
Resource Partitioning of Scots Pine (Pinus sylvestris L.) by Pine Shoot Beetles in Stands under Stress Conditions


Abstract
:1. Introduction
2. Material and Methods
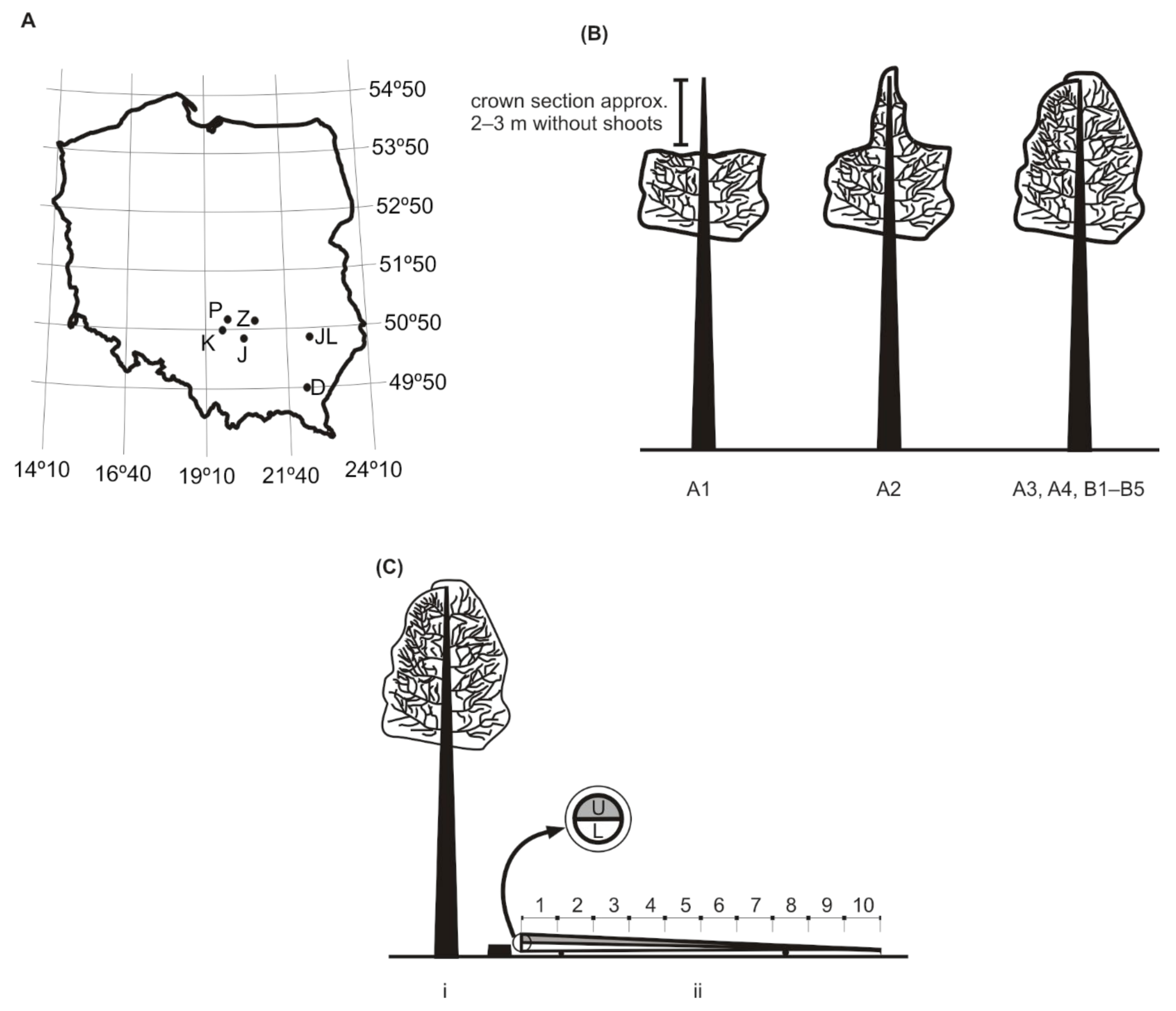
2.1. Study Site
- i.
- stand A1: crowns stunted, no shoots on a top section of ca. 2–3 m; stand located up to 100 m from the timber yard;
- ii.
- stand A2: crown in the shape of a cone in the phase of regeneration (reconstructed leader shoot); stand located 100–300 m from the timber yard;
- iii.
- stand A3: crown properly developed; stand located over 300 m from the timber yard.
2.2. Entomological Analysis
2.3. Pine Stand Quality
2.4. Evaluation of Biotic Interactions
2.5. T. piniperda Niche Model
- 1.
- habitat characteristics—site quality classes (Ia, I, II, III, IV and V), forest site type;
- 2.
- tree characteristics—degree of damage to crown (crown damaged or undamaged);
- 3.
- trap log characteristics—diameter in bark at the thicker end, length, stem surface area;
- 4.
- parameters of trap log colonisation—total colonisation density, range of colonisation of stems (1, 2, …, 10 correspond respectively to 10%, 20%, …, 100% of the length of the stem).
3. Results
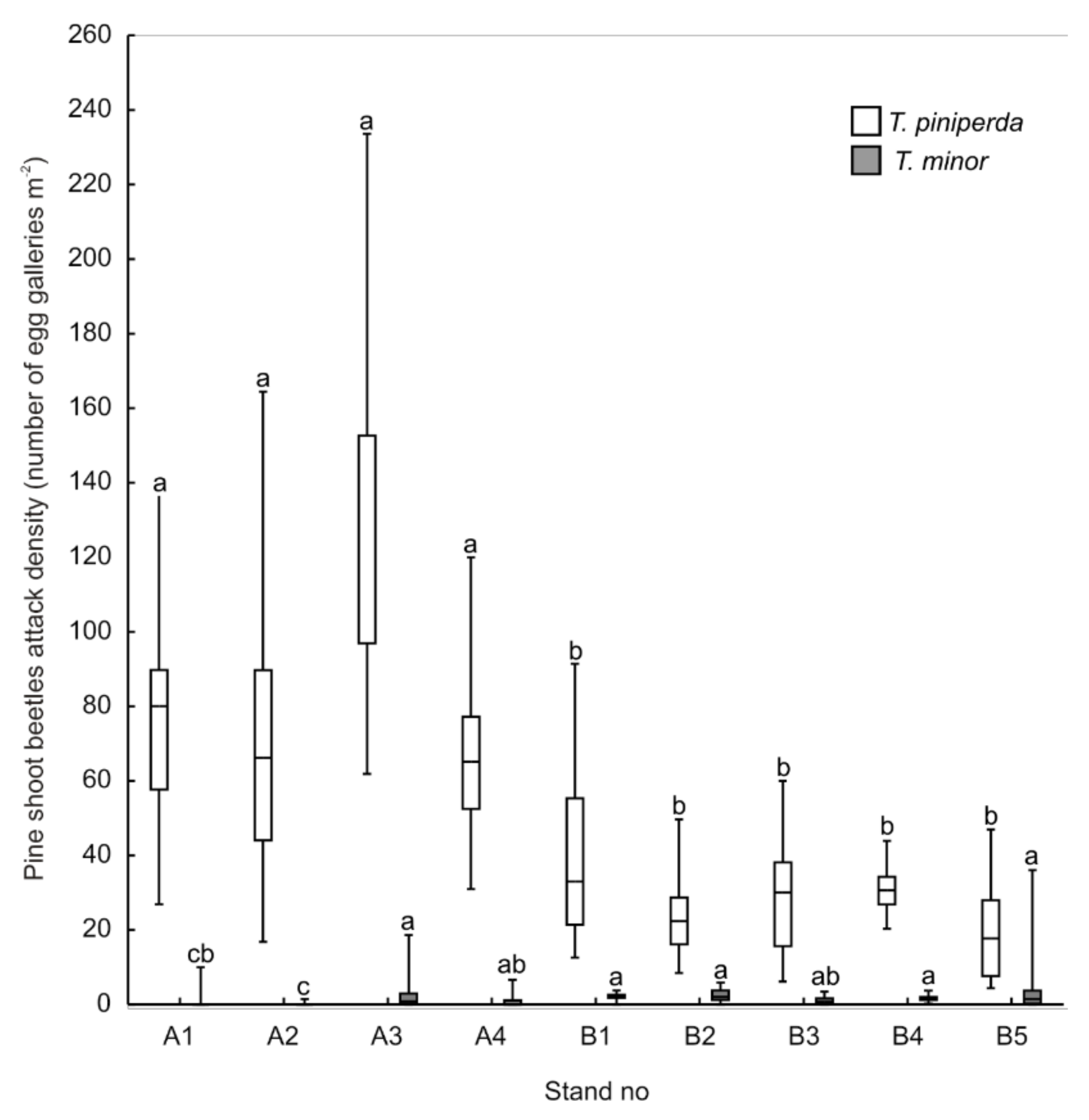
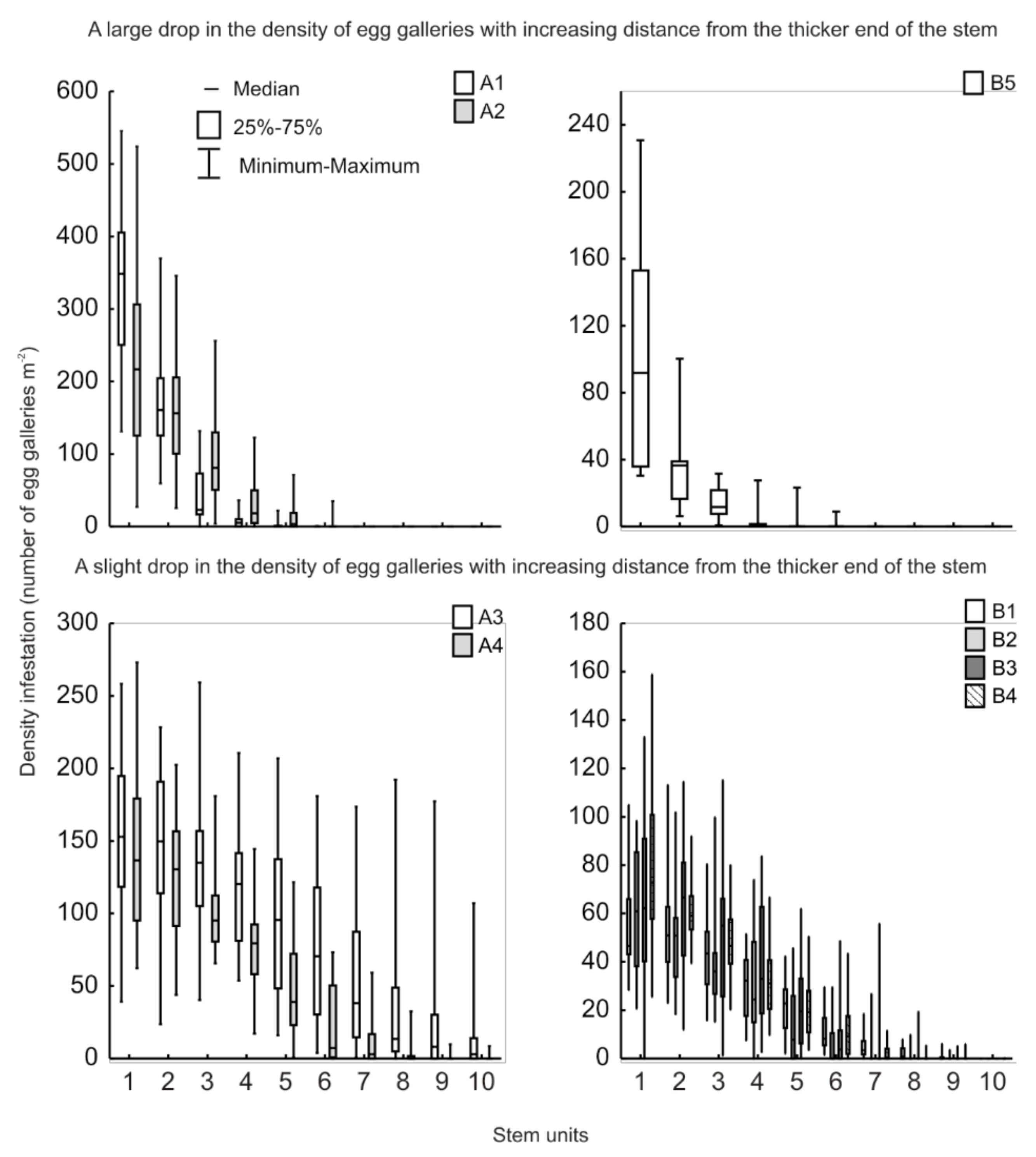
3.1. Colonisation of Trap Logs by Pine Shoot Beetles
3.2. Pine Stand Quality
3.3. Biotic Interactions in Populations of Pine Shoot Beetles
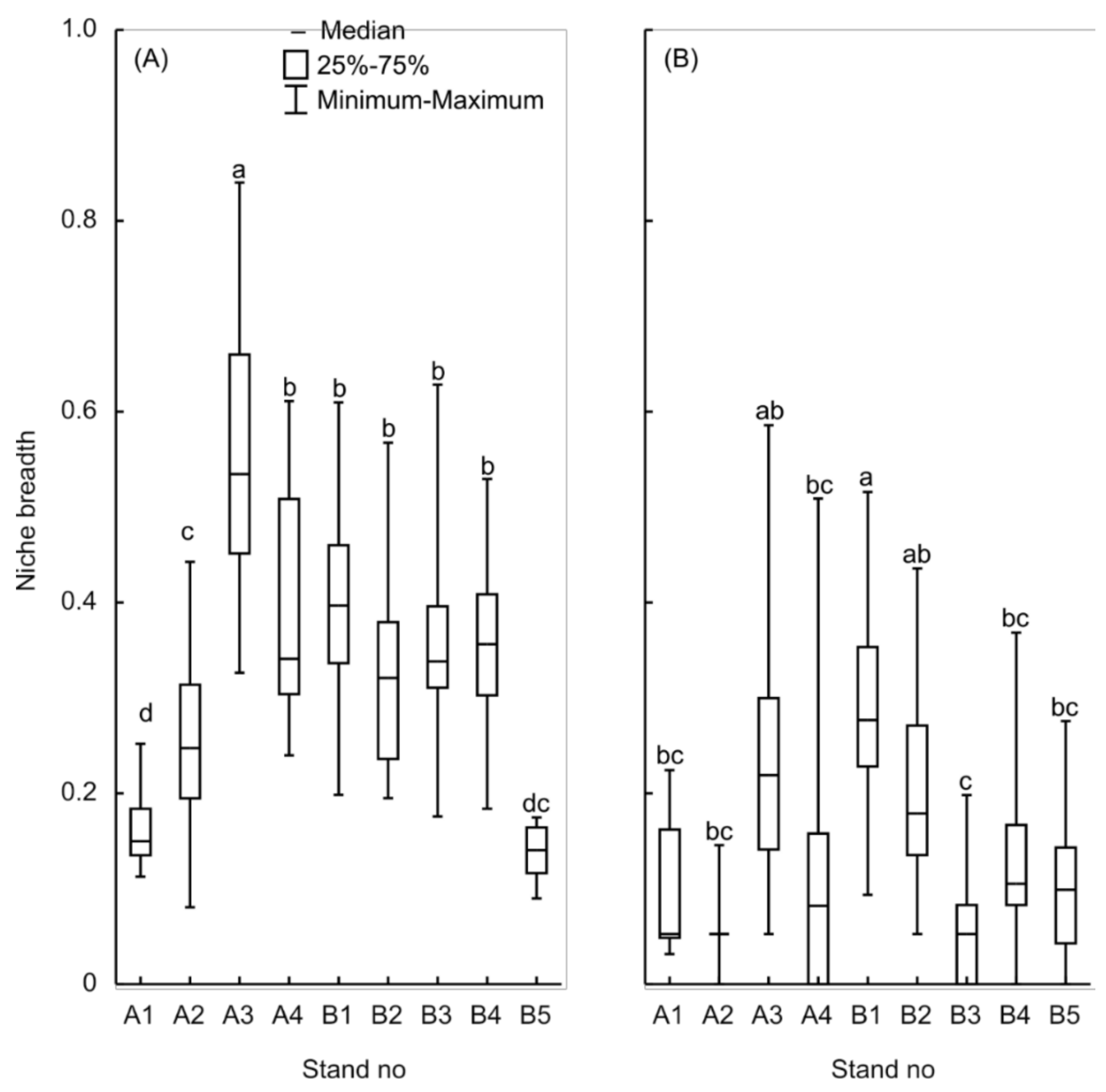
3.4. Tomicus Piniperda Niche Model
3.5. Evaluation of Accuracy of the Method
4. Discussion
4.1. Pine Stand Quality
4.2. Nutritional Suitability of Stems
4.3. Model Evaluation and Validation
- 1.
- Sample selection—determination of the number and distribution of P. sylvestris stems;
- 2.
- Estimation of niche breadth on the P. sylvestris stems selected in stage 1;
- 3.
- Estimation of the mean and confidence intervals for the T. piniperda niche breadth in a stand.
- 1.
- determination of the range of occurrence of galleries on stems;
- 2.
- evaluation of pine stand quality;
- 3.
- evaluation of the degree of damage to pine crowns.
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wood, S.L. The bark and ambrosia beetles of North and Central America (Coleoptera: Scolytidae), a taxonomic monograph. Great Basin Nat. Mem. 1982, 6, 1–1359. [Google Scholar]
- Grünwald, M. Ecological segregation of bark beetles (Coleoptera, Scolytidae) of spruce. Zeitschr. Angew. Entomol. 1986, 101, 176–187. [Google Scholar] [CrossRef]
- Rankin, L.J.; Borden, J.H. Competitive interactions between the mountain pine beetle and the pine engraver in lodgepole pine. Can. J. For. Res. 1991, 21, 1029–1036. [Google Scholar] [CrossRef]
- Flechtmann, C.A.H.; Dalusky, M.J.; Berisford, C.W. Bark and ambrosia beetle (Coleoptera: Scolytidae) responses to volatiles from aging loblolly pine billets. Environ. Entomol. 1999, 28, 638–648. [Google Scholar] [CrossRef]
- Saint-Germain, M.; Drapeau, P.; Buddle, C.M. Landing patterns of phloem- and wood-feeding Coleoptera on Black spruce of different physiological and decay states. Environ. Entomol. 2009, 38, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Barak, A.V.; McGrevy, D.; Tokaya, G. Dispersal and re-capture of marked, overwintering Tomicus piniperda (L.) (Coleoptera: Scolytidae) from Scotch pine bolts. Great Lakes Entomol. 2000, 33, 69–80. [Google Scholar]
- Långström, B.; Hellqvist, C. Spatial distribution of crown damage and growth losses caused by recurrent attacks of pine shoot beetles in pine stands surrounding a pulp mill in southern Sweden. J. Appl. Entomol. 1990, 110, 261–269. [Google Scholar] [CrossRef]
- Långström, B.; Hellqvist, C. Shoot damage and growth losses following three years of Tomicus-attacks in Scots pine stand close to a timber storage site. Silva Fenn. 1991, 25, 133–145. [Google Scholar] [CrossRef]
- Borkowski, A. Threats to pine stands by the pine shoot beetles Tomicus piniperda (L.) and T. minor (Hart.) around a sawmill in southern Poland. J. Appl. Entomol. 2001, 125, 489–492. [Google Scholar] [CrossRef]
- Öhrn, P. Seasonal Flight Patterns of the Spruce Bark Beetle (Ips typographus) in Sweden. Phenology, Voltininsm and Development. Licentiate Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2012; pp. 1–42. [Google Scholar]
- Carscadden, K.A.; Emery, N.C.; Arnillas, C.A.; Cadotte, M.W.; Afkhami, M.E.; Gravel, D.; Livingstone, S.W.; Wiens, J.J. Niche breadth: Causes and consequences for ecology, evolution, and conservation. Quart. Rev. Biol. 2020, 3, 179–214. [Google Scholar] [CrossRef]
- Paine, T.D.; Birch, M.C.; Švihra, P. Niche Breadth and resource partitioning by four sympatric species of bark beetles (Coleoptera: Scolytidae). Oecologia 1981, 48, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Amezaga, I.; Rodríguez, M.A. Resource partitioning of four sympatric bark beetles depending on swarming dates and tree species. For. Ecol. Manag. 1998, 109, 127–135. [Google Scholar] [CrossRef]
- Borkowski, A.; Skrzecz, I. Ecological segregation of bark beetles (Col., Curculionidae, Scolytinae) in Scots pine. Ecol. Res. 2016, 31, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Gao, B.T.; Ren, L.L.; Zong, S.X.; Ze, S.Z.; Luo, Y.Q. Niche-based relationship between sympatric bark living insect pests and tree vigor decline of Pinus yunnanensis. J. Appl. Entomol. 2019, 143, 1161–1171. [Google Scholar] [CrossRef]
- Lieutier, F.; Långström, B.; Faccoli, M. The Genus Tomicus. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2015; pp. 371–426. [Google Scholar]
- Borkowski, A. A spatial distribution of losses in growth of trees caused by feeding of pine shoot beetles Tomicus piniperda and T. minor (Col., Scolytidae) in Scots pine stands growing within the range of influence of a timber yard in southern Poland. J. For. Sci. 2006, 52, 130–135. [Google Scholar] [CrossRef] [Green Version]
- Grégoire, J.C.; Evans, H.F. Damage and control of BAWBILT organisms an overview. In Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis; Lieutier, F., Day, K.R., Battisti, A., Grégoire, J.C., Evans, H.F., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 19–37. [Google Scholar] [CrossRef]
- Långström, B.; Hellqvist, C.; Ehnström, B. Distribution and production of Tomicus piniperda in pine wood stacks. Sver. Skogs. Tidskr. 1984, 82, 23–35. [Google Scholar]
- Inoue, A. A model for the relationship between form-factors for stem volume and those for stem surface area in coniferous species. J. For. Res. 2006, 11, 289–294. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 4th ed.; W. H. Freeman and Co.: New York, NY, USA, 2012; pp. 1–887. [Google Scholar]
- Szymkiewicz, B. Tablice Zasobności i Przyrostu Drzewostanów; PWRiL: Warszawa, Poland, 2019; pp. 1–179. [Google Scholar]
- Näslund, M. Antalet prowtråd och hójdkurwans noggrannhet. Medd. Stat. Skogs. 1929, 25, 1–170. [Google Scholar]
- Levins, R. Evolution in Changing Environments: Some Theoretical Explorations; Princeton University Press: Princeton, NJ, USA, 1968; pp. 1–120. [Google Scholar]
- Hurlbert, S.H. The measurement of niche overlap and some relatives. Ecology 1978, 59, 67–77. [Google Scholar] [CrossRef]
- Morisita, M. Measuring of interspecific association and similarity between communities. Mem. Faculty Sci. Kyushu Univ. Ser. E 1959, 3, 65–80. [Google Scholar]
- Smith, E.P.; Zaret, T.M. Bias in estimating niche overlap. Ecology 1982, 63, 1248–1253. [Google Scholar] [CrossRef]
- Feinsinger, P.E.; Spears, E.; Poole, R.W. A Simple measure of niche breadth. Ecology 1981, 62, 27–32. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Homage to Santa Rosalia, or why are there so many kinds of Animals? Am. Nat. 1959, 93, 145–159. [Google Scholar] [CrossRef] [Green Version]
- StatSoft Inc. Statistica for Windows; StatSoft Inc.: Tulsa, OK, USA, 2020. [Google Scholar]
- Adlard, P.G. Myth and reality in growth estimation. For. Ecol. Manag. 1995, 71, 171–176. [Google Scholar] [CrossRef]
- Soares, P.; Tome, M.; Skovsgaard, J.P.; Vanclay, J.V. Evaluating a growth model for forest management using continuous forest inventory data. For. Ecol. Manag. 1995, 71, 251–265. [Google Scholar] [CrossRef]
- Kozak, A.; Kozak, R. Does cross validation provide additional information in the evaluation of regression models? Can. J. For. Res. 2003, 33, 976–987. [Google Scholar] [CrossRef]
- O’brien, R.M. A Caution regarding rules of thumb for variance inflation factors. Qual. Quant. 2007, 41, 673–690. [Google Scholar] [CrossRef]
- White, H.A. Heteroskedasticity-Consistent Covariance Matrix Estimator and a Direct Test for Heteroskedasticity. Econometrica 1980, 48, 17–838. [Google Scholar] [CrossRef]
- Temesgen, H.; Gadow, K. Generalized height–diameter models—an application for major tree species in complex stands of interior British Columbia. Eur. J. For. Res. 2004, 123, 45–51. [Google Scholar] [CrossRef]
- Thompson, S.K. Sampling; Wiley: New York, NY, USA, 2012; pp. 1–472. [Google Scholar] [CrossRef]
- Piene, H. Spruce budworm defoliation and growth loss in young balsam fir recovery of growth in spaced stands. Can. J. For. Res. 1989, 19, 1616–1624. [Google Scholar] [CrossRef]
- Nilsson, S. Increment losses caused by Blastophagus piniperda on Scots pine. R. Coll. For. Dept. Operat. Effic. Res. Notes 1974, 78, 1–64. [Google Scholar]
- Ericsson, A.; Hellqvist, C.; Långström, B.; Larsson, S.; Tenow, O. Effects on growth of simulated and induced shoot pruning by Tomicus piniperda as related to Carbohydrate and nitrogen dynamics in Scots pine. J. Appl. Ecol. 1985, 22, 105–124. [Google Scholar] [CrossRef]
- Bakke, A. Ecological studies on bark beetles (Coleoptera Scolytidae) associated with Scots pine (Pinus sylvestris) in Norway with particular reference to the influence of temperature. Medd. Nor. Skog. 1968, 21, 443–602. [Google Scholar]
- Hui, Y.; Xue-Song, D. Impacts of Tomicus minor on distribution and reproduction of Tomicus piniperda (Col., Scolytidae) on the trunk of the living Pinus yannanensis trees. J. Appl. Entomol. 2001, 123, 329–333. [Google Scholar] [CrossRef]
- Raffa, K.F.; Grégoire, J.C.; Lindgren, B.S. Natural history and ecology of bark beetles. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2015; pp. 1–40. [Google Scholar] [CrossRef]
- Inbar, M.; Wool, D. Phloem-feeding specialists sharing a host tree: Resource partitioning minimises interference competition among galling aphid species. Oikos 1995, 73, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Francke-Grosmann, H. Über die Ambrosiazucht der beiden Kieferborkenkäfer Myelophilus minor Htg. und Ips acuminatus Gyll. Medd. Stat. Skogs. 1952, 41, 1–52. [Google Scholar]
- Lekander, B.; Bejer-Petersen, B.; Kangas, E.; Bakke, A. The distribution of bark beetles in the Nordic countries. Acta Entomol. Fenn. 1977, 32, 1–36. [Google Scholar]
- Cochran, W.G. Sampling Techniques; Wiley: New York, NY, USA, 1977; pp. 1–600. [Google Scholar]
- Heurich, M.; Beudert, B.; Rall, H.; Křenová, Z. National parks as model regions for interdisciplinary long-term ecological research: The Bavarian Forest and Šumava National Parks underway to transboundary ecosystem research. In Long-Term Ecological Research; Müller, F., Baessler, C., Schubert, H., Klotz, S., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 327–344. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand No a | Distance from the Timber Yard (m) | Forest District | Pine Quality Classes b | Year of Cutting Trees | Number of Trap Logs | Stem Dimensions (Mean ± Standard Deviation) | ||
|---|---|---|---|---|---|---|---|---|
| Length (m) | Diameter Outsider Bark at | |||||||
| Thicker End (cm) | Thinner End (cm) | |||||||
| A1 | <100 | Zagnańsk | V | 2006 | 27 | 8.2 ± 0.9 | 20.6 ± 2.1 | 6.5 ± 0.6 |
| A2 | 100–300 | III | 1998, 2002 | 56 | 14.4 ± 2.7 | 24.2 ± 5.1 | 8.1 ± 1.9 | |
| A3 | >300 | Ia | 1996, 2003 | 30 | 13.9 ± 2.6 | 27.9 ± 4.8 | 7.5 ± 0.9 | |
| A4 | >300 | II | 2008 | 16 | 12.2 ± 1.6 | 23.2 ± 4.5 | 7.4 ± 1.5 | |
| B1 | >2000 | Przedbórz | III | 1992, 1993 | 40 | 12.2 ± 1.5 | 15.4 ± 1.8 | 7.0 ± 0.7 |
| B2 | Jędrzejów | III | 1998, 1999 | 25 | 12.7 ± 2.5 | 19.8 ± 3.4 | 7.5 ± 1.6 | |
| B3 | Dynów | III | 1996, 1997 | 25 | 16.5 ± 1.6 | 18.3 ± 3.1 | 9.7 ± 3.3 | |
| B4 | Koniecpol | III | 2000, 2001 | 30 | 10.0 ± 1.7 | 21.9 ± 2.9 | 10.3 ± 1.7 | |
| B5 | JanówLubelski | IV | 2005 | 10 | 12.3 ± 1.8 | 16.5 ± 2.5 | 9.0 ± 1.4 | |
| Stand No | T. Piniperda | T. Minor | t | d.f. | p | ||
|---|---|---|---|---|---|---|---|
| S.E. | S.E. | ||||||
| A1 | 0.15 a | 0.02 | 0.10 a | 0.04 | 1.1180 | 4 | 0.1687 |
| A2 | 0.26 a | 0.02 | 0.06 b | 0.02 | 9.0251 | 4 | 0.0008 |
| A3 | 0.58 a | 0.03 | 0.25 b | 0.03 | 9.6955 | 17 | <0.001 |
| A4 | 0.54 a | 0.05 | 0.11 b | 0.04 | 6.2584 | 10 | <0.001 |
| B1 | 0.39 a | 0.01 | 0.29 b | 0.02 | 5.9112 | 38 | <0.001 |
| B2 | 0.33 a | 0.02 | 0.21 b | 0.02 | 4.0550 | 22 | 0.0005 |
| B3 | 0.35 a | 0.02 | 0.06 b | 0.01 | 12.0026 | 20 | <0.001 |
| B4 | 0.35 a | 0.01 | 0.13 b | 0.01 | 13.1386 | 28 | <0.001 |
| B5 | 0.14 a | 0.01 | 0.11 a | 0.03 | 0.8603 | 8 | 0.4146 |
| Stand No | Proportional Similarity | Segregation | t Test (p) |
|---|---|---|---|
| A1 | 0.00 | + | <0.001 |
| A2 | 0.00 | + | <0.001 |
| A3 | 0.32 | + | <0.001 |
| A4 | 0.12 | + | <0.001 |
| B1 | 0.12 | + | <0.001 |
| B2 | 0.08 | + | <0.001 |
| B3 | 0.03 | + | <0.001 |
| B4 | 0.02 | + | <0.001 |
| B5 | 0.52 | + | 0.0479 |
| Name of Variable | Value of Parameter | T-Statistics Value | Probability Level | Vif | R2adj | RMSE | ANOVA | |
|---|---|---|---|---|---|---|---|---|
| F-Value | P-Level | |||||||
| Intercept | −0.0604 | −2.735 | 0.0074 | |||||
| rc | 0.0486 | 10.939 | <0.001 | 2.21 | ||||
| sqcIII | 0.0697 | 4.205 | <0.001 | 2.21 | ||||
| sqcI–II | 0.0796 | 3.076 | 0.0026 | 3.82 | ||||
| cu | 0.0468 | 3.628 | 0.0004 | 1.32 | 0.8132 | 0.056 | 117.45 | < 0.001 |
| Sample Plot No a | Mean ± SD | t | d.f. | Probability Level | |
|---|---|---|---|---|---|
| Real | Model | ||||
| A1 | 0.15 ± 0.04 | 0.14 ± 0.03 | 1.000 | 13 | 0.3356 |
| A2 | 0.25 ± 0.09 | 0.23 ± 0.04 | 2.000 | 28 | 0.0553 |
| A3 | 0.61 ± 0.16 | 0.54 ± 0.02 | 2.0463 | 15 | 0.0587 |
| A4 | 0.38 ± 0.13 | 0.38 ± 0.08 | 0.0132 | 8 | 0.9898 |
| B1 | 0.40 ± 0.08 | 0.40 ± 0.06 | 0.0939 | 20 | 0.9261 |
| B2 | 0.33 ± 0.10 | 0.31 ± 0.06 | 0.7383 | 13 | 0.4745 |
| B3 | 0.34 ± 0.11 | 0.34 ± 0.06 | −0.1211 | 14 | 0.9054 |
| B4 | 0.34 ± 0.09 | 0.37 ± 0.04 | −0.9398 | 16 | 0.3836 |
| B5 | 0.13 ± 0.03 | 0.15 ± 0.03 | −1.1073 | 4 | 0.3305 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borkowski, A. Resource Partitioning of Scots Pine (Pinus sylvestris L.) by Pine Shoot Beetles in Stands under Stress Conditions. Forests 2021, 12, 1336. https://doi.org/10.3390/f12101336
Borkowski A. Resource Partitioning of Scots Pine (Pinus sylvestris L.) by Pine Shoot Beetles in Stands under Stress Conditions. Forests. 2021; 12(10):1336. https://doi.org/10.3390/f12101336
Chicago/Turabian StyleBorkowski, Andrzej. 2021. "Resource Partitioning of Scots Pine (Pinus sylvestris L.) by Pine Shoot Beetles in Stands under Stress Conditions" Forests 12, no. 10: 1336. https://doi.org/10.3390/f12101336
APA StyleBorkowski, A. (2021). Resource Partitioning of Scots Pine (Pinus sylvestris L.) by Pine Shoot Beetles in Stands under Stress Conditions. Forests, 12(10), 1336. https://doi.org/10.3390/f12101336