Decline of European Beech in Austria: Involvement of Phytophthora spp. and Contributing Biotic and Abiotic Factors


 and
and
Abstract
:1. Introduction
2. Material and Methods
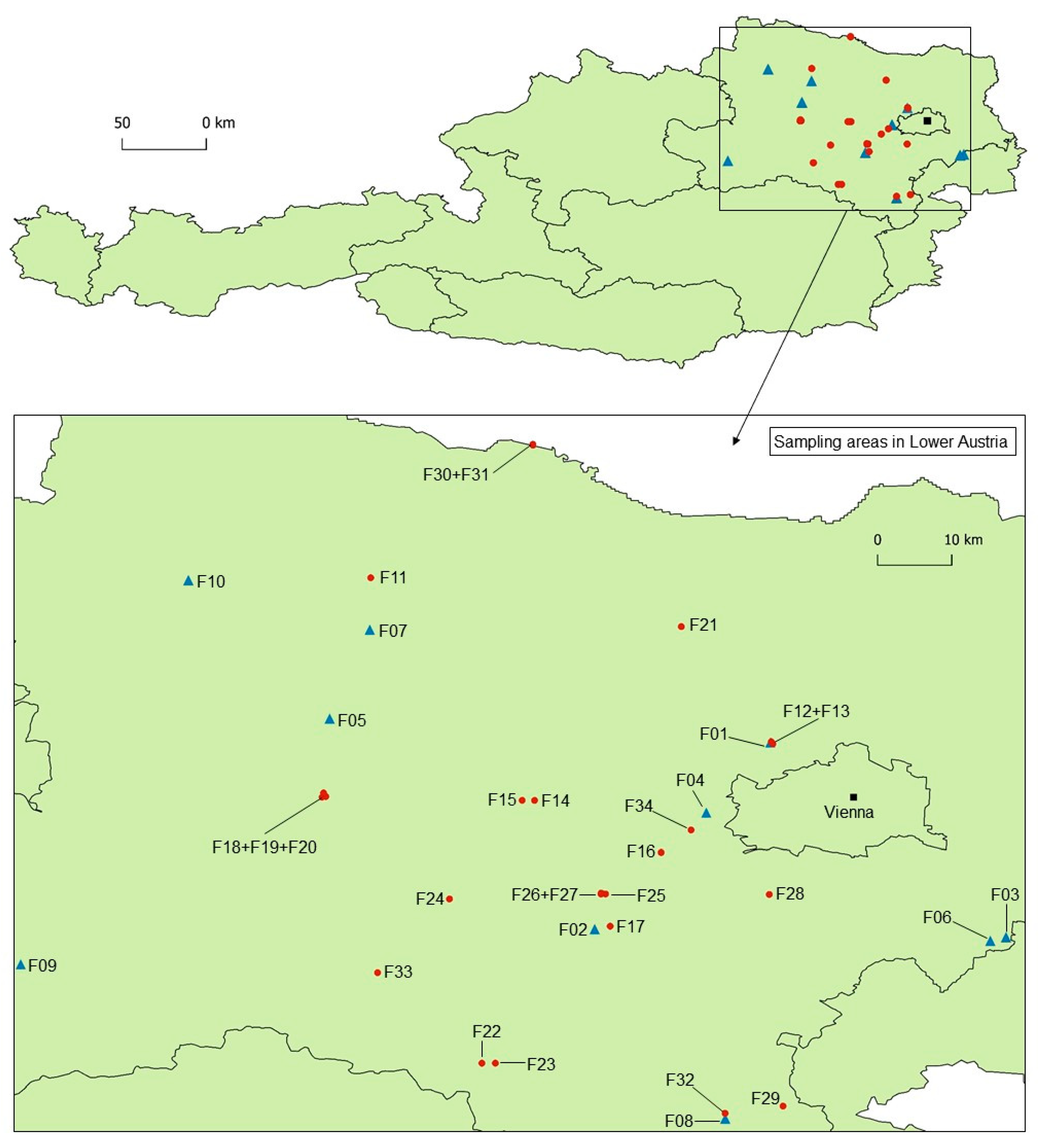
2.1. Study Sites and Field Assessments
2.2. Sampling and Isolation Procedures, Species Identification
2.2.1. Soil Sampling and Baiting
2.2.2. Isolations from Bleeding Collar Rot and Aerial Bark Lesions
2.3. Root Biometry
2.4. Ectomycorrhizal Frequency
2.5. Climate, Geological Substrate and Soil Analysis
2.6. Statistical Analysis
3. Results
3.1. Phytophthora-Related Disease Symptoms
3.2. Distribution of Phytophthora Species and Their Association with Disease Symptoms
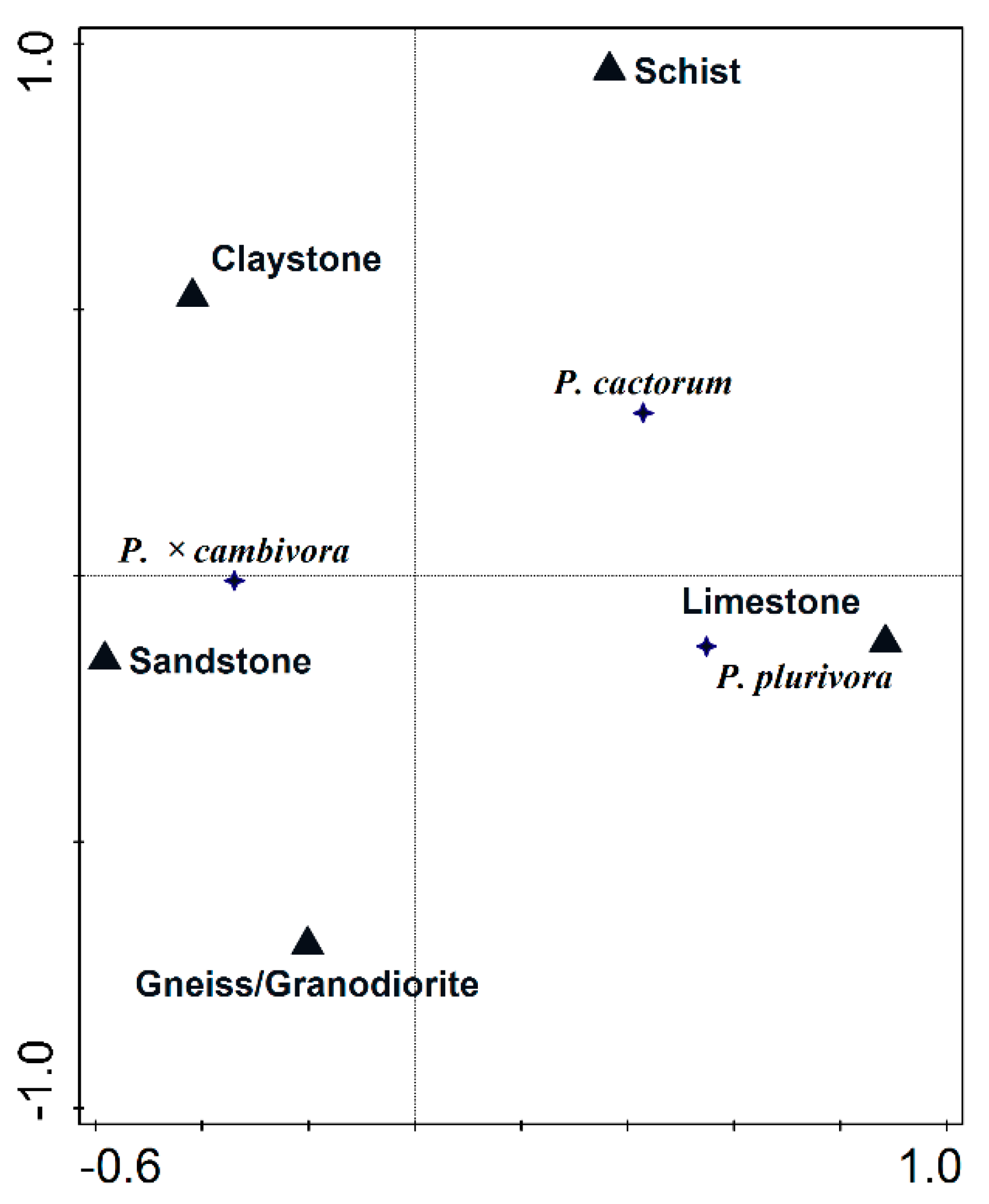
3.3. Influence of Geological Substrate, Soil Texture and Soil pH on Phytophthora Distribution
3.4. Fungal Pathogens and Pests
3.5. Relationship between Root Parameters, Ectomycorrhizal Frequency, Crown Transparency and Phytophthora Infestation in Ten Intensively Surveyed Beech Stands
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Petit, R.J.; Hampe, A. Some evolutionary consequences of being a tree. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 187–214. [Google Scholar] [CrossRef] [Green Version]
- Fritz, P. (Ed.) Ökologischer Waldumbau in Deutschland (Ecological Reconstruction of Forests in Germany); Oekom Verlag: Munich, Germany, 2006; p. 351. ISBN 978-3-86581-001-4. [Google Scholar]
- Walentowski, H.; Ewald, J.; Fischer, A.; Kölling, C.; Türk, W. Handbuch der Natürlichen Waldgesellschaften Bayerns (Handbook of the Natural Forest Types of Bavaria), 2nd ed.; Geobotanica: Freising, Germany, 2006; p. 441. ISBN 3-930560-04-6. [Google Scholar]
- Del Río, S.; Álvarez-Esteban, R.; Cano, E.; Pinto-Gomes, C.; Penas, Á. Potential impacts of climate change on habitat suitability of Fagus sylvatica L. forests in Spain. Plant Biosyst. 2018, 152, 1205–1213. [Google Scholar] [CrossRef]
- Jump, A.S.; Hunt, J.M.; Penuelas, J. Rapid climate change-related growth decline at the southern range edge of Fagus sylvatica. Glob. Chang. Biol. 2006, 12, 2163–2174. [Google Scholar] [CrossRef] [Green Version]
- Czúcz, B.; Galhidy, L.; Matyas, C. Present and forecasted xeric climatic limits of beech and sessile oak distribution at low altitudes in Central Europe. Ann. For. Sci. 2011, 68, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Gora, V.; König, J.; Lunderstädt, J. Population dynamics of beech scale (Cryptococcus fagisuga) (Coccina, Pseudococcidae) related to physiological defence reactions of attacked beech trees (Fagus sylvatica). Chemoecology 1996, 7, 112–120. [Google Scholar] [CrossRef]
- Mazzoglio, P.J.; Paoletta, M.; Patetta, A.; Currado, I. Calliteara pudibunda (Lepidoptera, Lymantriidae) in Northwest Italy. Bull. Insectology 2005, 58, 25–34. [Google Scholar]
- Lonsdale, D. Nectria infection of beech bark: Variations in disease in relation to predisposing factors. Ann. Sci. For. 1980, 37, 307–317. [Google Scholar] [CrossRef]
- Lonsdale, D.; Wainhouse, D. Beech bark disease. In Forestry Commission Bulletin; HMSO: London, UK, 1987; p. 15. ISBN 0-11-710207-5. [Google Scholar]
- Metzler, B.; Meierjohann, E.; Kublin, E.; Von Wuehlisch, G. Spatial dispersal of Nectria ditissima canker of beech in an international provenance trial. For. Pathol. 2002, 32, 137–144. [Google Scholar] [CrossRef]
- Houston, D.R. Beech bark disease: 1934 to 2004: What’s new since Ehrlich? In Beech Bark Disease, Proceedings of the Beech Bark Disease Symposium, Saranac Lake, NY, USA, 16–18 June 2004; Gen. Tech. Rep. NE-331; US. Department of Agriculture Forest Service, Northern Research Station: Newtown Square, PA, USA, 2005; pp. 2–13. Available online: https://www.nrs.fs.fed.us/pubs/7401 (accessed on 18 July 2020).
- Day, W.R. Root-rot of sweet chestnut and beech caused by species of Phytophthora. I. Cause and symptoms of disease: Its relation to soil conditions. Forestry 1938, 12, 101–116. [Google Scholar] [CrossRef]
- Day, W.R. Root-rot of sweet chestnut and beech caused by species of Phytophthora. II. Inoculation experiments and methods of control. Forestry 1939, 13, 46–58. [Google Scholar] [CrossRef]
- Jung, T.; Blaschke, H. Phytophthora root rot in declining forest trees. Phyton (Horn, Austria) 1996, 36, 95–102. [Google Scholar]
- Balci, Y.; Halmschlager, E. Phytophthora species in oak ecosystems in Turkey and their association with declining oak trees. Plant Pathol. 2003, 52, 694–702. [Google Scholar] [CrossRef]
- Motta, E.; Annesi, T.; Pane, A.; Cooke, D.E.L.; Cacciola, S.O. A new Phytophthora sp. causing a basal canker on beech in Italy. Plant Dis. 2003, 87, 1005. [Google Scholar] [CrossRef] [PubMed]
- Brasier, C.M.; Beales, P.A.; Denman, S.; Rose, J. Phytophthora kernoviae sp. nov., an invasive pathogen causing bleeding stem lesions on forest trees and foliar necrosis of ornamentals in the UK. Mycol. Res. 2005, 109, 853–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, T.; Hudler, G.W.; Jensen-Tracy, S.L.; Griffiths, H.M.; Fleischmann, F.; Oßwald, W. Involvement of Phytophthora spp. in the decline of European beech in Europe and the USA. Mycologist 2005, 19, 159–166. [Google Scholar] [CrossRef]
- Jung, T.; Vettraino, A.M.; Cech, T.L.; Vannini, A. The impact of invasive Phytophthora species on European forests. In Phytophthora: A Global Perspective; Lamour, K., Ed.; CABI: Wallingford, UK, 2013; pp. 146–158. ISBN 978-1-78064-093-8. [Google Scholar]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Aguín Casal, O.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; Chavarriaga, D.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Pathol. 2016, 46, 134–163. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Pérez–Sierra, A.; Durán, A.; Horta Jung, M.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil- and airborne Phytophthora species in forests and woodlands. Persoonia 2018, 40, 182–220. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; La Spada, F.; Pane, A.; Aloi, F.; Evoli, M.; Horta Jung, M.; Scanu, B.; Faedda, R.; Rizza, C.; Puglisi, I.; et al. Diversity and distribution of Phytophthora species in protected natural areas in Sicily. Forests 2019, 10, 259. [Google Scholar] [CrossRef] [Green Version]
- Cech, T.L.; Jung, T. Phytophthora—Wurzelhalsfäulen an Buchen nehmen auch in Österreich zu (Phytophthora root rot of beech is also increasing in Austria). Forstsch. Aktuell 2005, 34, 7. [Google Scholar]
- Hartmann, G.; Blank, R.; Kunca, A. Collar rot of Fagus sylvatica caused by Phytophthora cambivora: Damage, site relations and susceptibility of broadleaf hosts. In Progress in Research on Phytophthora Diseases of Forest Trees, Proceedings of the 3rd International IUFRO Working Party 7.02.09 Meeting, Freising, Germany, 11–17 September 2004; Brasier, C., Jung, T., Osswald, W., Eds.; Forest Research: Farnham, UK, 2006; pp. 135–138. ISBN 0-85538-721-1. [Google Scholar]
- Brown, A.V.; Brasier, C.M. Colonization of tree xylem by Phytophthora ramorum, P. kernoviae and other Phytophthora species. Plant Pathol. 2007, 56, 227–241. [Google Scholar] [CrossRef]
- Munda, A.; Zerjav, M.; Schroers, H.J. First report of Phytophthora citricola occurring on Fagus sylvatica in Slovenia. Plant Dis. 2007, 91, 907. [Google Scholar] [CrossRef] [PubMed]
- Vettraino, A.M.; Jung, T.; Vannini, A. First report of Phytophthora cactorum associated with beech decline in Italy. Plant Dis. 2008, 92, 1708. [Google Scholar] [CrossRef] [PubMed]
- Cerny, K.; Strnadova, V.; Gregorova, B.; Holub, V.; Tomsovsky, M.; Mrazkova, M.; Gabrielova, S. Phytophthora cactorum causing bleeding canker of common beech, horse chestnut, and white poplar in the Czech Republic. Plant Pathol. 2009, 58, 394. [Google Scholar] [CrossRef]
- Jung, T. Beech decline in Central Europe driven by the interaction between Phytophthora infections and climatic extremes. For. Pathol. 2009, 39, 73–94. [Google Scholar] [CrossRef]
- Schmitz, S.; Zini, J.; Chandelier, A. Involvement of Phytophthora species in the decline of beech (Fagus sylvatica) in the southern part of Belgium. In Phytophthoras in Forests and Natural Ecosystems: Fourth Meeting of the International Union of Forest Research Organizations (IUFRO) Working Party S07.02.09; General Technical Report PSW-GTR-221; Goheen, E.M., Frankel, S.J., Eds.; USDA Forest Service Pacific Southwest Research Station: Albany, CA, USA, 2009; pp. 320–323. [Google Scholar]
- Stępniewska, H.; Dłuszyński, J. Incidence of Phytophthora cambivora in bleeding lesions on beech stems in selected forest stands in South-eastern Poland. Phytopathologia 2010, 56, 39–51. [Google Scholar]
- Weiland, G.E.; Nelson, A.H.; Hudler, G.W. Aggressiveness of Phytophthora cactorum, P. citricola I and P. plurivora from European beech. Plant Dis. 2010, 94, 1009–1014. [Google Scholar] [CrossRef] [Green Version]
- Nechwatal, J.; Hahn, J.; Schönborn, A.; Schmitz, G. A twig blight of understorey European beech (Fagus sylvatica) caused by soilborne Phytophthora spp. For. Pathol. 2011, 41, 493–500. [Google Scholar] [CrossRef]
- Milenković, I.; Keča, N.; Karadžić, D.; Nowakowska, J.A.; Borys, M.; Sikora, K.; Oszako, T. Incidence of Phytophthora species in beech stands in Serbia. Folia For. Pol. 2012, 54, 223–232. [Google Scholar]
- Cacciola, S.O.; Motta, E.; Raudino, F.; Chimento, A.; Pane, A.; Magnano di San Lio, G. Phytophthora pseudosyringae the causal agent of bleeding cankers of beech in central Italy. J. Plant Pathol. 2005, 87, 289. [Google Scholar]
- Telfer, K.H.; Brurberg, M.B.; Herrero, M.L.; Stensvand, A.; Talgø, V. Phytophthora cambivora found on beech in Norway. For. Pathol. 2015, 45, 415–425. [Google Scholar] [CrossRef]
- Cleary, M.; Blomquist, M.; Ghasemkhani, M.; Witzell, J. First report of Phytophthora gonapodyides causing stem canker on European beech (Fagus sylvatica) in southern Sweden. Plant Dis. 2016, 100, 2174. [Google Scholar] [CrossRef]
- Jung, T.; Horta Jung, M.; Cacciola, S.O.; Cech, T.; Bakonyi, J.; Seress, D.; Mosca, S.; Schena, L.; Seddaiu, S.; Pane, A.; et al. Multiple new cryptic pathogenic Phytophthora species from Fagaceae forests in Austria, Italy and Portugal. IMA Fungus 2017, 8, 219–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink, A.H.; Brücher, T.; Krüger, A.; Leckebusch, G.C.; Pinto, J.G.; Ulbrich, U. The 2003 European summer heatwaves and drought–synoptic diagnosis and impacts. Weather 2004, 59, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Brasier, C.M.; Scott, J.K. European oak declines and global warming: A theoretical assessment with special reference to the activity of Phytophthora cinnamomi. EPPO Bull. 1994, 24, 221–234. [Google Scholar] [CrossRef]
- Brasier, C.M. Phytophthora cinnamomi and oak decline in southern Europe. Environmental constraints including climate change. Ann. Sci. For. 1996, 53, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Brasier, C.M. The biosecurity threat to the UK and global environment from international trade in plants. Plant Pathol. 2008, 57, 792–808. [Google Scholar] [CrossRef]
- Jung, T.; Blaschke, H.; Neumann, P. Isolation, identification and pathogenicity of Phytophthora species from declining oak stands. Eur. J. For. Pathol. 1996, 26, 253–272. [Google Scholar] [CrossRef]
- Jung, T.; Blaschke, H.; Oßwald, W. Involvement of soilborne Phytophthora species in Central European oak decline and the effect of site factors on the disease. Plant Pathol. 2000, 49, 706–718. [Google Scholar] [CrossRef]
- Anonymous. Climate Change 2014—Synthesis Report: Impacts, Adaptation, and Vulnerability. In Proceedings of the Intergovernmental Panel on Climate Change (IPCC), Geneva, Switzerland; 2015. Available online: https://www.ipcc.ch/report/ar4/wg2/ (accessed on 18 July 2020).
- Burgess, T.I.; Scott, J.K.; McDougall, K.L.; Stukely, M.J.C.; Crane, C.; Dunstan, W.A.; Brigg, F.; Andjic, V.; White, D.; Rudman, T.; et al. Current and projected global distribution of Phytophthora cinnamomi, one of the world’s worst plant pathogens. Glob. Chang. Biol. 2017, 23, 1661–1674. [Google Scholar] [CrossRef] [Green Version]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; APS Press, American Phytopathological Society: St. Paul, MI, USA, 1996; p. 592. ISBN 0-89054-212-0. [Google Scholar]
- Fields, K.J.; Pressel, S. Unity in diversity: Structural and functional insights into the ancient partnerships between plants and fungi. New Phytol. 2018, 220, 996–1006. [Google Scholar] [CrossRef]
- Corcobado, T.; Vivas, M.; Moreno, G.; Solla, A. Ectomycorrhizal symbiosis in declining and non-declining Quercus ilex trees infected with or free of Phytophthora cinnamomi. For. Ecol. Manag. 2014, 324, 72–80. [Google Scholar] [CrossRef]
- Sapsford, S.J.; Paap, T.; Hardy, G.E.S.J.; Burgess, T.I. The ‘chicken or the egg’: Which comes first, forest tree decline or loss of mycorrhizae? Plant Ecol. 2017, 218, 1093–1106. [Google Scholar] [CrossRef]
- Marx, D.H. Ectomycorrhizae as biological deterrents to pathogenic root infections. Annu. Rev. Phytopathol. 1972, 10, 429–454. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Forest Condition in Europe. Results of the 1993 Survey. Convention on Long-Range Transboundary Air Pollution. In International Co-Operative Programme on Assessment and Monitoring of Air Pollution Effects on Forests; EC-UN/ECE: Brussels, Geneva, 1994. [Google Scholar]
- Jung, T.; Blaschke, M. Phytophthora root and collar rot of alders in Bavaria: Distribution, modes of spread and possible management strategies. Plant Pathol. 2004, 53, 197–208. [Google Scholar] [CrossRef]
- Scanu, B.; Hunter, G.C.; Linaldeddu, B.T.; Franceschini, A.; Maddau, L.; Jung, T.; Denman, S. A taxonomic re-evaluation reveals that Phytophthora cinnamomi and P. cinnamomi var. parvispora are separate species. For. Pathol. 2014, 44, 1–20. [Google Scholar] [CrossRef]
- Jung, T.; Hansen, E.M.; Winton, L.; Oßwald, W.; Delatour, C. Three new species of Phytophthora from European oak forests. Mycol. Res. 2002, 106, 397–411. [Google Scholar] [CrossRef]
- Jung, T.; Nechwatal, J.; Cooke, D.E.L.; Hartmann, G.; Blaschke, M.; Oßwald, W.F.; Duncan, J.M.; Delatour, C. Phytophthora pseudosyringae sp. nov., a new species causing root and collar rot of deciduous tree species in Europe. Mycol. Res. 2003, 107, 772–789. [Google Scholar] [CrossRef]
- Jung, T.; Horta Jung, M.; Scanu, B.; Seress, D.; Kovács, D.M.; Maia, C.; Pérez-Sierra, A.; Chang, T.T.; Chandelier, A.; Heungens, A.; et al. Six new Phytophthora species from ITS Clade 7a including two sexually functional heterothallic hybrid species detected in natural ecosystems in Taiwan. Persoonia 2017, 38, 100–135. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Burgess, T.I. Re-evaluation of Phytophthora citricola isolates from multiple woody hosts in Europe and North America reveals a new species, Phytophthora plurivora sp. nov. Persoonia 2009, 22, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Cooke, D.E.L.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A molecular phylogeny of Phytophthora and related oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef]
- Jung, T.; Chang, T.T.; Bakonyi, J.; Seress, D.; Pérez-Sierra, A.; Yang, X.; Hong, C.; Scanu, B.; Fu, C.H.; Hsueh, K.L.; et al. Diversity of Phytophthora species in natural ecosystems of Taiwan and association with disease symptoms. Plant Pathol. 2017, 66, 194–211. [Google Scholar] [CrossRef]
- Agerer, R. (Ed.) Colour Atlas of Ectomycorrhizae; Einhorn Verlag: Schwäbisch Gmünd, Germany, 1987; Volume 1, p. 58. ISBN 0-89054-212-0. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, 5th ed.; Microcomputer Power: Ithaca, NY, USA, 2012; p. 496. Available online: http://www.canoco5.com/ (accessed on 18 July 2020).
- Schoebel, C.N.; Stewart, J.; Gruenwald, N.J.; Rigling, D.; Prospero, S. Population history and pathways of spread of the plant pathogen Phytophthora plurivora. PLoS ONE 2014, 9, e85368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vettraino, A.M.; Brasier, C.M.; Brown, A.V.; Vannini, A. Phytophthora himalsilva sp. nov. an unusually phenotypically variable species from a remote forest in Nepal. Fungal Biol. 2011, 115, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Huai, W.X.; Tian, G.; Hansen, E.M.; Zhao, W.X.; Goheen, E.M.; Grünwald, N.J.; Cheng, C. Identification of Phytophthora species baited and isolated from forest soil and streams in northwestern Yunnan province, China. For. Pathol. 2013, 43, 87–103. [Google Scholar] [CrossRef]
- Jung, T.; Scanu, B.; Brasier, C.M.; Webber, J.; Milenković, I.; Corcobado, T.; Tomšovský, T.; Pánek, M.; Bakonyi, J.; Maia, C.; et al. A survey in natural forest ecosystems of Vietnam reveals high diversity of both new and described Phytophthora taxa including P. ramorum. Forests 2020, 11, 93. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Cooke, D.E.L.; Blaschke, H.; Duncan, J.M.; Oßwald, W. Phytophthora quercina sp. nov., causing root rot of European oaks. Mycol. Res. 1999, 103, 785–798. [Google Scholar] [CrossRef]
- Balci, Y.; Halmschlager, E. Incidence of Phytophthora species in oak forests in Austria and their possible involvement in oak decline. For. Pathol. 2003, 33, 157–174. [Google Scholar] [CrossRef]
- Pérez–Sierra, A.; López–García, C.; León, M.; García-Jiménez, J.; Abad-Campos, P.; Jung, T. Previously unrecorded low temperature Phytophthora species associated with Quercus decline in a Mediterranean forest in Eastern Spain. For. Pathol. 2013, 43, 331–339. [Google Scholar] [CrossRef]
- Jung, T.; Durán, A.; Sanfuentes von Stowasser, E.; Schena, L.; Mosca, S.; Fajardo, S.; González, M.; Navarro Ortega, A.D.; Bakonyi, J.; Seress, D.; et al. Diversity of Phytophthora species in Valdivian rainforests and association with severe dieback symptoms. For. Pathol. 2018, 48, e12443. [Google Scholar] [CrossRef]
- Lotter, A.F.; Birks, H.J.B.; Hofmann, W.; Marcetto, A. Modern diatom, cladocera, chironomid, and chrysophyte cyst assemblages as quantitative indicators for the reconstruction of past environmental conditions in the Alps. I. Climate. J. Paleolimnol. 1997, 18, 395–420. [Google Scholar] [CrossRef] [Green Version]
- Shearer, B.L.; Tippett, J.T. Jarrah Dieback: The Dynamics and Management of Phytophthora Cinnamomi in the Jarrah (Eucalyptus Marginata) Forests of South-Western Australia; Department of Conservation and Land Management: Perth, Australia, 1989; p. 76. ISSN 1032-8106.
- Hansen, E.M.; Goheen, D.J.; Jules, E.S.; Ullian, B. Managing Port–Orford–Cedar and the introduced pathogen Phytophthora lateralis. Plant Dis. 2000, 84, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vettraino, A.M.; Barzanti, G.P.; Bianco, M.C.; Ragazzi, A.; Capretti, P.; Paoletti, E.; Luisi, N.; Anselmi, N.; Vannini, A. Occurrence of Phytophthora species in oak stands in Italy and their association with declining oak trees. For. Pathol. 2002, 32, 19–28. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Morel, O.; Perlerou, C.; Robin, C.; Diamandis, S.; Vannini, A. Occurrence and distribution of Phytophthora species associated with ink disease of chestnut in Europe. Eur. J. Plant Pathol. 2005, 111, 169–180. [Google Scholar] [CrossRef]
- Brasier, C.M.; Jung, T. Recent developments in Phytophthora diseases of trees and natural ecosystems in Europe. In Progress in Research on Phytophthora Diseases of Forest Trees, Proceedings of the 3rd International IUFRO Working Party 7.02.09 Meeting, Freising, Germany, 11–17 September 2004; Brasier, C., Jung, T., Osswald, W., Eds.; Forest Research: Farnham, Surrey, UK, 2006; pp. 5–16. ISBN 0-85538-721-1. [Google Scholar]
- Milenković, I.; Keča, N.; Karadžić, D.; Radulović, Z.; Nowakowska, J.A.; Oszako, T.; Sikora, K.; Corcobado, T.; Jung, T. Isolation and pathogenicity of Phytophthora species from poplar plantations in Serbia. Forests 2018, 9, 330. [Google Scholar] [CrossRef] [Green Version]
- Fleischmann, F.; Schneider, D.; Matyssek, R.; Oßwald, W.F. Investigations on Net CO2 assimilation, transpiration and root growth of Fagus sylvatica infested with four different Phytophthora species. Plant Biol. 2002, 4, 144–152. [Google Scholar] [CrossRef]
- Brasier, C.M.; Jung, T. Progress in understanding Phytophthora diseases of trees in Europe. In Phytophthora in Forests and Natural Ecosystems, Proceedings of the 2nd International IUFRO Working Party 7.02.09 Meeting, Albany, Western Australia, 30 September–5 October 2001; McComb, J.A., Hardy, G.E.S.J., Eds.; Murdoch University Print: Perth, Australia, 2003; pp. 4–18. ISBN 0-86905-825-5. [Google Scholar]
- Cleary, M.; Blomquist, M.; Vetukuri, R.R.; Böhlenius, H.; Witzell, J. Susceptibility of common tree species in Sweden to Phytophthora cambivora, P. plurivora and P. cactorum. For. Pathol. 2017, 47, e12329. [Google Scholar] [CrossRef]
- Jung, T.; Nechwatal, J. Phytophthora gallica sp. nov., a new species from rhizosphere soil of declining oak and reed stands in France and Germany. Mycol. Res. 2008, 112, 1195–1205. [Google Scholar] [CrossRef]
- Orlikowski, L.B.; Ptaszek, M.; Rodziewicz, A.; Nechwatal, J.; Thinggaard, K.; Jung, T. Phytophthora root and collar rot of mature Fraxinus excelsior in forest stands in Poland and Denmark. For. Pathol. 2011, 41, 510–519. [Google Scholar] [CrossRef]
- Milenković, I.; Keča, N.; Karadžić, D.; Radulović, Z.; Tomšovský, M.; Jung, T. Occurrence and pathogenicity of Phytophthora ×cambivora on Prunus laurocerasus in Serbia. For. Pathol. 2018, 48, e12436. [Google Scholar] [CrossRef]
- Eamus, D.; Chen, X.; Kelley, G.; Hutley, L.B. Root biomass and root fractal analyses of an open Eucalyptus forest in a savanna of north Australia. Aust. J. Bot. 2002, 50, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Meng, S.; Jia, Q.; Zhou, G.; Zhou, H.; Liu, Q.; Yu, J. Fine Root Biomass and Its Relationship with aboveground traits of Larix gmeliniit trees in Northeastern China. Forests 2018, 9, 35. [Google Scholar] [CrossRef] [Green Version]
- Ellenberg, H. Vegetation Mitteleuropas Mit den Alpen (Vegetation of Central Europe and the Alps), 4th ed.; Eugen Ulmer: Stuttgart, Germany, 1986; p. 989. ISBN 3800134306. [Google Scholar]
- Friedrichs, D.A.; Trouet, V.; Büntgen, U.; Frank, D.C.; Esper, J.; Neuwirth, B.; Löffler, J. Species-specific climate sensitivity of tree growth in Central-West Germany. Trees 2009, 23, 729–739. [Google Scholar] [CrossRef]
- Scharnweber, T.; Manthey, M.; Criegee, C.; Bauwe, A.; Schröder, C.; Wilmking, M. Drought matters—Declining precipitation influences growth of Fagus sylvatica L. and Quercus robur L. in north-eastern Germany. For. Ecol. Manag. 2011, 262, 947–961. [Google Scholar] [CrossRef]
- El-Hamalawi, Z.A.; Menge, J.A. The role of snails and ants in transmitting the avocado stem canker pathogen, Phytophthora citricola. J. Am. Soc. Hortic. Sci. 1996, 121, 973–977. [Google Scholar] [CrossRef] [Green Version]
- Lakatos, F.; Molnár, M. Mass mortality of beech (Fagus sylvatica L.) in South-West Hungary. Acta Silv. Lignaria Hung. 2009, 5, 75–82. [Google Scholar]
- Montecchio, L.; Causin, R.; Rossi, S.; Mutto Accordi, S. Changes in ectomycorrhizal diversity in a declining Quercus ilex coastal forest. Phytopathol. Mediterr. 2004, 43, 26–34. [Google Scholar] [CrossRef]
- Bloom, J.M.; Vannini, A.; Vettraino, A.M.; Hale, M.D.; Godbold, D.L. Ectomycorrhizal community structure in a healthy and a Phytophthora-infected chestnut (Castanea sativa Mill.) stand in central Italy. Mycorrhiza 2008, 20, 25–38. [Google Scholar] [CrossRef]
- Luoma, D.L.; Eberhard, J.C. Relationships between Swiss needle cast and ectomycorrhizal fungus diversity. Mycologia 2014, 106, 666–675. [Google Scholar] [CrossRef] [Green Version]
- Spinoni, J.; Vogt, J.V.; Barbosa, P.; Dosio, A.; McCormick, N.; Bigano, A.; Füssel, H.-M. Changes of heating and cooling degree-days in Europe from 1981 to 2100. Int. J. Climatol. 2018, 38, e191–e208. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Stand | Altitude (m. a.s.l.) | Geological Substrate | Soil Texture | pH (CaCl2) | P | N | Corg | Phytophthora spp. Isolated from | ||
|---|---|---|---|---|---|---|---|---|---|---|
| no. | Name | Rhizosphere Soil (no. of Trees) a | Collar Rot and Aerial Cankers (no. of Trees) a | |||||||
| F01 | Hadersfeld 1 | 351 | Sandstone | Sandy loam | 3.9 | 0.3 | 1.1 | 13.0 | CAM (1) b,c | |
| F02 | Hocheck 1 | 992 | Limestone | Sandy loam | 7.4 | 1.7 | 9.1 | 113.0 | ||
| F03 | Kaiserstein-bruch | 237 | Limestone | Clay | 7.2 | 0.3 | 2.8 | 39.5 | PLU (1), SYR (1) b | PLU (1) d |
| F04 | Purkersdorf | 406 | Claystone and sandstone | Loamy sand | 4.2 | 0.3 | 1 | 13.8 | CAM (2) b | |
| F05 | Rossatz | 614 | Gneiss | Clayey sand | 3.7 | 0.3 | 1.1 | 22.8 | PLU (1) b | |
| F06 | Sommerein | 308 | Schist | Loamy silt | 4.4 | 0.2 | 1.3 | 20.0 | CAC (2), PSY (1) bc | CAC (1) e |
| F07 | Stackelberg | 527 | Gneiss | Clayey sand | 3.8 | 1.0 | 1.0 | 17.0 | CAM (1) b,c | |
| F08 | Thernberg 2 | 560 | Limestone | Sandy silt | 7.5 | 0.6 | 3.6 | 57.5 | PLU (1) | PLU (1) e |
| F09 | Ybbsitz | 1061 | Limestone | Clayey loam | 7.2 | 0.9 | 5.9 | 82.3 | b | |
| F10 | Zwettl | 519 | Granodiorite | Clayey sand | 4.2 | 2.7 | 3.2 | 36.0 | CAM (2) b | CAM (1) e |
| Forest Stand | Altitude (m a.s.l.) | Geological Substrate | Phytophthora spp. Isolated from | ||
|---|---|---|---|---|---|
| no. | Name | Rhizosphere Soil (no. of Trees) a | Collar rot and Aerial Cankers (no. of Trees) a | ||
| F11 | Fuglau | 420 | Schist | n.a. | CAM (1) b |
| F12 | Hadersfeld 2 | 380 | Sandstone | n.a. | CAM (4) b |
| F13 | Hadersfeld 3 | 313 | Sandstone | n.a. | n.a. |
| F14 | Haspelwald 1 | 363 | Claystone | CAM (1) | n.a. |
| F15 | Haspelwald 2 | 353 | Claystone | n.a. | CAM (1) b |
| F16 | Hengstlberg | 530 | Claystone | n.a. | CAM (6) b,c |
| F17 | Hocheck 2 | 630 | Limestone | n.a. | PLU (2) b,d, negative (1) b |
| F18 | Hohenegg 1 | 390 | Gneiss | n.a. | CAM (2) b |
| F19 | Hohenegg 2 | 397 | Gneiss | CAM (1), GON (1), PLU (1), TUB (1) | n.a. |
| F20 | Hohenegg 3 | 481 | Gneiss | n.a. | n.a. |
| F21 | Hollabrunn | 309 | Alluvial deposits | n.a. | n.a. |
| F22 | Höllental-Schwarza | 562 | Limestone | CAC (1) e | n.a. |
| F23 | Höllental-Weichtalklamm | 968 | Limestone | n.a. | n.a. |
| F24 | Kerschenbach | 434 | Claystone and sandstone | CAM (1) e | negative (1) b |
| F25 | Kleinmariazell 1 | 531 | Claystone | n.a. | n.a. |
| F26 | Kleinmariazell 2 | 581 | Claystone | n.a. | n.a. |
| F27 | Kleinmariazell 3 | 637 | Claystone | n.a. | CAM (1), negative (1) b |
| F28 | Mödling Richardhof | 457 | Limestone | PLU (2) f | n.a. |
| F29 | Rosalia | Gneiss | n.a. | CAM (1) b | |
| F30 | Thayatal 1 | 360 | Limestone | CAC (1), PLU (1) e | n.a. |
| F31 | Thayatal 2 | 330 | Gneiss | CAM (1) | n.a. |
| F32 | Thernberg 1 | 394 | Schist | n.a. | PLU (1) b |
| F33 | Türnitz | 1255 | Limestone | n.a. | n.a. |
| F34 | Wienerwaldsee | 338 | Claystone | CAC (1) | CAM (1) |
| Forest Stands | Phytophthora spp. a | Crown Transparency (%) | FRL/CRLrel b | FRS/CRSrel b | FRT/CRLrel b | FRT/CRSrel b | MT/NMTrel b | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 8 Phytophthora-Infested Stands (16 Healthy and 16 Declining Beech Trees) | |||||||||||||
| H | D | H | D | H | D | H | D | H | D | H | D | ||
| F01 Hadersfeld 1 | CAM | 10.0 | 37.5 | 100.0 | 118.1 | 100.0 | 142.9 | 100.0 | 111.7 | 100.0 | 129.0 | 100.0 | 26.0 |
| F03 Kaisersteinbruch 1 | PLU, SYR | 10.0 | 50.0 | 100.0 | 112.1 | 100.0 | 97.5 | 100.0 | 153.1 | 100.0 | 147.0 | 100.0 | 19.7 |
| F04 Purkersdorf | CAM | 10.0 | 42.5 | 100.0 | 91.6 | 100.0 | 98.4 | 100.0 | 70.0 | 100.0 | 69.7 | 100.0 | 43.5 |
| F05 Rossatz | PLU | 10.0 | 62.5 | 100.0 | 116.1 | 100.0 | 132.2 | 100.0 | 113.3 | 100.0 | 119.8 | 100.0 | 52.5 |
| F06 Sommerein | CAC, PSY | 12.5 | 47.5 | 100.0 | 96.4 | 100.0 | 88.1 | 100.0 | 134.6 | 100.0 | 122.1 | 100.0 | 79.1 |
| F07 Stackelberg | CAM | 10.0 | 30.0 | 100.0 | 30.5 | 100.0 | 35.4 | 100.0 | 30.2 | 100.0 | 28.1 | 100.0 | 58.6 |
| F08 Thernberg 2 | PLU | 10.0 | 37.5 | 100.0 | 89.7 | 100.0 | 94.4 | 100.0 | 89.3 | 100.0 | 88.2 | 100.0 | 24.8 |
| F10 Zwettl | CAM | 12.5 | 32.5 | 100.0 | 38.7 | 100.0 | 36.2 | 100.0 | 35.3 | 100.0 | 32.2 | 100.0 | 37.7 |
| Mean of 8 infested stands | 10.6 | 34.0 | 100.0 | 86.6 | 100.0 | 90.6 | 100.0 | 92.2 | 100.0 | 92.0 | 100.0 | 42.7 | |
| Significance c | *** | * | n.s. | n.s. | n.s. | ||||||||
| 2 Phytophthora-Free Stands (4 Healthy and 4 Declining Beech Trees) | |||||||||||||
| H | D | H | D | H | D | H | D | H | D | H | D | ||
| F02 Hocheck 1 | - | 10.0 | 55.0 | 100.0 | 78.6 | 100.0 | 71.3 | 100.0 | 85.6 | 100.0 | 82.3 | 100.0 | 23.2 |
| F09 Ybbsitz | - | 5.0 | 37.5 | 100.0 | 77.8 | 100.0 | 82.3 | 100.0 | 71.6 | 100.0 | 63.6 | 100.0 | 48.1 |
| Mean of 2 non-infested stands | 7.5 | 46.3 | 100.0 | 78.2 | 100.0 | 76.8 | 100.0 | 78.6 | 100.0 | 72.9 | 100.0 | 35.7 | |
| Significance c | *** | n.s. | n.s. | n.s. | n.s. | n.s. | |||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corcobado, T.; Cech, T.L.; Brandstetter, M.; Daxer, A.; Hüttler, C.; Kudláček, T.; Horta Jung, M.; Jung, T. Decline of European Beech in Austria: Involvement of Phytophthora spp. and Contributing Biotic and Abiotic Factors. Forests 2020, 11, 895. https://doi.org/10.3390/f11080895
Corcobado T, Cech TL, Brandstetter M, Daxer A, Hüttler C, Kudláček T, Horta Jung M, Jung T. Decline of European Beech in Austria: Involvement of Phytophthora spp. and Contributing Biotic and Abiotic Factors. Forests. 2020; 11(8):895. https://doi.org/10.3390/f11080895
Chicago/Turabian StyleCorcobado, Tamara, Thomas L. Cech, Martin Brandstetter, Andreas Daxer, Christine Hüttler, Tomáš Kudláček, Marília Horta Jung, and Thomas Jung. 2020. "Decline of European Beech in Austria: Involvement of Phytophthora spp. and Contributing Biotic and Abiotic Factors" Forests 11, no. 8: 895. https://doi.org/10.3390/f11080895