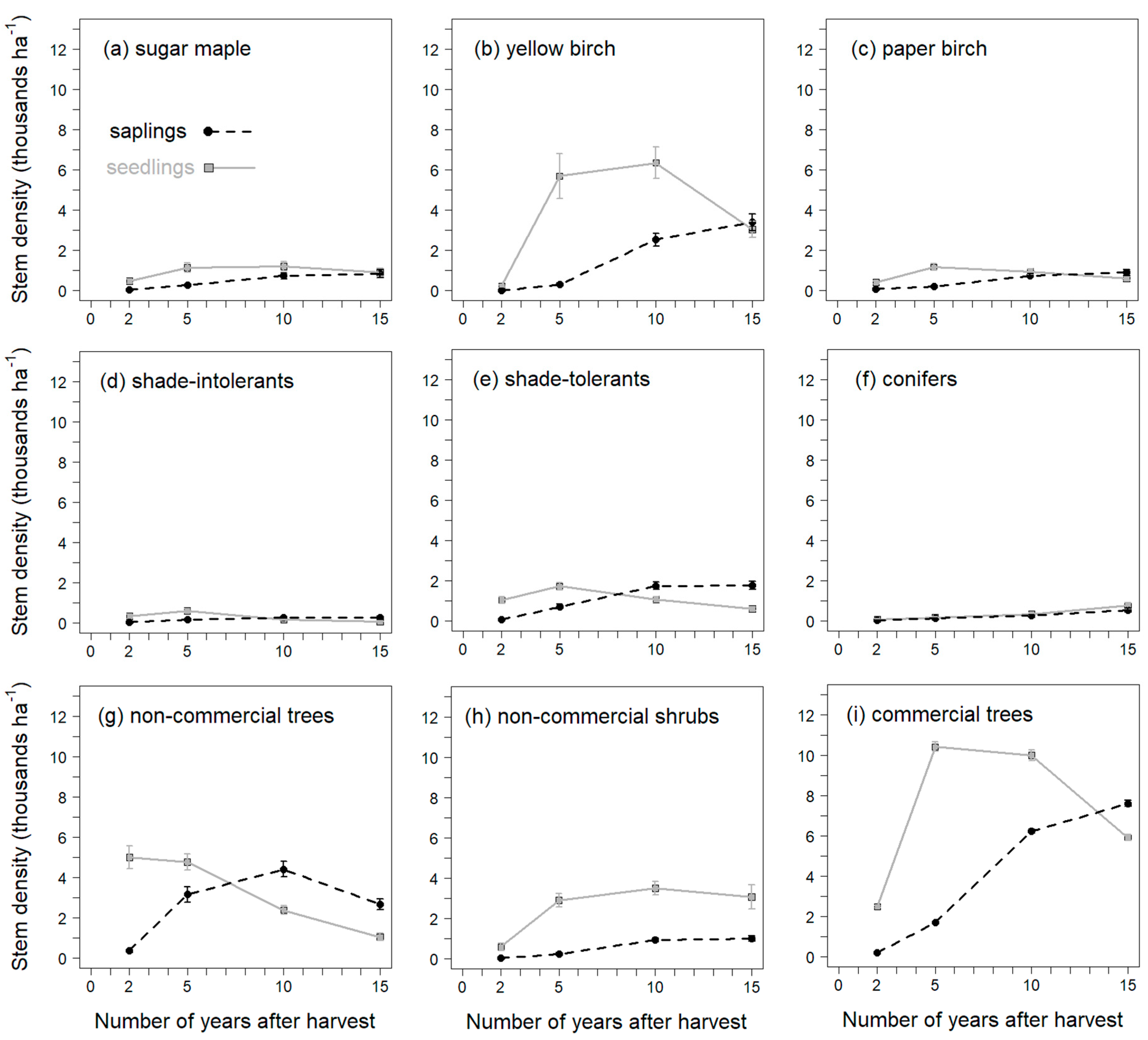
−1). By traditional gaps, they meant gaps placed where favorable microsites as well as mature seed trees could be found, in opposition to systematic gaps that would be located along a fixed grid. In the present study, canopy openings were systematically positioned within a stand, although there were always several seed trees present around an opening. In year 10, we observed mean YB densities of 8865 stems ha
−1 (6350 seedlings ≥ 1 m in height plus 2515 saplings;
Figure 4b). Given the difference in how developmental stages were grouped between Shabaga et al.’s [
71] study and ours, we consider our numbers to be quite on par with theirs.
However, much higher densities than the mean of the present study were reported by Bédard and DeBlois [
47] and Gauthier et al. [
48] in experimental canopy openings ranging from 15 to 35 m in diameter in Quebec. Gauthier et al. [
48] reported over 12,000 large (≥ 1 m in height) YB seedlings ha
−1 5 years after a harvest that occurred in 2005 (in comparison to 5680 YB seedlings ha
−1 in our study), while for SM in the same height class they observed densities > 20,000 ha
−1 (1130 ha
−1 in our study). Bédard and DeBlois [
47] observed stem densities of YB ≥ 1 m ranging from ≈13,000 large seedlings ha
−1 in fenced deer-exclusion openings to ≈2000 ha
−1 in non-fenced openings 5 years after a harvest that took place in 2000. Although the mean YB density in year 5 across all our studied openings is lower than in the two aforementioned studies, we did observe such high YB densities in a few cases (data not shown). Soil scarification in these two experimental studies was synchronized with a good seed year, while our study covered all the years from 2000 to 2005. The lack of synchronization with a good seed year has been shown to potentially limit the efficacy of scarification to regenerate YB in an experimental study [
72]. Nonetheless, we did not observe any obvious annual variation in YB regeneration in our study that could be explained by differences in seed years (data not shown). As a matter of fact, the receptivity of soil microsites to YB regeneration can reach up to 3 years [
73]. Regarding relative abundance, both Gauthier et al. [
48] and Bédard and DeBlois [
47] reported values of 21% for YB in year 5. In comparison, our study sites presented in year 5 a mean %YB of 16% (SD = 13, range = 0.2 to 76%) for stems ≥ 1 m in height, which increased by year 15 to 27% (SD = 17, range = 4 to 73%).
The first regeneration survey, conducted in the second year after soil scarification, presented a situation where the non-commercial trees and shrubs dominated stem density (
Figure 4). These are either shade-intolerant, pioneer species that develop rapidly after the disturbance created by harvesting operations, or alternatively are pre-established shade-tolerants able to take advantage of the new opening and enhanced light availability [
74,
75]. Given that these first surveys are aimed in part at identifying potential regeneration failures requiring immediate actions to address and correct the issue, the outlook may be gloomy when target species are few within a regrowth of weeds. Nyland et al. [
75] indeed warned that a strong early occupation of regenerating sites by pin cherry (
Prunus pensylvanica) could become problematic. They defined the threshold as three individuals over 1 m in height per 4 m
2 (their actual numbers were three pin cherry over 3 ft per milacre). This would correspond to 7500 stems ha
−1. At our study sites, mean densities of NCOM-tree reached 7900 stems ha
−1 in year 5 (
Figure 4g) and quickly decreased afterwards. However, that species pool was not composed only of pin cherry but also of two other species, mountain maple and striped maple. These three species, respectively, represented on average 28%, 50%, and 22% of the NCOM-tree density (data not shown). Overall, given successional dynamics of tree species in temperate hardwood forests, combined to the short life expectancy of several pioneers, desirable hardwoods eventually overgrow their competitors unless dominance by non-commercial trees is very strong [
32]. Previous authors have similarly reported that early presence of short-lived competitors such as
Rubus spp. did not have lasting impact on YB development [
71,
76,
77].
The slight decrease in vertical dominance of SM, YB, INTOL, and TOL between year-2 and year-5, concurrent with the increase in dominance of NCOM-tree and NCOM-shrub, likely reflects the growth dynamics of the latter species (
Table 6). The fact that dominance by SM and TOL did not increase greatly between year 2 and year 15 might be due to them being established prior to harvest, as well as not being able to profit from the canopy opening as much as YB did. Indeed, in that regard, YB was the most successful of the hardwood species, becoming increasingly dominant as the years passed, to the point that by year 15 it was the most frequently dominant species (
Table 6). Quite contrasting results were reported by Poznanovic et al. [
70]: between years-2 and -9 after group selection harvest, YB went from being present in 14% of sampling units down to 12%. This meant that a certain number of subplots occupied by that species in year 2 had none in year 9. Therefore, in their case, the issue was not whether YB was dominant, but whether it was present, since it represented at best 5.5% of total sapling density, rather dominated by sugar maple [
70]. Later studies at these sites suggested that soil scarification would have been needed to better promote mid-tolerant and intolerant hardwoods [
67].
Height or developmental stage attained by a target species at a certain age may also prove a valuable parameter in assessing whether post-harvest regeneration was successful. For instance, some experimental studies have reported heights for successfully regenerated YB within canopy openings: between 2.45 and 7.0 m in year 10 [
69], or mostly below 4.5 m in year 9 [
70], or between 6.0 and 10.5 m in year 15 after harvest [
78]. In the present study, the highest mean-per-cluster maximum height for YB in year 15 was 8.10 m, with a mean of 3.23 m across all clusters (SD = 1.74;
Figure S3). In that regard, our study would not be among the most successful. As a matter of fact, YB was not very often the tallest individual within a quadrat (
Figure 3b), even though dominance increased noticeably over time (
Table 6). Indeed, hardwood seedlings dominated by competitors in early years may turn out to become dominant at a later point in stand development. For example, Marquis [
40] had observed that some sites harvested using patch cutting in order to promote birches (yellow + paper) were overcrowded with pin cherry 3 years after harvest. However, returning at the same sites 47 years later, Leak [
79] reported that birches composed 60% of the dominant trees in these patches.
4.2. Predicting Future Stand Composition
Although predicting the future stand composition remains challenging, our study showed that the later abundance of target species within post-harvest regenerating canopy openings can be approximated using some early basic inventory metrics. The better performance of the 6.25 m2 quadrat over the 25 m2 plot indicated that smaller plots, such as the more common 4 m² plots (or milacre), are better. In addition, the generally higher coefficients of correlation obtained with observations in year 5, instead of year 2, advocate waiting until year 5 to evaluate the regeneration success.
We first investigated whether the presence of a species in a dominant position in a quadrat in year 2 or 5 was a good indicator of the species that will dominate the quadrat in year 15. In the best case, 83% of the quadrats dominated by a non-commercial tree in year 15 were already dominated by this species pool in year 5. At the opposite, only 33% of the quadrats dominated by either YB or a conifer in year 15 were already in this situation in year 5. Yet, when a non-commercial tree dominated in year 5, there was only a 46% chance for it to still be dominant in year 15, whereas a quadrat dominated by a YB in year 5 would still be dominated by that species in year 15 in 69% of cases. Consequently, this analysis of dominance led us to a certain number of observations: (1) early dominance within a quadrat is not necessarily representative of later dominance within the same quadrat; (2) early surveys (year 2) do not help much in predicting which species will occupy the top of the future canopy within the quadrat; (3) however, the fact that target species are overgrown by competitors in early years does not preclude them becoming more prominent over time; (4) the non-commercial tree species that are highly abundant after a harvest and rapidly occupy the upper layer of the developing canopy, thus competing strongly against commercial hardwoods, see their dominant presence receding over time.
Northern hardwoods being mid- to late-successional species, it is normal to see a delay in their accession to canopy dominance, as previously showed by McClure et al. [
80] in 44- to 48-year-old canopy openings. Yet, these authors observed that all canopy trees had established no later than 4 years after opening creation, although the exact moment of establishment varied per species. Indeed, they stated that 75% of canopy sugar maples had been advance regeneration whereas 79% of canopy yellow birches had germinated only after opening creation. Hence, early years may be crucial for regenerating openings, since future canopy trees need to at least be present even though they are not yet dominant. For instance, in a study assessing regeneration after strip cutting in Massachusetts, USA, Allison et al. (2003) observed that pin cherry (
Prunus pensylvanica) had been the most abundant species to regenerate immediately after harvest. However, four decades later, it had almost disappeared, being replaced by, in decreasing order of abundance: red maple, paper birch, sugar maple, and black birch (
Betula lenta, a mid-tolerant species like yellow birch [
12]). This shows that a harvested site dominated in early years by non-commercial, competing woody species may nonetheless mostly comprise desirable species in later years. Our data suggest a similar phenomenon over a 15-year time period. Adequate silvicultural interventions, such as scarifying the soil, may accelerate the post-harvest succession.
We identified some pertinent variables that could be measured in surveys conducted 5 years after the last treatment, either harvest or soil scarification, to better predict the composition relative abundance or relative basal area) in year 15. However, we did not delve into comparing how much time in the field must be dedicated to measuring each of these variables vs. their respective predictive power. The exact sets of variables differed slightly depending on the species under scrutiny, but %YB (or %SM) and Stock-1 in year 5 were always useful, except for Stock-1 in explaining %SM (
Table 3 and
Table 4). These variables indicate that abundant YB regeneration in year 15 can be obtained when it is well distributed within the opening and at a relatively high density among the stems at least 1 m in height. For example, if we would have created these openings with the aim of obtaining at least 50% of YB among all commercial species in year 15, it would have required the combination of a YB stocking between 20% and 100% with a relative abundance ranging from 35% to 80% (Equations (5) and (6)). More specifically, Equations (5) and (6) inform us that when Stock-1 ranges from 20% to 100% in year 5 (i.e., the value range observed at our sites for YB in that year), %YB in that same year must range from 50% to 35%, respectively, for the aforementioned Stock-1 values, in order to obtain 50% YB in BA or abundance in year 15. Only 16% of our clusters had a %YB in year 5 greater than 35% (with the highest value being 80%), and they concurrently had a Stock-1 between 71% and 100%. Obtaining a %YB above 50% in year 15 would thus require proportionally higher %YB in year 5, but could be achieved from a wide range of Stock-1 values. It demonstrates how %YB in stems ≥ 1 m in height was the most important indicator of future YB abundance. In the case of SM, the variables %SM, StD, Stock-1≥1m, and Stock-tallest in year 5 were identified as interesting predictors (Equations (9) and (10)). The models were more complex than for YB, but %SM of stems ≥ 1 m in height in year 5 was again the most important variable explaining variations in year 15 abundance. SM was less abundant within our canopy openings compared with YB, peaking at %SM values of 40% in year 5 and 60% in year 15 (
Figure 7). Consequently, regeneration objectives would need to be adjusted: for instance, a %SM above 20% in year 15 would require year 5 variables in the order of 50% for Stock-1≥1m, 40% for Stock-tallest, and 20% for %SM.
In a paper comparing early post-harvest regeneration to stand composition ≈50 years later, Leak [
37] reported that later relative basal area of pole-sized SM was best predicted by early relative number of SM saplings. This same measurement based on seedlings was not an accurate predictor of relative basal area of poles, because seedlings were so abundant in early years that they greatly overestimated later presence of SM poles. For YB, Leak [
37] concluded that relative basal area after 50 years was most related to the early-year stocking based on the dominant (i.e., tallest) stem. In our study, we observed on the contrary that year-15 YB proportional presence was not related to Stock-tallest measured in year 2 or 5, but that this measure was appropriate for SM. These differences might be due to the differing types of harvest (Leak’s single-tree selection and diameter-limit cuttings vs. our group selection and patch cuttings) and the respective sizes of the resulting canopy openings.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}