
Forest Regeneration Patterns Differ Considerably between Sites with and without Windthrow Wood Logging in the High Tatra Mountains



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
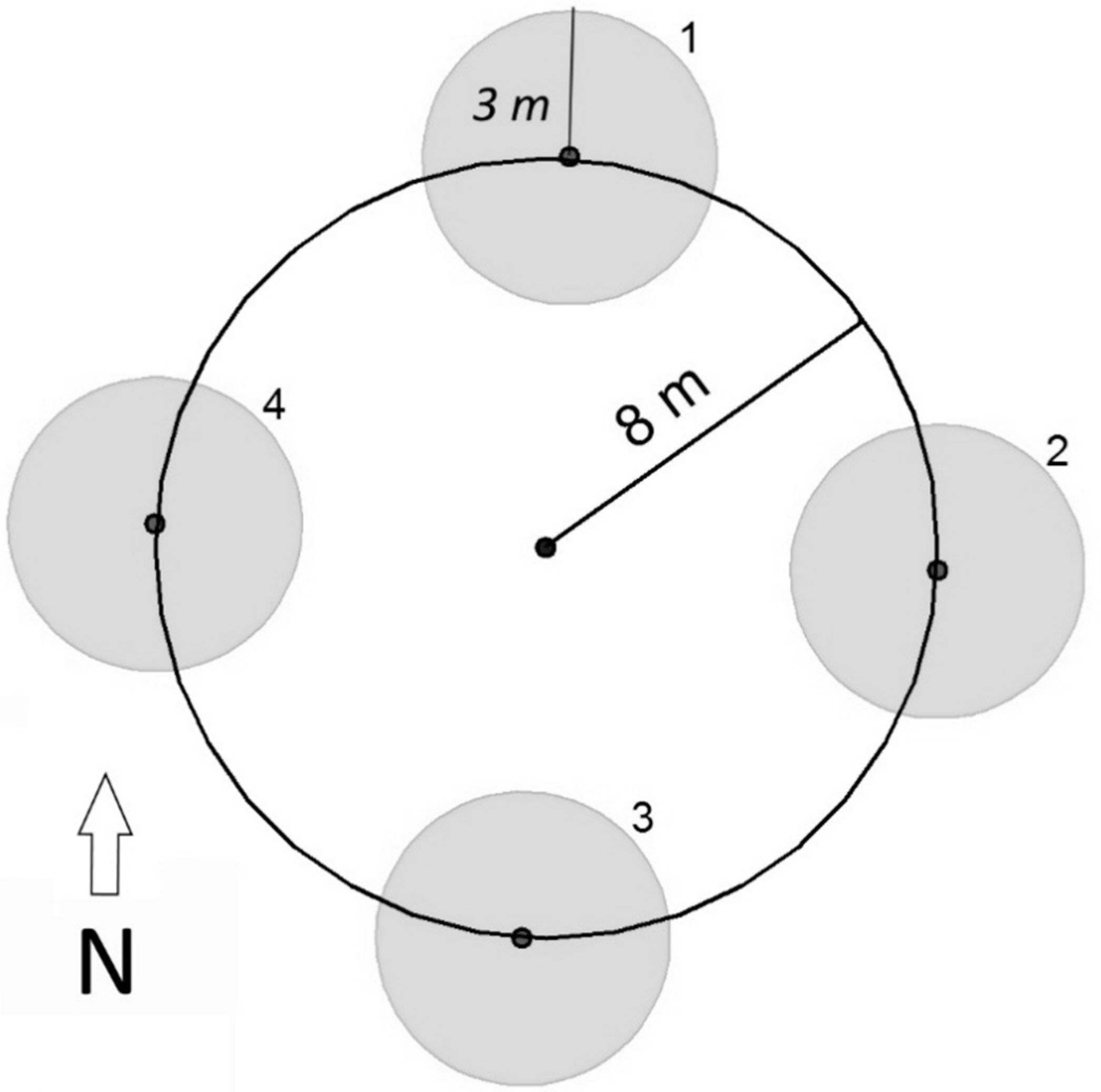
2.2. Tree Measurements
2.3. Data Processing and Statistical Approach
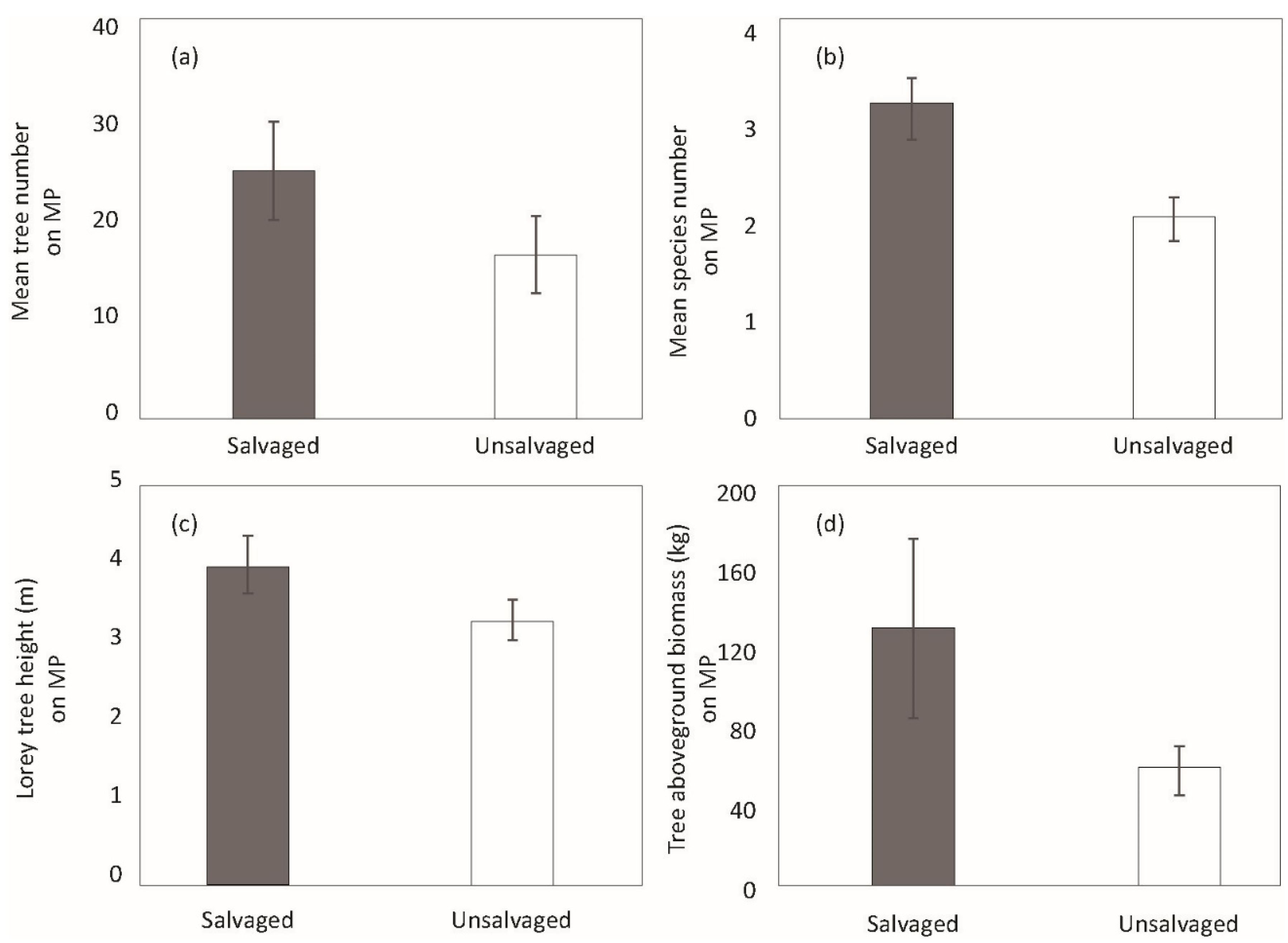
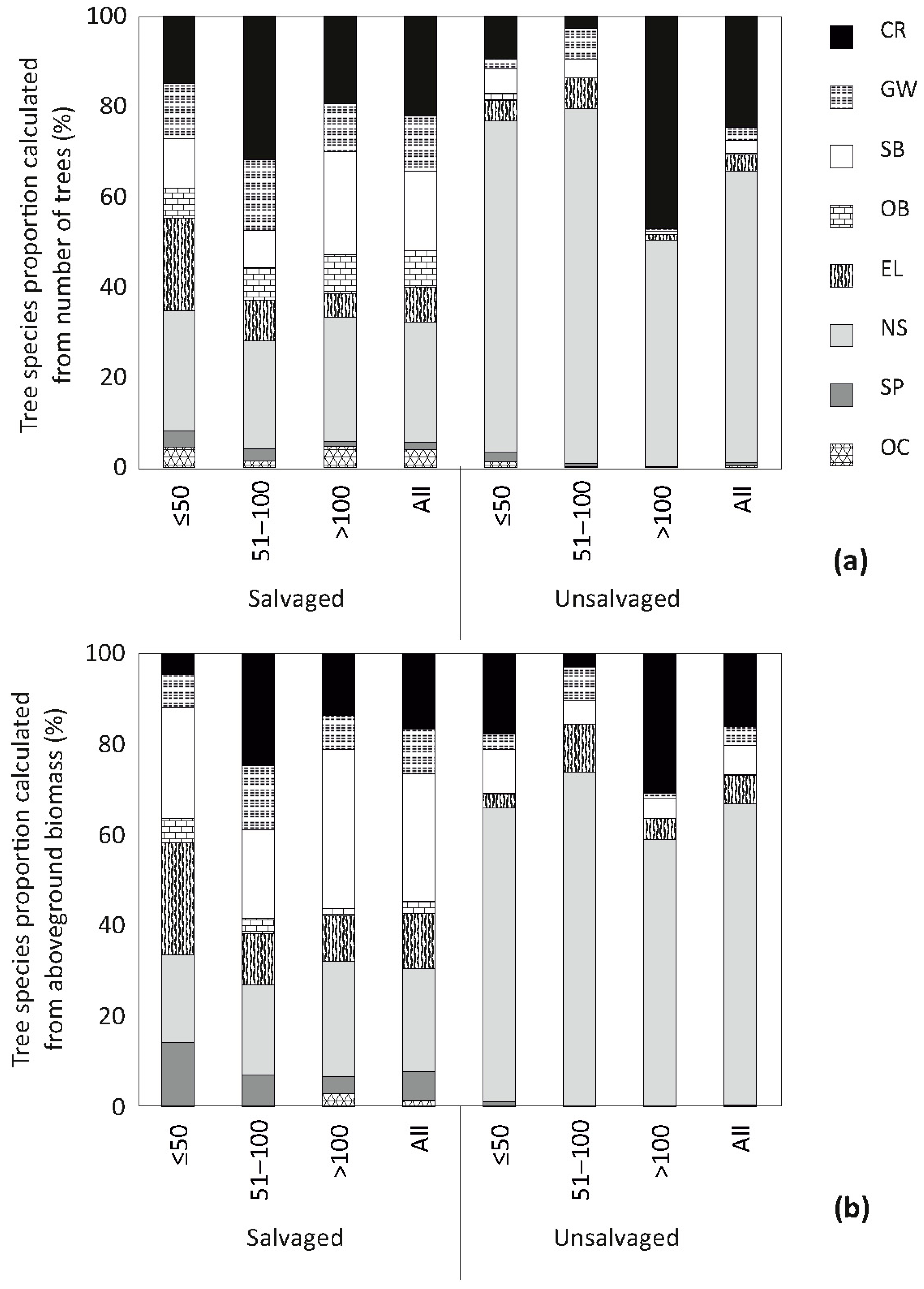
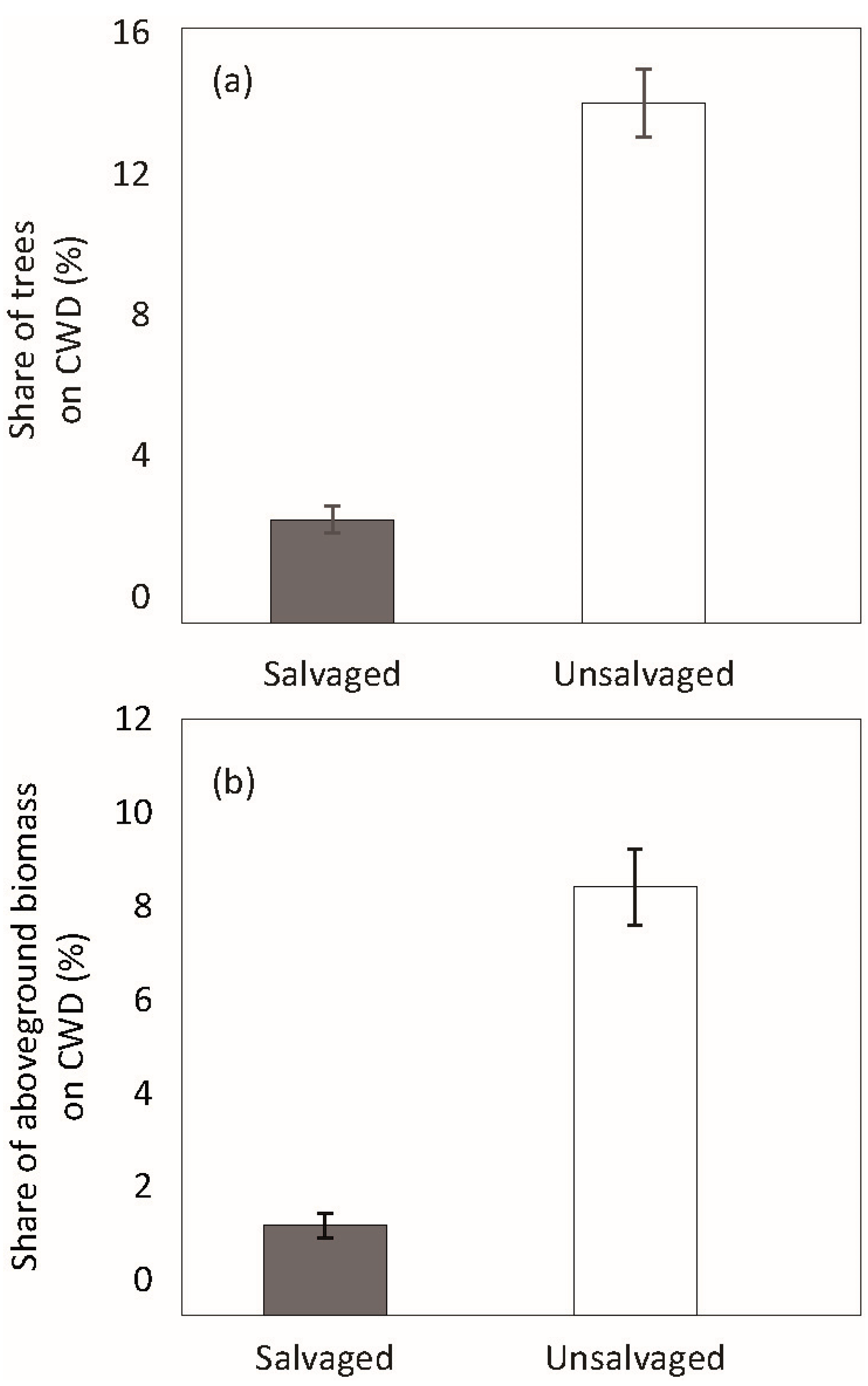
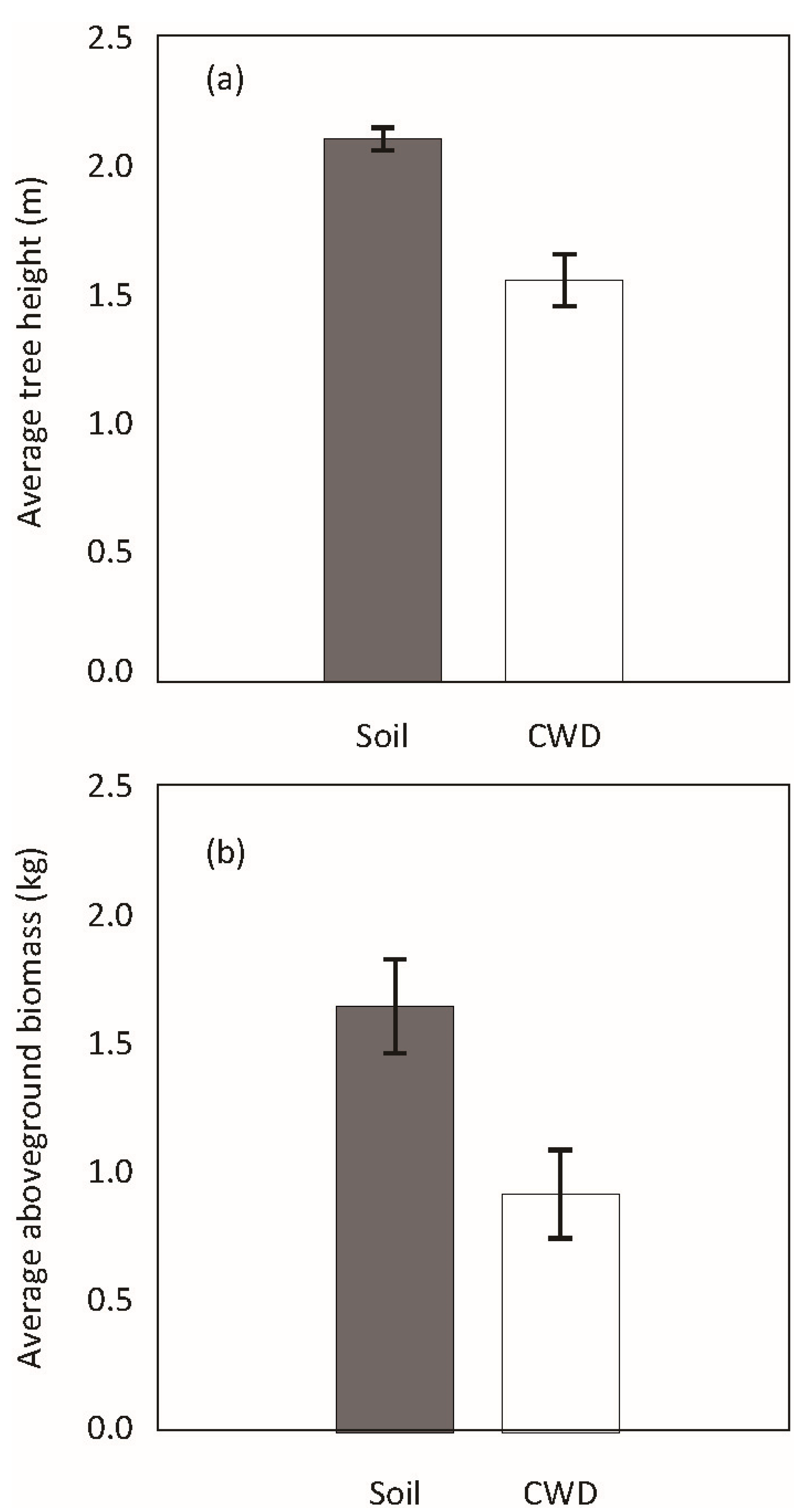
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Seidl, R.; Schelhaas, M.-J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef] [Green Version]
- Gregow, H.; Laaksonen, A.; Alper, M.E. Increasing large scale windstorm damage in Western, Central and Northern European forests, 1951–2010. Sci. Rep. 2017, 7, srep46397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink, A.H.; Brücher, T.; Ermert, V.; Krüger, A.; Pinto, J.G. The European storm Kyrill in January 2007: Synoptic evolution, meteorological impacts and some considerations with respect to climate change. Nat. Hazards Earth Syst. Sci. 2009, 9, 405–423. [Google Scholar] [CrossRef] [Green Version]
- Schmoeckel, J.; Kottmeier, C. Storm damage in the Black Forest caused by the winter storm “Lothar”—Part 1: Airborne damage assessment. Nat. Hazards Earth Syst. Sci. 2008, 8, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Seidl, R.; Rammer, W. Climate change amplifies the interactions between wind and bark beetle disturbances in forest landscapes. Landsc. Ecol. 2017, 32, 1485–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, N.; Nilsson, U.; Mossberg, M.; Bergh, J. Impacts of climate change, weather extremes and alternative strategies in managed forests. Écoscience 2018, 26, 53–70. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.; Peltola, H.; Väisänen, H.; Kellomäki, S. The effects of fragmentation on the susceptibility of a boreal forest ecosystem to wind damage. For. Ecol. Manag. 2009, 257, 1165–1173. [Google Scholar] [CrossRef]
- Konôpka, B.; Zach, P.; Kulfan, J. Wind–An important ecological factor and destructive agent in forests. For. J. 2016, 62, 123–130. [Google Scholar] [CrossRef]
- Merganič, J.; Merganičová, K.; Výbošťok, J.; Valent, P.; Bahýľ, J.; Yousefpour, R. Searching for Pareto Fronts for Forest Stand Wind Stability by Incorporating Timber and Biodiversity Values. Forests 2020, 11, 583. [Google Scholar] [CrossRef]
- Mitchell, S.J. Wind as a natural disturbance agent in forests: A synthesis. Int. J. For. Res. 2012, 86, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Kunca, A.; Zúbrik, M.; Galko, J.; Vakula, J.; Leontovyč, R.; Konôpka, B.; Nikolov, C.; Gubka, A.; Longauerová, V.; Maľová, M.; et al. Salvage felling in the Slovak forests in the period 2004–2013. For. J. 2015, 61, 188–195. [Google Scholar] [CrossRef] [Green Version]
- Koreň, M. Vetrová kalamita 19 novembra 2004: Nové pohľady a konsekvencie. Tatry 2005, 44, 6–29. (In Slovak) [Google Scholar]
- Konôpka, B.; Šebeň, V.; Pajtík, J. Species Composition and Carbon Stock of Tree Cover at a Postdisturbance Area in Tatra National Park, Western Carpathians. Mt. Res. Dev. 2019, 39, 71. [Google Scholar] [CrossRef]
- Vološčuk, I.; Bohuš, I.; Bublinec, E.; Hradiská, J.; Drdoš, J.; Dúbravcová, Z.; Gáper, J.; Greguš, C.; Haková, J.; Chovancová, B.; et al. Tatra National Park; Gradus Ltd.: Martin, Slovakia, 1994; p. 558. (In Slovak) [Google Scholar]
- Fleischer, P.; Pichler, V.; Fleischer, P.; Holko, L.; Máliš, F.; Gömöryová, E.; Cudlín, P.; Holeksa, J.; Michalová, Z.; Homolová, Z.; et al. Forest ecosystem services affected by natural disturbances, climate and land-use changes in the Tatra Mountains. Clim. Res. 2017, 73, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Wilson, E.; Leslie, A. Sustainable forest management. Geogr. Rev. 2009, 22, 8–13. [Google Scholar]
- Johnstone, J.F.; Allen, C.D.; Franklin, J.F.; Frelich, E.L.; Harvey, B.J.; Higuera, P.; Mack, M.C.; Meentemeyer, R.K.; Metz, M.R.; Perry, G.L.; et al. Changing disturbance regimes, ecological memory, and forest resilience. Front. Ecol. Environ. 2016, 14, 369–378. [Google Scholar] [CrossRef]
- Bergeron, Y.; Drapeau, P.; Gauthier, S.; LeComte, N. Using knowledge of natural disturbances to support sustainable forest management in the northern Clay Belt. For. Chron. 2007, 83, 326–337. [Google Scholar] [CrossRef] [Green Version]
- Slyder, J.B.; Wenzel, J.W.; Royo, A.A.; Spicer, M.E.; Carson, W.P. Post-windthrow salvage logging increases seedling and understory diversity with little impact on composition immediately after logging. New For. 2019, 51, 409–420. [Google Scholar] [CrossRef]
- Stokland, J.N.; Siitonen, J.; Jonsson, B.G. Biodiversity in Dead Wood; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Merganič, J.; Vorčák, J.; Merganičová, K.; Ďurský, J.; Miková, A.; Škvarenina, J.; Tuček, J.; Minďáš, J. Diversity Monitoring in Mountain Forests of Eastern Orava; EFRA Zvolen: Tvrdošín, Slovakia, 2003; p. 200. [Google Scholar]
- Vorčák, J.; Merganič, J.; Merganičová, K. Deadwood and spruce regeneration. Lesnická Práce 2005, 5, 18–19. (In Slovak) [Google Scholar]
- Vorčák, J.; Merganič, J.; Merganičová, K. Regeneration processes of natural spruce forest in the the subalpine forest belt of National Nature Reserve of Babia hora. In Babia Góra Nasze Wspólne Dziedzictwo; Jackowska, D.P., Ed.; Babiogórski Park Narodowy: Zawoja, Poland, 2005; pp. 81–94. (In Slovak) [Google Scholar]
- Vorčák, J.; Merganič, J.; Saniga, M. Structural diversity change and regeneration processes of the Norway spruce natural forest in Babia hora NNR in relation to altitude. J. For. Sci. 2006, 52, 399–409. [Google Scholar] [CrossRef] [Green Version]
- Bače, R.; Svoboda, M.; Pouska, V.; Janda, P.; Červenka, J. Natural regeneration in Central-European subalpine spruce forests: Which logs are suitable for seedling recruitment? For. Ecol. Manag. 2012, 266, 254–262. [Google Scholar] [CrossRef]
- Orman, O.; Szewczyk, J. European beech, silver fir, and Norway spruce differ in establishment, height growth, and mortality rates on coarse woody debris and forest floor—A study from a mixed beech forest in the Western Carpathians. Ann. For. Sci. 2015, 72, 955–965. [Google Scholar] [CrossRef]
- Bottero, A.; Garbarino, M.; Long, J.N.; Motta, R. The interacting ecological effects of large-scale disturbances and salvage logging on montane spruce forest regeneration in the western European Alps. For. Ecol. Manag. 2013, 292, 19–28. [Google Scholar] [CrossRef]
- Holeksa, J.; Zywiec, M.; Parusel, J.; Szewczyk, J.; Zielonka, T. Subalpine spruce forests in the Babia Góra National Park. In Structure, Production, Coarse Wood Debris and Regeneration Process of Norway Spruce Natural Forest in National Nature Re-Serves Babia Góra and Pilsko; Technical University: Zvolen, Slovakia, 2008; pp. 49–96. [Google Scholar]
- Milne-Rostkowska, F.; Holeksa, J.; Bogdziewicz, M.; Piechnik, Ł.; Seget, B.; Kurek, P.; Buda, J.; Żywiec, M. Where can palatable young trees escape herbivore pressure in a protected forest? For. Ecol. Manag. 2020, 472, 118221. [Google Scholar] [CrossRef]
- Michalová, Z.; Morrissey, R.C.; Wohlgemuth, T.; Bače, R.; Fleischer, P.; Svoboda, M. Salvage-Logging after Windstorm Leads to Structural and Functional Homogenization of Understory Layer and Delayed Spruce Tree Recovery in Tatra Mts., Slovakia. Forests 2017, 8, 88. [Google Scholar] [CrossRef] [Green Version]
- Vítková, L.; Bače, R.; Kjučukov, P.; Svoboda, M. Deadwood management in Central European forests: Key considerations for practical implementation. For. Ecol. Manag. 2018, 429, 394–405. [Google Scholar] [CrossRef]
- Senf, C.; Müller, J.; Seidl, R. Post-disturbance recovery of forest cover and tree height differ with management in Central Europe. Landsc. Ecol. 2019, 34, 2837–2850. [Google Scholar] [CrossRef] [Green Version]
- Erickson, A.; Nitschke, C.; Coops, N.; Cumming, S.; Stenhouse, G. Past-century decline in forest regeneration potential across a latitudinal and elevational gradient in Canada. Ecol. Model. 2015, 313, 94–102. [Google Scholar] [CrossRef]
- Kitenberga, M.; Elferts, D.; Adamovics, A.; Katrevics, J.; Donis, J.; Baders, E.; Jansons, A. Effect of salvage logging and forest type on the post-fire regeneration of Scots pine in hemiboreal forests. New For. 2020, 51, 1069–1085. [Google Scholar] [CrossRef]
- FAO. Global Forest Resources Assessment 2020. In Key Findings; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020; p. 12. [Google Scholar]
- Konôpka, B.; Pajtík, J.; Šebeň, V. Carbon stock change in forest stands biomass following a large-scale disturbance in the High Tatras. Rep. For. Res. 2017, 61, 239–246. [Google Scholar]
- Šebeň, V.; Konôpka, B. Tree height and species composition of young forest stands fifteen years after the large-scale wind disturbance in Tatra National Park. Cent. Eur. For. J. 2020, 66, 131–140. [Google Scholar]
- Marhefka, J. Forest revitalization after the windstorm calamity on November 19th 2004. TANAP Stud. 2015, 11, 83–94. [Google Scholar]
- Šebeň, V.; Bošeľa, M. Obnova lesa v Tatrách so spracovanou a nespracovanou kalamitou. In Proceedings of the International Conference, Nový Smokovec, Slovakia, 31 January–1 February 2011; pp. 76–82. (In Slovak). [Google Scholar]
- Pajtík, J.; Konôpka, B.; Šebeň, V. Mathematical Biomass Models for Young Individuals of Forest Tree Species in the Region of Western Carpathians; National Forest Centre, Forest Research Institute: Zvolen, Slovakia, 2018; p. 89. [Google Scholar]
- Lorey, T. Die mittlere Bestandeshöhe. Allg. Forst. J. Ztg. 1878, 54, 149–155. (In German) [Google Scholar]
- Core Team R. A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Reif, A.; Przybilla, M. Regeneration der Fichte in den Hochlagen des NP ayerischen Wald. Allg. Forst. Z. Waldwirtsch. 1998, 8, 400–403. (In German) [Google Scholar]
- Mai, W. Uber Ammenstamme im Gebirgswald; LWF Aktuell 18: Freising, Germany, 1999. (In German) [Google Scholar]
- San-Miguel-Ayanz, J.; de Rigo, D.; Caudallo, G. European Atlas of Forest Tree Species; Joint Research Centre, European Union: Luxembourg, 2016; p. 200. [Google Scholar]
- Caspersen, J.P.; Saprunoff, M. Seedling recruitment in a northern temperate forest: The relative importance of supply and establishment limitation. Can. J. For. Res. 2005, 35, 978–989. [Google Scholar] [CrossRef]
- Hagge, J.; Bässler, C.; Brandl, R.; Drexler, M.; Gruppe, A.; Hotes, S.; Hothorn, T.; Langhammer, P.; Müller, J.; Mysterud, A.; et al. Deadwood retention in forests lowers short-term browsing pressure on silver fir saplings by overabundant deer. For. Ecol. Manag. 2019, 451, 177531. [Google Scholar] [CrossRef]
- Côté, S.D.; Rooney, T.; Tremblay, J.-P.; Dussault, C.; Waller, D.M. Ecological Impacts of Deer Overabundance. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 113–147. [Google Scholar] [CrossRef] [Green Version]
- Merganič, J.; Russ, R.; Beranová, J.; Merganičová, K. Assessment of the impact of deer on the diversity of young trees in forest ecosystems in selected localities of the Czech Republic. Ekológia 2009, 28, 424–437. [Google Scholar] [CrossRef] [Green Version]
- Putman, R.J. Grazing in Temperate Ecosystems: Large Herbivores and the Ecology of the New Forest; Croom Helm: London, UK, 1986; p. 224. [Google Scholar]
- Gill, R.M.A. A Review of Damage by Mammals in North Temperate Forests: 3. Impact on Trees and Forests. Forrests 1992, 65, 363–388. [Google Scholar] [CrossRef] [Green Version]
- Reimoser, F.; Armstrong, H.; Suchant, R. Measuring forest damage of ungulates: What should be considered. For. Ecol. Manag. 1999, 120, 47–58. [Google Scholar] [CrossRef]
- Martin, J.L.; Daufresne, T. Introduced species and their impacts on the forest ecosystem of Haida Gwaii. In Proceedings of the Cedar Symposium, Queen Charlotte Island/Haida Qwaii, BC, Canada, 28–30 May 1996; Wiggins, G.G., Ed.; Ministry of Forests: Victoria, BC, Canada, 1999; pp. 69–85. [Google Scholar]
- Helle, T.; Aspi, J. Effects of Winter Grazing by Reindeer on Vegetation. Oikos 1983, 40, 337. [Google Scholar] [CrossRef]
- De Jager, N.R.; Pastor, J. Effects of simulated moose Alces alces browsing on the morphology of rowan Sorbus aucuparia. Wildl. Biol. 2010, 16, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Myking, T.; Solberg, E.J.; Austrheim, G.; Speed, J.D.M.; Bøhler, F.; Astrup, R.; Eriksen, R. Browsing of sallow (Salix caprea L.) and rowan (Sorbus aucuparia L.) in the context of life history strategies: A literature review. Eur. J. For. Res. 2013, 132, 399–409. [Google Scholar] [CrossRef]
- Konôpka, B.; Pajtík, J.; Shipley, L.A. Intensity of red deer browsing on young rowans differs between freshly-felled and standing individuals. For. Ecol. Manag. 2018, 429, 511–519. [Google Scholar] [CrossRef]
- Rooney, T. Deer impacts on forest ecosystems: A North American perspective. Forrests 2001, 74, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Motta, R. Ungulate impact on rowan (Sorbus aucuparia L.) and Norway spruce (Picea abies (L.) Karst.) height structure in mountain forests in the eastern Italian Alps. For. Ecol. Manag. 2003, 181, 139–150. [Google Scholar] [CrossRef]
- Motta, R.; Berretti, R.; Lingua, E.; Piussi, P. Coarse woody debris, forest structure and regeneration in the Valbona Forest Reserve, Paneveggio, Italian Alps. For. Ecol. Manag. 2006, 235, 155–163. [Google Scholar] [CrossRef]
- Harmon, M.E.; Franklin, J.F. Tree Seedlings on Logs in Picea-Tsuga Forests of Oregon and Washington. Ecolology 1989, 70, 48–59. [Google Scholar] [CrossRef]
- Santiago, L.S. Use of Coarse Woody Debris by the Plant Community of a Hawaiian Montane Cloud Forest. Biotropica 2000, 32, 633–641. [Google Scholar] [CrossRef]
- Zielonka, T. When does dead wood turn into a substrate for spruce replacement? J. Veg. Sci. 2006, 17, 739–746. [Google Scholar] [CrossRef]
- Merganičová, K.; Merganič, J. Coarse woody debris carbon stocks in natural spruce forests of Babia hora. J. For. Sci. 2010, 56, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Guo, X. Natural Regeneration on Coarse Woody Debris; UBC Undergraduate Research Graduation Essay, FRTS 497; University of British Columbia: Vancouver, BC, Canada, 2011. [Google Scholar] [CrossRef]
- Holeksa, J. Rozpad drzewostanu i odnowienie swierka a struktura i dynamika karpackiego boru gornoreglowego. Monogr. Bot. Lodz 1998, 82, 210. (In Polish) [Google Scholar]
- Lindenmayer, D.B.; Ough, K. Salvage logging in the mountain ash eucalypt forests of the Central Highlands of Victoria and its potential impacts on biodiversity. Conserv. Biol. 2006, 20, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Bouget, C.; Nusillard, B.; Pineau, X.; Ricou, C. Effect of deadwood position on saproxylic beetles in temperate forests and conservation interest of oak snags. Insect Conserv. Divers. 2012, 5, 264–278. [Google Scholar] [CrossRef]
- Tillon, L.; Bouget, C.; Paillet, Y.; Aulagnier, S. How does deadwood structure temperate forest bat assemblages? Eur. J. For. Res. 2016, 135, 433–449. [Google Scholar] [CrossRef]
- Thorn, S.; Seibold, S.; Leverkus, A.B.; Michler, T.; Müller, J.; Noss, R.F.; Stork, N.; Vogel, S.; Lindenmayer, D.B. The living dead: Acknowledging life after tree death to stop forest degradation. Front. Ecol. Environ. 2020, 18, 505–512. [Google Scholar] [CrossRef]
- Díaz, S.; Pascual, U.; Stenseke, M.; Marín-López, B.; Watson, R.T.; Molnár, Z.; Hill, R.; Chan, K.M.A.; Baste, I.A.; Brauman, K.A.; et al. Assessing nature’s contributions to people. Science 2018, 359, 270–272. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monitoring Plots | Altitude Range (m a.s.l.) | Amount of Deadwood * (m3 ha−1) | Number of Recorded Trees | Number of Recorded Tree Species ** | Mean Lorey’s Height (m) |
|---|---|---|---|---|---|
| Salvaged | 865–1348 | 209 ± 23 | 1876 | 11 | 4.0 |
| Unsalvaged | 853–1388 | 40 ± 3 | 1206 | 6 | 3.3 |
| Tree Species— English (Abbrev.) | Tree Species—Latin Name | Monitoring Plots | Number of Measured Trees | Tree Height Characteristics (m) | |||
|---|---|---|---|---|---|---|---|
| Mean | SD | Min. | Max. | ||||
| Common rowan (CR) | Sorbus aucuparia | Salvaged | 436 | 1.9 | 1.6 | 0.2 | 7.5 |
| Unsalvaged | 207 | 2.3 | 1.4 | 0.1 | 6.0 | ||
| Goat willow (GW) | Salix caprea | Salvaged | 241 | 2.4 | 1.7 | 0.3 | 7.0 |
| Unsalvaged | 37 | 2.3 | 1.8 | 0.3 | 6.1 | ||
| Silver birch (SB) | Betula pendula | Salvaged | 321 | 2.6 | 2.0 | 0.2 | 9.0 |
| Unsalvaged | 38 | 2.4 | 1.9 | 0.2 | 6.2 | ||
| Other broadleaves (OB) | Salvaged | 161 | 1.7 | 1.5 | 0.3 | 6.0 | |
| Unsalvaged | 5 | 1.0 | 0.9 | 0.4 | 2.6 | ||
| Europan larch (EL) | Larix decidua | Salvaged | 147 | 2.4 | 1.4 | 0.2 | 7.5 |
| Unsalvaged | 49 | 2.0 | 1.4 | 0.2 | 5.5 | ||
| Norway spruce (NS) | Picea abies | Salvaged | 451 | 2.0 | 1.3 | 0.2 | 6.0 |
| Unsalvaged | 851 | 2.1 | 1.3 | 0.1 | 7.0 | ||
| Scots pine (SP) | Pinus sylvestris | Salvaged | 30 | 2.9 | 1.4 | 0.8 | 5.5 |
| Unsalvaged | 11 | 1.1 | 0.8 | 0.3 | 3.2 | ||
| Other coniferous (OC) | Salvaged | 47 | 1.4 | 1.2 | 0.2 | 6.0 | |
| Unsalvaged | 5 | 1.0 | 0.7 | 0.3 | 2.0 | ||
| All species | Salvaged | 1834 | 2.2 | 1.6 | 0.2 | 9.0 | |
| Unsalvaged | 1203 | 2.1 | 1.4 | 0.1 | 7.0 | ||
| All species | All | 3037 | 2.2 | 1.5 | 0.1 | 9.0 | |
| Density Group (Trees Per Are) | Monitoring Plots | Number of Plots | Share (%) | Average Number of Trees Per Are | Average Tree Height (m) |
|---|---|---|---|---|---|
| Below 50 | Salvaged | 34 | 42.4 | 24.1 ± 13.0 | 2.2 ± 1.0 |
| Unsalvaged | 47 | 58.7 | 24.3 ± 13.7 | 2.2 ± 0.9 | |
| 51–100 | Salvaged | 23 | 28.8 | 76.7 ± 13.1 | 2.4 ± 0.9 |
| Unsalvaged | 21 | 26.3 | 68.8 ± 11.7 | 2.2 ± 0.6 | |
| Over 100 | Salvaged | 23 | 28.8 | 196.1 ± 70.7 | 2.3 ± 0.9 |
| Unsalvaged | 12 | 15.0 | 183.1 ± 83.0 | 2.1 ± 0.3 | |
| All plots | Salvaged | 80 | 100.0 | 90.3 ± 82.2 | 2.3 ± 0.9 |
| Unsalvaged | 80 | 100.0 | 60.2 ± 54.9 | 2.2 ± 0.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konôpka, B.; Šebeň, V.; Merganičová, K. Forest Regeneration Patterns Differ Considerably between Sites with and without Windthrow Wood Logging in the High Tatra Mountains. Forests 2021, 12, 1349. https://doi.org/10.3390/f12101349
Konôpka B, Šebeň V, Merganičová K. Forest Regeneration Patterns Differ Considerably between Sites with and without Windthrow Wood Logging in the High Tatra Mountains. Forests. 2021; 12(10):1349. https://doi.org/10.3390/f12101349
Chicago/Turabian StyleKonôpka, Bohdan, Vladimír Šebeň, and Katarína Merganičová. 2021. "Forest Regeneration Patterns Differ Considerably between Sites with and without Windthrow Wood Logging in the High Tatra Mountains" Forests 12, no. 10: 1349. https://doi.org/10.3390/f12101349
APA StyleKonôpka, B., Šebeň, V., & Merganičová, K. (2021). Forest Regeneration Patterns Differ Considerably between Sites with and without Windthrow Wood Logging in the High Tatra Mountains. Forests, 12(10), 1349. https://doi.org/10.3390/f12101349