Biomass and Volume Modeling along with Carbon Concentration Variations of Short-Rotation Poplar Plantations




Abstract
1. Introduction
2. Materials and Methods
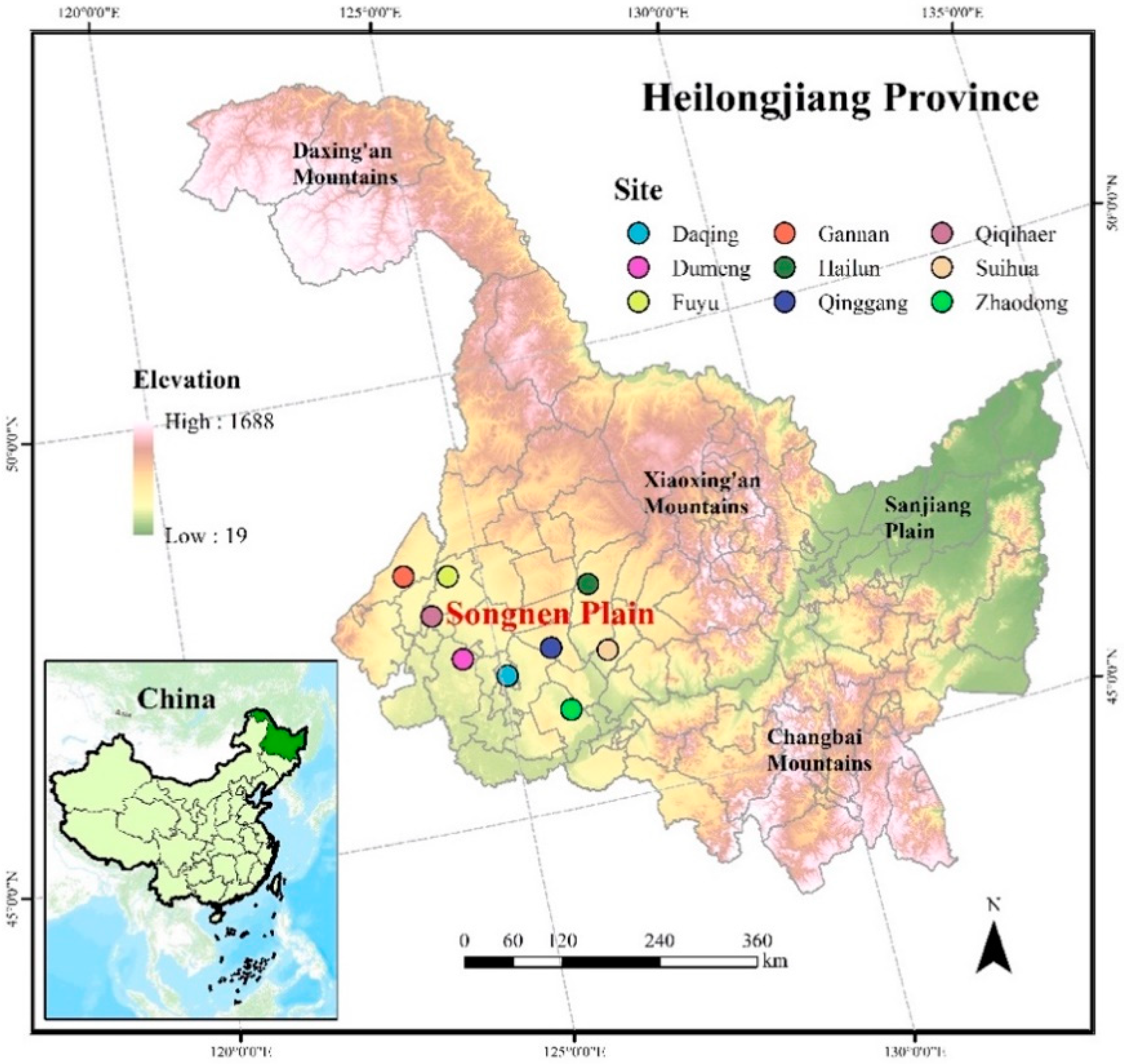
2.1. Site Description
2.2. Data Collection
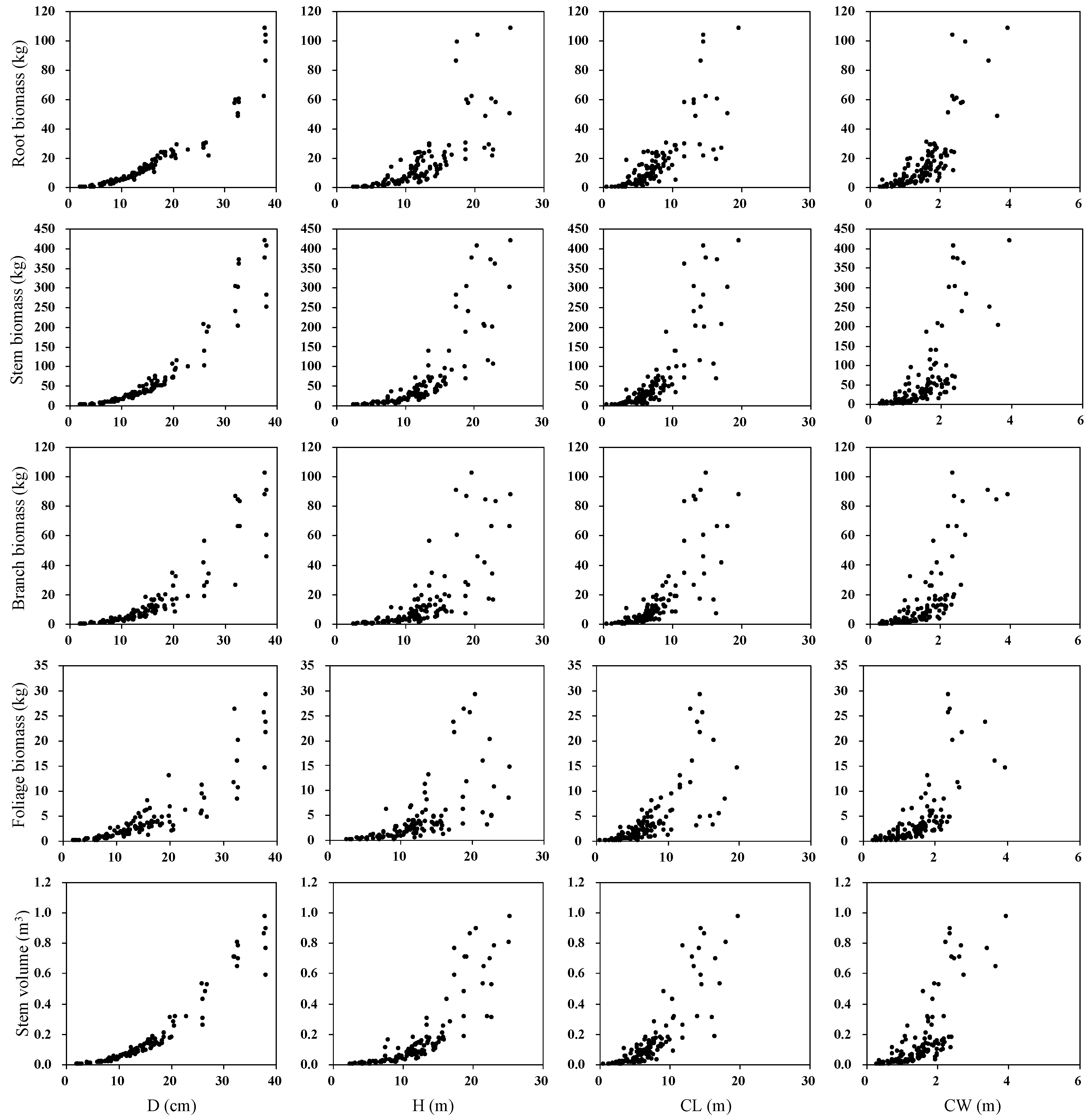
2.2.1. Tree Measurements
2.2.2. Biomass Measurements
2.2.3. Stem Volume and Biomass Conversion and Expansion Factor Measurements
2.2.4. Carbon Concentration Measurements
2.3. Statistical Analysis
2.3.1. Variable Selection of Biomass and Volume Models
2.3.2. Additive Biomass Model System
2.3.3. Weighting Function for Heteroscedasticity
2.3.4. Model Fitting and Evaluation
2.3.5. Effects of Tree Sizes (D) and Components on Carbon Concentration
3. Results
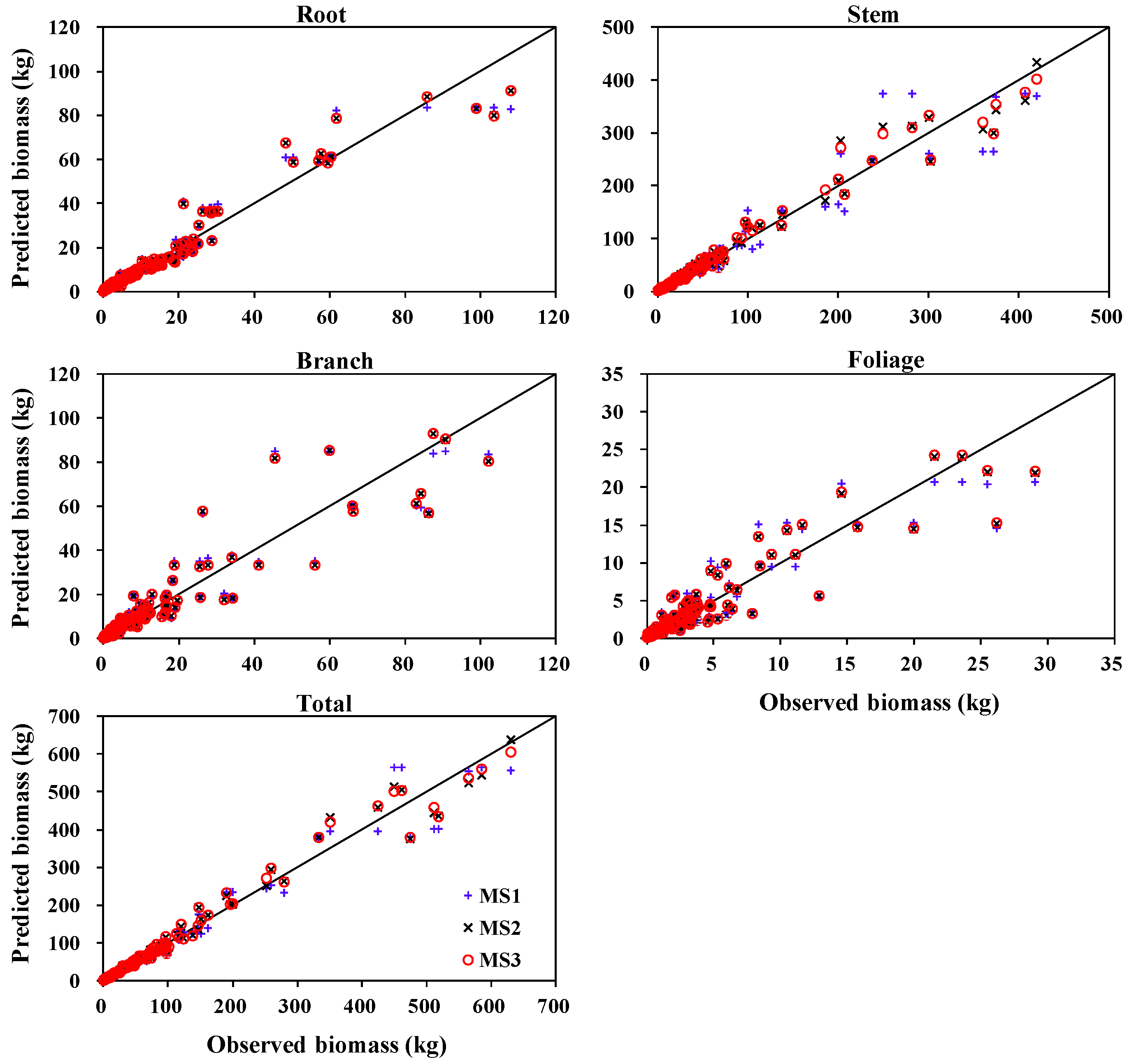
3.1. Biomass Equations
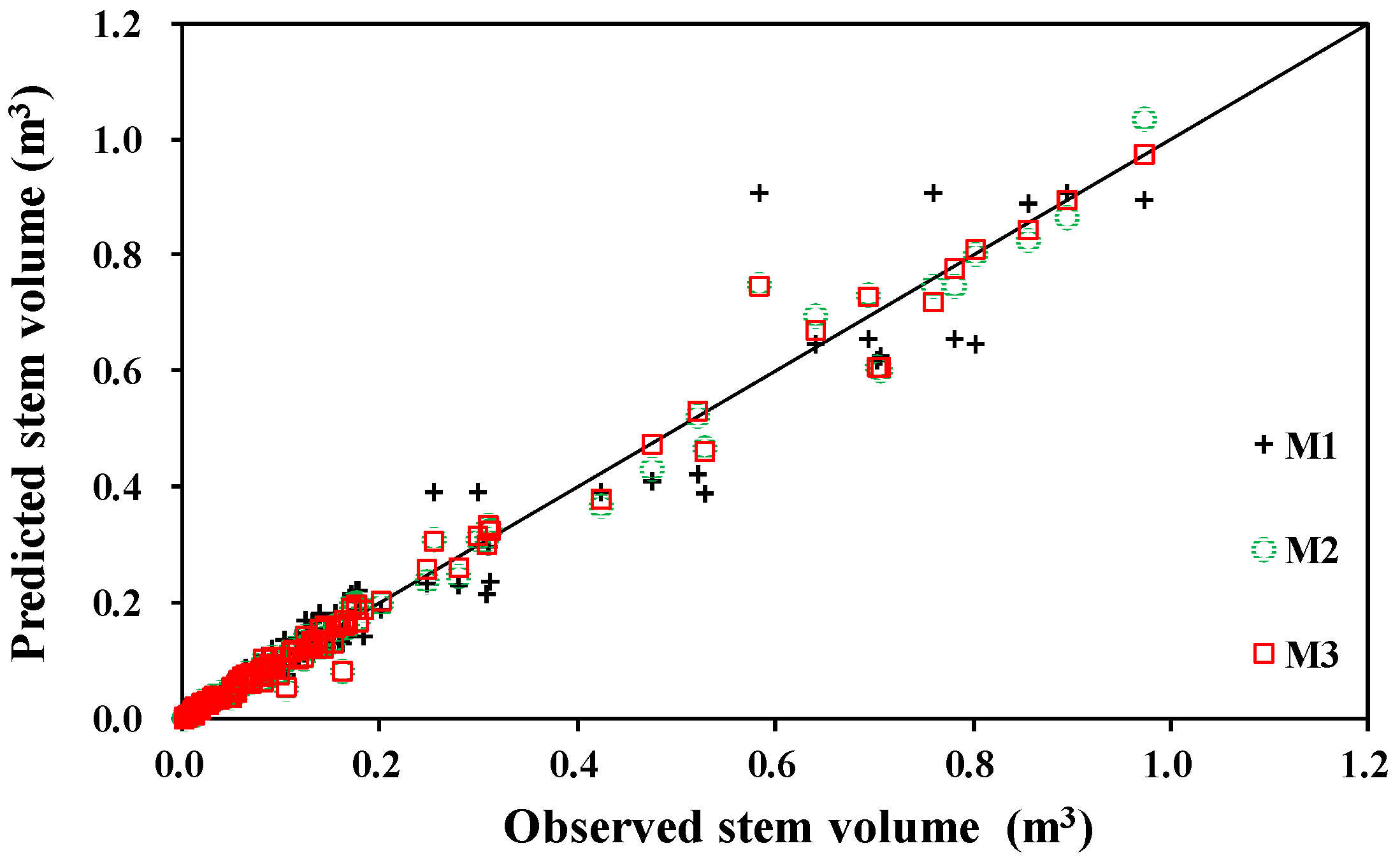
3.2. Stem Volume Equations
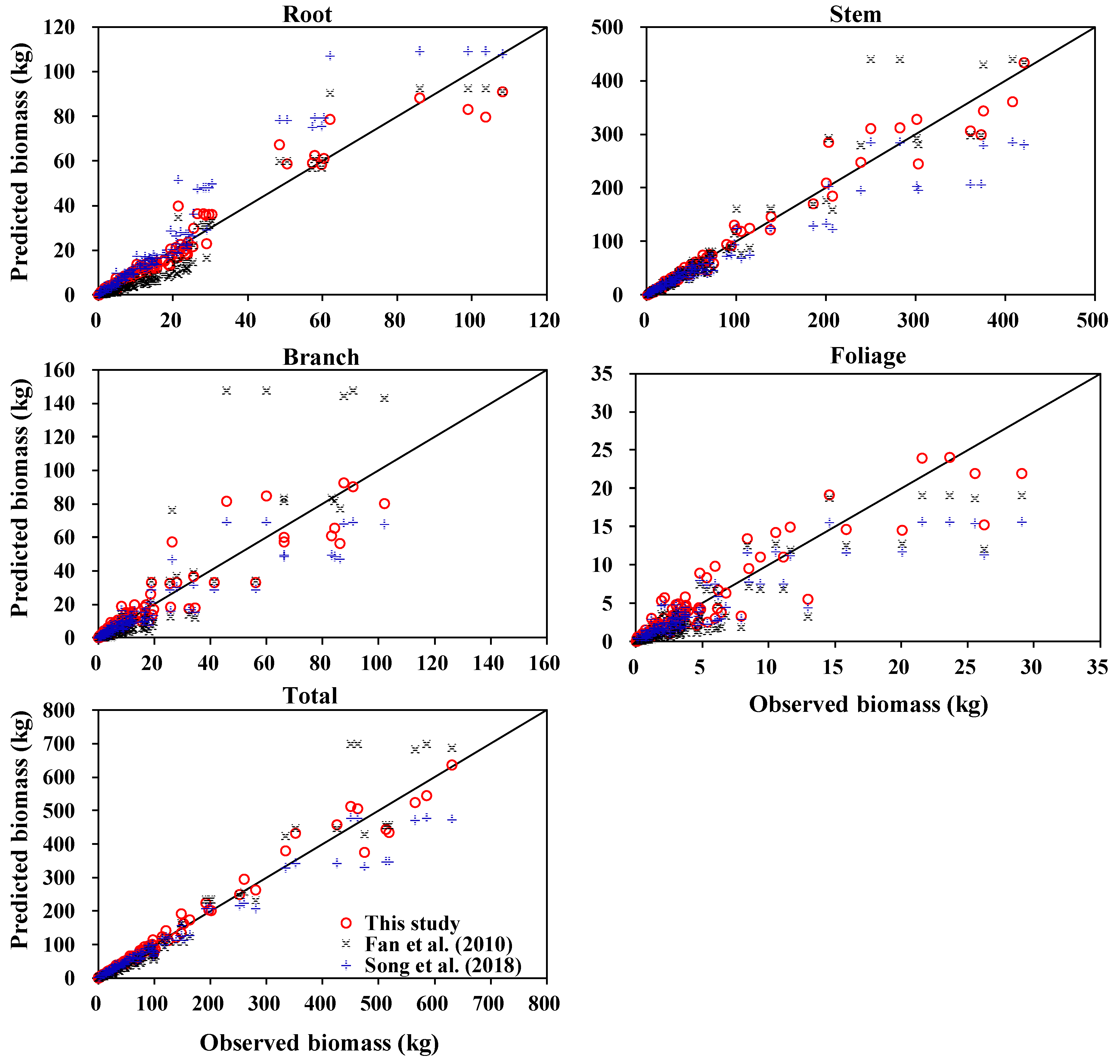
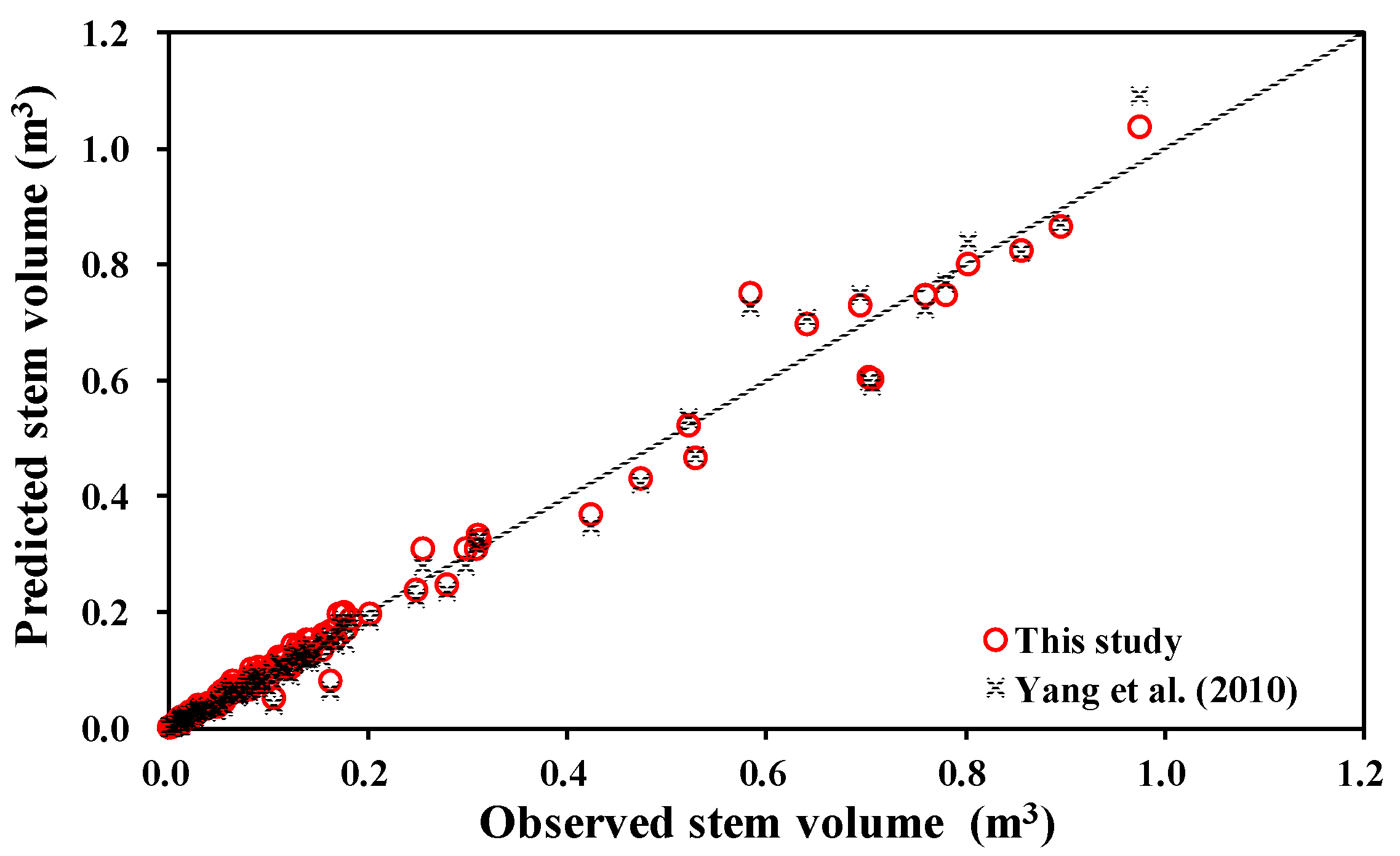
3.3. Comparison with Other Biomass and Stem Volume Models
3.4. Variations of Carbon Concentration
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Masse, S.; Marchand, P.P.; Bernier-Cardou, M. Forecasting the deployment of short-rotation intensive culture of willow or hybrid poplar: Insights from a Delphi study. Can. J. For. Res. 2014, 44, 422–431. [Google Scholar] [CrossRef]
- Parmar, K.; Keith, A.M.; Rowe, R.L.; Sohi, S.P.; Moeckel, C.; Pereira, M.G.; McNamara, N.P. Bioenergy driven land use change impacts on soil greenhouse gas regulation under Short Rotation Forestry. Biomass Bioenergy 2015, 82, 40–48. [Google Scholar] [CrossRef]
- Arevalo, C.B.M.; Bhatti, J.S.; Chang, S.X.; Sidders, D. Land use change effects on ecosystem carbon balance: From agricultural to hybrid poplar plantation. Agric. Ecosyst. Environ. 2011, 141, 342–349. [Google Scholar] [CrossRef]
- Don, A.; Osborne, B.; Hastings, A.; Skiba, U.; Carter, M.S.; Drewer, J.; Flessa, H.; Freibauer, A.; Hyvonen, N.; Jones, M.B.; et al. Land-use change to bioenergy production in Europe: Implications for the greenhouse gas balance and soil carbon. GCB Bioenergy 2012, 4, 372–391. [Google Scholar] [CrossRef]
- Lamers, P.; Hamelinck, C.; Junginger, M.; Faaij, A. International bioenergy trade-A review of past developments in the liquid biofuel market. Renew. Sust. Energ. Rev. 2011, 15, 2655–2676. [Google Scholar] [CrossRef]
- Werner, C.; Haas, E.; Grote, R.; Gauder, M.; Graeff-Hoenninger, S.; Claupein, W.; Butterbach-Bahl, K. Biomass production potential from Populus short rotation systems in Romania. GCB Bioenergy 2012, 4, 642–653. [Google Scholar] [CrossRef]
- Yan, M.; Wang, L.; Ren, H.; Zhang, X. Biomass production and carbon sequestration of a short-rotation forest with different poplar clones in northwest China. Sci. Total Environ. 2017, 586, 1135–1140. [Google Scholar]
- Lupi, C.; Larocque, G.; DesRochers, A.; Labrecque, M.; Mosseler, A.; Major, J.; Beaulieu, J.; Tremblay, F.; Gordon, A.M.; Thomas, B.R.; et al. Evaluating sampling designs and deriving biomass equations for young plantations of poplar and willow clones. Biomass Bioenergy 2015, 83, 196–205. [Google Scholar] [CrossRef]
- Oliveira, N.; Rodriguez-Soalleiro, R.; Perez-Cruzado, C.; Canellas, I.; Sixto, H.; Ceulemans, R. Above- and below-ground carbon accumulation and biomass allocation in poplar short rotation plantations under Mediterranean conditions. For. Ecol. Manag. 2018, 428, 57–65. [Google Scholar] [CrossRef]
- Li, S.; Zhang, Z.; He, C.; An, X. Progress on Hybridization Breeding of Poplar in China. World For. Res. 2004, 17, 37–41. [Google Scholar]
- Liang, W.; Hu, H.; Liu, F.; Zhang, D. Research advance of biomass and carbon storage of poplar in China. J. For. Res. 2006, 17, 75–79. [Google Scholar] [CrossRef]
- Jiang, Z.; Wang, X.; Fei, B.; Ren, H.; Liu, X. Effect of stand and tree attributes on growth and wood quality characteristics from a spacing trial with Populus xiaohei. Ann. For. Sci. 2007, 64, 807–814. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, Z.; Ren, H. Distribution of wet heartwood in stems of Populus xiaohei from a spacing trial. Scand. J. For. Res. 2008, 23, 38–45. [Google Scholar] [CrossRef]
- State Forestry and Grassland administration. Forest Resource Survey Report (2014–2018); China Forestry Publishing: Beijing, China, 2019; p. 451.
- Chang, Y.; Liu, G.; Jiang, J.; Wang, Y.; Wang, D. The Genetic Transformation of the Genes Resistant to Insect for Populus xiaohei. J. North-East For. Univ. 2004, 32, 30–31. [Google Scholar]
- Jiang, P.; Cui, L.; Qin, Z.; Wang, W.; Gu, J.; Wang, G. Tree Height and DBH Growth Models of Populus simonii × P. nigra. J. Northwest For. Univ. 2013, 28, 129–133. [Google Scholar]
- Jiang, Z.; Fan, S.; Feng, H.; Zhang, Q.; Liu, G.; Zong, Y. Biomass and Distribution Patterns of Populus xiaohei Plantation in Sandy Land of North China. Sci. Silvae Sin. 2007, 43, 15–20. [Google Scholar]
- Li, Z.; Zhao, X.; Yang, C.; Wang, G.; Wang, F.; Zhang, L.; Zhang, L.; Liu, G.; Jiang, J. Variation and growth adaptability analysis of transgenic Populus simonii × P. nigra lines carrying TaLEA gene. J. Beijing For. Univ. 2013, 35, 57–62. [Google Scholar]
- Burkhart, H.E.; Tomé, M. Modeling Forest Trees and Stands; Springer: New York, NY, USA, 2012; p. 458. [Google Scholar]
- Zhang, X.; Zhao, Y.; Ashton, M.S.; Lee, X. Measuring Carbon in Forests. In Managing Forest Carbon in a Changing Climate; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Brunori, A.; Dini, F.; Cantini, C.; Sala, G.; La Mantia, T.; Caruso, T.; Marra, F.P.; Trotta, C.; Nasini, L.; Regni, L.; et al. Biomass and volume modeling in Olea europaea L. cv “Leccino”. Trees-Struct. Funct. 2017, 31, 1859–1874. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Li, F. A Three-Step Proportional Weighting System of Nonlinear Biomass Equations. For. Sci. 2015, 61, 35–45. [Google Scholar] [CrossRef]
- Krejza, J.; Svetlik, J.; Bednar, P. Allometric relationship and biomass expansion factors (BEFs) for above- and below-ground biomass prediction and stem volume estimation for ash (Fraxinus excelsior L.) and oak (Quercus robur L.). Trees-Struct. Funct. 2017, 31, 1303–1316. [Google Scholar] [CrossRef]
- Wang, C.K. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. For. Ecol. Manag. 2006, 222, 9–16. [Google Scholar] [CrossRef]
- Wassenberg, M.; Chiu, H.-S.; Guo, W.; Spiecker, H. Analysis of wood density profiles of tree stems: Incorporating vertical variations to optimize wood sampling strategies for density and biomass estimations. Trees-Struct. Funct. 2015, 29, 551–561. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Zasada, M.; Bronisz, K.; Bronisz, A.; Bijak, S. Biomass conversion and expansion factors for a chronosequence of young naturally regenerated silver birch (Betula pendula Roth) stands growing on post-agricultural sites. For. Ecol. Manag. 2017, 384, 208–220. [Google Scholar] [CrossRef]
- Schroeder, P.; Brown, S.; Mo, J.; Birdsey, R.; Cieszewski, C. Biomass Estimation for Temperate Broadleaf Forests of the United States Using Inventory Data. For. Sci. 1997, 43, 424–434. [Google Scholar]
- Segura, M.; Kanninen, M. Allometric models for tree volume and total aboveground biomass in a tropical humid forest in Costa Rica. Biotropica 2005, 37, 2–8. [Google Scholar] [CrossRef]
- Goussanou, C.A.; Guendehou, S.; Assogbadjo, A.E.; Kaire, M.; Sinsin, B.; Cuni-Sanchez, A. Specific and generic stem biomass and volume models of tree species in a West African tropical semi-deciduous forest. Silva Fenn. 2016, 50, 22. [Google Scholar] [CrossRef]
- Henry, M.; Bombelli, A.; Trotta, C.; Alessandrini, A.; Birigazzi, L.; Sola, G.; Vieilledent, G.; Santenoise, P.; Longuetaud, F.; Valentini, R.; et al. GlobAllomeTree: International platform for tree allometric equations to support volume, biomass and carbon assessment. Iforest-Biogeosci. For. 2013, 6, 326–330. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, X.; Ouyang, Z.; Lu, F.; Feng, L.; Tao, J. A review of biomass equations for China’s tree species. Earth Syst. Sci. Data 2020, 12, 21–40. [Google Scholar] [CrossRef]
- Sileshi, G.W. A critical review of forest biomass estimation models, common mistakes and corrective measures. For. Ecol. Manag. 2014, 329, 237–254. [Google Scholar] [CrossRef]
- Temesgen, H.; Affleck, D.; Poudel, K.; Gray, A.; Sessions, J. A review of the challenges and opportunities in estimating above ground forest biomass using tree-level models. Scand. J. For. Res. 2015, 30, 326–335. [Google Scholar] [CrossRef]
- Zianis, D.; Muukkonen, P.; Mäkipää, R.; Mencuccini, M. Biomass and stem volume equations for tree species in Europe. Silva Fenn. Monogr. 2005, 4, 1–63. [Google Scholar]
- Jenkins, J.C.; Chojnacky, D.C.; Heath, L.S.; Birdsey, R.A. National-scale biomass estimators for United States tree species. For. Sci. 2003, 49, 12–35. [Google Scholar]
- Muukkonen, P. Generalized allometric volume and biomass equations for some tree species in Europe. Eur. J. For. Res. 2007, 126, 157–166. [Google Scholar] [CrossRef]
- Bi, H.Q.; Turner, J.; Lambert, M.J. Additive biomass equations for native eucalypt forest trees of temperate Australia. Trees-Struct. Funct. 2004, 18, 467–479. [Google Scholar] [CrossRef]
- Zhao, D.; Kane, M.; Markewitz, D.; Teskey, R.; Clutter, M. Additive Tree Biomass Equations for Midrotation Loblolly Pine Plantations. For. Sci. 2015, 61, 613–623. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, Y.; Zhang, Z.; Xie, L.; Li, F. Comparison of Tree Biomass Modeling Approaches for Larch (Larix olgensis Henry) Trees in Northeast China. Forests 2020, 11, 202. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Li, F. Additive Biomass Equations Based on Different Dendrometric Variables for Two Dominant Species (Larix gmelini Rupr. and Betula platyphylla Suk.) in Natural Forests in the Eastern Daxing’an Mountains, Northeast China. Forests 2018, 9, 261. [Google Scholar] [CrossRef]
- Parresol, B.R. Additivity of nonlinear biomass equations. Can. J. For. Res. 2001, 31, 865–878. [Google Scholar] [CrossRef]
- Tang, S.; Wang, Y. A parameter estimation program for the error-in-variable model. Ecol. Model. 2002, 156, 225–236. [Google Scholar] [CrossRef]
- Fan, S.; Liu, G.; Zhang, Q.; Feng, H.; Zong, Y.; Ren, H. A Study on Biomass and Productivity of Populus × xiaohei Plantation on Sandy Land in North China. For. Res. 2010, 23, 71–76. [Google Scholar]
- Song, B.; Li, F.; Dong, L.; Zhou, Y. Additive system of biomass equations for planted Populus simonii × P. nigra in western Heilongjiang Province of northeastern China. J. Beijing For. Univ. 2018, 40, 58–68. [Google Scholar]
- Houghton, R.A. Converting terrestrial ecosystems from sources to sinks of carbon. Ambio 1996, 25, 267–272. [Google Scholar]
- Kraenzel, M.; Castillo, A.; Moore, T.; Potvin, C. Carbon storage of harvest-age teak (Tectona grandis) plantations, Panama. For. Ecol. Manag. 2003, 173, 213–225. [Google Scholar] [CrossRef]
- Gao, B.; Taylor, A.R.; Chen, H.Y.H.; Wang, J. Variation in total and volatile carbon concentration among the major tree species of the boreal forest. For. Ecol. Manag. 2016, 375, 191–199. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, C.; Wang, X.; Quan, X. Carbon concentration variability of 10 Chinese temperate tree species. For. Ecol. Manag. 2009, 258, 722–727. [Google Scholar] [CrossRef]
- Yang, S.; Yin, X.; Sun, Y. The preparation for dual tree volume table of Populus xiaohei in eastern part of Heilongjiang Province. For. Prospect Des. 2010, 2, 59–61. [Google Scholar]
- West, P.W. Tree and Forest Measurement, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2015; p. 214. [Google Scholar]
- Affleck, D.L.R.; Diéguez-Aranda, U. Additive Nonlinear Biomass Equations: A Likelihood-Based Approach. For. Sci. 2016, 62, 129–140. [Google Scholar] [CrossRef]
- Zhao, D.; Westfall, J.; Coulston, J.W.; Lynch, T.B.; Bullock, B.P.; Montes, C.R. Additive biomass equations for slash pine trees: Comparing three modeling approaches. Can. J. For. Res. 2019, 49, 27–40. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/ETS® 15.1 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2018. [Google Scholar]
- Cunliffe, A.M.; McIntire, C.D.; Boschetti, F.; Sauer, K.J.; Litvak, M.; Anderson, K.; Brazier, R.E. Allometric Relationships for Predicting Aboveground Biomass and Sapwood Area of Oneseed Juniper (Juniperus monosperma) Trees. Front. Plant Sci. 2020, 11, 12. [Google Scholar] [CrossRef]
- Kenzo, T.; Himmapan, W.; Yoneda, R.; Tedsorn, N.; Vacharangkura, T.; Hitsuma, G.; Noda, I. General estimation models for above- and below-ground biomass of teak (Tectona grandis) plantations in Thailand. For. Ecol. Manag. 2020, 457, 117701. [Google Scholar] [CrossRef]
- Goicoa, T.; Militino, A.F.; Ugarte, M.D. Modelling aboveground tree biomass while achieving the additivity property. Environ. Ecol. Stat. 2011, 18, 367–384. [Google Scholar] [CrossRef]
- Widagdo, F.R.A.; Li, F.; Zhang, L.; Dong, L. Aggregated Biomass Model Systems and Carbon Concentration Variations for Tree Carbon Quantification of Natural Mongolian Oak in Northeast China. Forests 2020, 11, 397. [Google Scholar] [CrossRef]
- Satoo, T. Forest Biomass; Martinus Nijhoff/Dr. W. Junk: The Hague, The Netherlands, 1982. [Google Scholar]
- Gonzalez-Garcia, M.; Hevia, A.; Majada, J.; Barrio-Anta, M. Above-ground biomass estimation at tree and stand level for short rotation plantations of Eucalyptus nitens (Deane & Maiden) Maiden in Northwest Spain. Biomass Bioenergy 2013, 54, 147–157. [Google Scholar]
- Guo, Z.; Fang, J.; Pan, Y.; Birdsey, R. Inventory-based estimates of forest biomass carbon stocks in China: A comparison of three methods. For. Ecol. Manag. 2010, 259, 1225–1231. [Google Scholar] [CrossRef]
- Jalkanen, A.; Makipaa, R.; Stahl, G.; Lehtonen, A.; Petersson, H. Estimation of the biomass stock of trees in Sweden: Comparison of biomass equations and age-dependent biomass expansion factors. Ann. For. Sci. 2005, 62, 845–851. [Google Scholar] [CrossRef]
- Lehtonen, A.; Makipaa, R.; Heikkinen, J.; Sievanen, R.; Liski, J. Biomass expansion factors (BEFs) for Scots pine, Norway spruce and birch according to stand age for boreal forests. For. Ecol. Manag. 2004, 188, 211–224. [Google Scholar] [CrossRef]
- Laiho, R.; Laine, J. Tree stand biomass and carbon content in an age sequence of drained pine mires in southern Finland. For. Ecol. Manag. 1997, 93, 161–169. [Google Scholar] [CrossRef]
- Lamlom, S.H.; Savidge, R.A. A reassessment of carbon content in wood: Variation within and between 41 North American species. Biomass Bioenergy 2003, 25, 381–388. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | N | Min | Max | Mean | SD |
|---|---|---|---|---|---|
| (cm) | 128 | 2.0 | 38.0 | 14.3 | 8.5 |
| (m) | 128 | 2.5 | 25.3 | 11.8 | 5.0 |
| (m) | 128 | 0.5 | 19.7 | 7.2 | 3.8 |
| 128 | 0.3 | 4.0 | 1.5 | 0.7 | |
| (year) | 128 | 2.0 | 35.0 | 17.8 | 8.0 |
| Total biomass (kg) | 128 | 0.62 | 630.52 | 93.78 | 134.01 |
| Root biomass (kg) | 128 | 0.11 | 108.13 | 15.49 | 20.19 |
| Stem biomass (kg) | 128 | 0.34 | 420.27 | 60.74 | 90.42 |
| Branch biomass (kg) | 128 | 0.06 | 102.15 | 13.65 | 21.13 |
| Foliage biomass (kg) | 128 | 0.01 | 29.07 | 3.90 | 5.48 |
| Stem volume (m3) | 128 | 0.0009 | 0.9740 | 0.1555 | 0.2145 |
| Components | Equations | RMSE | AIC | |
|---|---|---|---|---|
| Root | 0.9361 | 5.0829 | 783.48 | |
| 0.9369 | 5.0526 | 783.94 | ||
| 0.9446 | 4.7349 | 767.32 | ||
| 0.9360 | 5.0873 | 785.70 | ||
| 0.9460 | 4.6723 | 765.91 | ||
| 0.9369 | 5.0507 | 785.85 | ||
| 0.9446 | 4.7338 | 769.26 | ||
| 0.9463 | 4.6608 | 767.28 | ||
| Stem | 0.9323 | 23.443 | 1174.82 | |
| 0.9686 | 15.9484 | 1078.20 | ||
| 0.9349 | 22.9881 | 1171.80 | ||
| 0.9448 | 21.1674 | 1150.68 | ||
| 0.9743 | 14.4471 | 1054.89 | ||
| 0.9696 | 15.7087 | 1076.33 | ||
| 0.9482 | 20.4911 | 1144.37 | ||
| 0.9753 | 14.1656 | 1051.86 | ||
| Branch | 0.8640 | 7.7596 | 891.78 | |
| 0.8672 | 7.6678 | 890.73 | ||
| 0.8711 | 7.5563 | 886.98 | ||
| 0.8678 | 7.6515 | 890.18 | ||
| 0.8732 | 7.4942 | 886.86 | ||
| 0.8672 | 7.6674 | 892.71 | ||
| 0.8732 | 7.494 | 886.86 | ||
| 0.8725 | 7.5151 | 889.58 | ||
| Foliage | 0.8346 | 2.2203 | 571.44 | |
| 0.8629 | 2.0212 | 549.39 | ||
| 0.8321 | 2.2369 | 575.35 | ||
| 0.8407 | 2.1788 | 568.62 | ||
| 0.8603 | 2.0406 | 553.84 | ||
| 0.8629 | 2.0213 | 551.41 | ||
| 0.8385 | 2.1941 | 572.41 | ||
| 0.8598 | 2.0441 | 556.27 |
| Model Systems | Components | βi0 | βi1 | βi2 | βi3 | βi4 | R2 | RMSE | ϕ | MPE | MAPE | EF |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MS1 | Root | −3.0963 ** (0.1163) | 2.0676 ** (0.0378) | 0.9363 | 5.0361 | 0.5500 | 0.38 | 29.04 | 0.9283 | |||
| Stem | −2.7171 ** (0.1324) | 2.3758 ** (0.0418) | 0.9316 | 23.3782 | 0.5040 | 2.35 | 18.28 | 0.9209 | ||||
| Branch | −4.1284 ** (0.2534) | 2.3560 ** (0.0768) | 0.8640 | 7.6981 | 0.3500 | 0.46 | 37.51 | 0.8434 | ||||
| Foliage | −4.4739 ** (0.2364) | 2.0621 ** (0.0773) | 0.8321 | 2.2194 | 0.5650 | 2.07 | 53.17 | 0.8168 | ||||
| Total | — | — | 0.9633 | 25.5756 | 1.74 | 13.86 | 0.9578 | |||||
| MS2 | Root | −2.7455 ** (0.1447) | 1.8924 ** (0.0599) | 0.2784 ** (0.0775) | 0.9442 | 4.6942 | 0.5356 | −0.01 | 27.96 | 0.9324 | ||
| Stem | −3.6308 ** (0.1238) | 1.8585 ** (0.0537) | 0.9137 ** (0.0779) | 0.9685 | 15.8047 | 0.4968 | −0.02 | 14.25 | 0.9627 | |||
| Branch | −3.8488 ** (0.3012) | 2.2030 ** (0.1144) | 0.2750 * (0.1361) | 0.8702 | 7.4923 | 0.3375 | 0.52 | 35.46 | 0.8419 | |||
| Foliage | −4.0767 ** (0.2724) | 2.4441 ** (0.1387) | −0.5718 * (0.1894) | 0.8578 | 2.0182 | 0.5503 | 1.42 | 53.92 | 0.8459 | |||
| Total | — | — | — | 0.9781 | 19.7545 | 0.12 | 10.08 | 0.9748 | ||||
| MS3 | Root | −2.7445 ** (0.1447) | 1.8919 ** (0.0599) | 0.2794 ** (0.0775) | 0.9442 | 4.6938 | 0.5356 | −0.02 | 27.95 | 0.9321 | ||
| Stem | −4.0585 ** (0.1600) | 2.0416 ** (0.0700) | 1.0536 ** (0.0955) | −0.1871 ** (0.0557) | −0.1709 * (0.0733) | 0.9741 | 14.2158 | 0.4965 | 0.04 | 12.60 | 0.9664 | |
| Branch | −3.8541 ** (0.3011) | 2.2050 ** (0.1144) | 0.2745 * (0.1359) | 0.8701 | 7.4940 | 0.3375 | 0.46 | 35.43 | 0.8413 | |||
| Foliage | −4.0972 ** (0.2722) | 2.4532 ** (0.1383) | −0.5734 * (0.1888) | 0.8603 | 2.0165 | 0.5503 | 1.09 | 53.68 | 0.8459 | |||
| Total | — | — | — | — | — | 0.9800 | 18.8546 | 0.14 | 11.10 | 0.9755 |
| Equations | β0 | β1 | β2 | β3 | β4 | R2 | RMSE | AIC | ϕ | MPE | MAPE | EF |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| −8.1001 ** (0.1434) | 2.1995 ** (0.0432) | 0.9498 | 0.0479 | −410.70 | 0.6010 | −0.63 | 26.7245 | 0.9416 | ||||
| −9.1235 ** (0.1155) | 1.7252 ** (0.0432) | 0.8949 ** (0.0629) | 0.9845 | 0.0266 | −559.46 | 0.6441 | −0.26 | 14.9243 | 0.9807 | |||
| −8.0681 ** (0.1267) | 1.8657 ** (0.0724) | 0.4289 ** (0.0782) | 0.9605 | 0.0425 | −439.56 | 0.6575 | −0.35 | 24.9401 | 0.9526 | |||
| −8.2324 ** (0.1781) | 2.2634 ** (0.0678) | −0.0955 ns (0.0806) | 0.9498 | 0.0479 | −408.73 | 0.6214 | −0.49 | 26.9817 | 0.9390 | |||
| −9.2455 ** (0.1341) | 1.7788 ** (0.0514) | 0.9813 ** (0.0838) | −0.1222 ns (0.0699) | 0.9853 | 0.0259 | −564.04 | 0.7114 | −0.18 | 14.3864 | 0.9813 | ||
| −9.3333 ** (0.1314) | 1.8184 ** (0.0516) | 0.9024 ** (0.0606) | −0.1489 ** (0.0486) | 0.9860 | 0.0253 | −570.38 | 0.6508 | −0.23 | 15.3972 | 0.9823 | ||
| −8.2102 ** (0.1577) | 1.9390 ** (0.0862) | 0.4209 ** (0.0774) | −0.0966 ns (0.0718) | 0.9609 | 0.0423 | −438.72 | 0.6926 | −0.24 | 25.1799 | 0.9523 | ||
| −9.4847 ** (0.1487) | 1.8789 ** (0.0582) | 1.0105 ** (0.0797) | −0.1409 * (0.0663) | −0.1564 ** (0.0485) | 0.9869 | 0.0245 | −576.36 | 0.6633 | −0.29 | 14.9778 | 0.9826 |
| Components | N | Min | Max | Mean | SD |
|---|---|---|---|---|---|
| Root | 128 | 28.8 | 296.7 | 118.2 | 41.1 |
| Stem | 128 | 199.1 | 616.0 | 380.2 | 67.3 |
| Branch | 128 | 28.9 | 266.7 | 90.7 | 44.6 |
| Foliage | 128 | 4.7 | 89.1 | 31.2 | 16.9 |
| Components | Min | Max | Mean | SD |
|---|---|---|---|---|
| Root | 42.31 | 50.00 | 45.98a | 2.31 |
| Stem | 41.88 | 50.65 | 47.74b | 1.84 |
| Branch | 43.29 | 51.61 | 48.32b | 1.95 |
| Foliage | 45.56 | 52.18 | 48.46b | 1.90 |
| WMCC | 44.38 | 49.64 | 47.43 | 1.51 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, L.; Widagdo, F.R.A.; Xie, L.; Li, F. Biomass and Volume Modeling along with Carbon Concentration Variations of Short-Rotation Poplar Plantations. Forests 2020, 11, 780. https://doi.org/10.3390/f11070780
Dong L, Widagdo FRA, Xie L, Li F. Biomass and Volume Modeling along with Carbon Concentration Variations of Short-Rotation Poplar Plantations. Forests. 2020; 11(7):780. https://doi.org/10.3390/f11070780
Chicago/Turabian StyleDong, Lihu, Faris Rafi Almay Widagdo, Longfei Xie, and Fengri Li. 2020. "Biomass and Volume Modeling along with Carbon Concentration Variations of Short-Rotation Poplar Plantations" Forests 11, no. 7: 780. https://doi.org/10.3390/f11070780
APA StyleDong, L., Widagdo, F. R. A., Xie, L., & Li, F. (2020). Biomass and Volume Modeling along with Carbon Concentration Variations of Short-Rotation Poplar Plantations. Forests, 11(7), 780. https://doi.org/10.3390/f11070780