Aboveground Biomass of Living Trees Depends on Topographic Conditions and Tree Diversity in Temperate Montane Forests from the Slătioara-Rarău Area (Romania)

 ,
,  , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
- assessing tree species diversity, tree size heterogeneity and aboveground biomass of living trees at sample plot level;
- identifying the effect of some abiotic factors on tree species diversity and aboveground biomass;
- identifying how aboveground biomass depends on tree species diversity and size heterogeneity, within the entire study area and at habitat type level.
2. Materials and Methods
2.1. Study Location
2.2. Data Collection
2.3. Data Processing
2.3.1. GIS Data Processing
2.3.2. Diversity Indices Computation
2.3.3. Aboveground Biomass Assessment
2.3.4. Data Analysis
3. Results
3.1. Overview of Tree Species, Forest Habitats and Aboveground Biomass
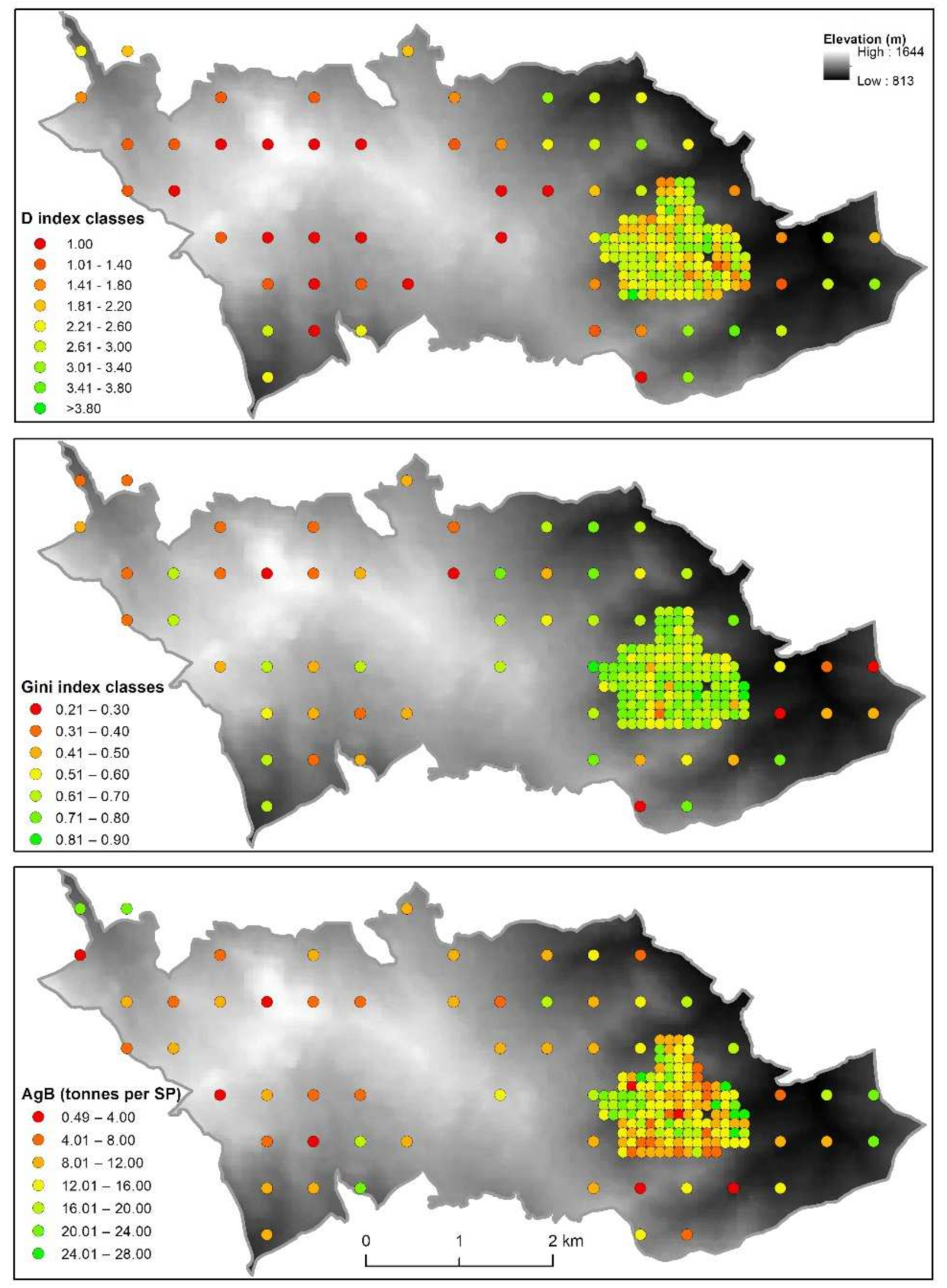
3.2. Relationships between the Attributes of the SPs
3.3. Influence of Ecological Factors on Forest Diversity and AgB
3.4. Tree Species Richness/Size Heterogeneity and AgB Relations
3.5. Influence of Forest Diversity on AgB of Living Trees by Habitat Type
4. Discussion
4.1. Tree Species/Size Diversity and AgB Relations
4.2. Aboveground Biomass and Topographical Parameters
4.3. Tree Diversity Influence on Aboveground Biomass
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Watson, J.E.M.; Evans, T.; Venter, O.; Williams, B.; Tulloch, A.; Stewart, C.; Thompson, I.; Ray, J.C.; Murray, K.; Salazar, A.; et al. The exceptional value of intact forest ecosystems. Nat. Ecol. Evol. 2018, 2, 599–610. [Google Scholar] [CrossRef] [PubMed]
- Lähde, E.; Eskelinen, T.; Väänänen, A. Growth and diversity effects of silvicultural alternatives on an old-growth forest in Finland. For. Int. J. For. Res. 2002, 75, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Pastur, G.M.; Lencinas, M.V.; Cellini, J.M.; Peri, P.L.; Soler Esteban, R. Timber management with variable retention in Nothofagus pumilio forests of Southern Patagonia. For. Ecol. Manag. 2009, 258, 436–443. [Google Scholar] [CrossRef]
- Brunet, J.; Örjan, F.; Richnau, G. Biodiversity in European beech forests—A review with recommendations for sustainable forest management. Ecol. Bull. 2010, 53, 77–94. [Google Scholar]
- Larson, A.J.L.J.; Franklin, J.F.F.F. The tree mortality regime in temperate old-growth coniferous forests: The role of physical damage. Can. J. For. Res. 2010, 40, 2091–2103. [Google Scholar] [CrossRef] [Green Version]
- Pastur, G.J.M.; Peri, P.L.; Cellini, J.M.; Lencinas, M.V.; Barrera, M.; Ivancich, H. Canopy structure analysis for estimating forest regeneration dynamics and growth in Nothofagus pumilio forests. Ann. For. Sci. 2011, 68, 587–594. [Google Scholar] [CrossRef]
- Foster, D.R.; Orwig, D.A.; McLachlan, J.S. Ecological and conservation insights from reconstructive studies of temperate old-growth forests. Trends Ecol. Evol. 1996, 11, 419–424. [Google Scholar] [CrossRef]
- Sabatini, F.M.; Burrascano, S.; Keeton, W.S.; Levers, C.; Lindner, M.; Pötzschner, F.; Verkerk, P.J.; Bauhus, J.; Buchwald, E.; Chaskovsky, O.; et al. Where are Europe’s last primary forests? Divers. Distrib. 2018, 24, 1426–1439. [Google Scholar] [CrossRef] [Green Version]
- Nikolov, S.C. Effect of stand age on bird communities in late-successional Macedonian pine forests in Bulgaria. For. Ecol. Manag. 2009, 257, 580–587. [Google Scholar] [CrossRef]
- Zemanová, L.; Trotsiuk, V.; Morrissey, R.C.; Bače, R.; Mikoláš, M.; Svoboda, M. Old trees as a key source of epiphytic lichen persistence and spatial distribution in mountain Norway spruce forests. Biodivers. Conserv. 2017, 26, 1943–1958. [Google Scholar] [CrossRef]
- Zhou, G.; Liu, S.; Li, Z.; Zhang, D.; Tang, X.; Zhou, C.; Yan, J.; Mo, J. Old-growth forests can accumulate carbon in soils. Science 2006, 314, 1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seedre, M.; Kopáček, J.; Janda, P.; Bače, R.; Svoboda, M. Carbon pools in a montane old-growth Norway spruce ecosystem in Bohemian Forest: Effects of stand age and elevation. For. Ecol. Manag. 2015, 346, 106–113. [Google Scholar] [CrossRef]
- Meyer, P. Network of Strict Forest Reserves as reference system for close to nature forestry in Lower Saxony, Germany|Request PDF. For. Snow Landsc. Res. 2005, 33–44. [Google Scholar]
- Nagel, T.A.; Mikac, S.; Dolinar, M.; Klopcic, M.; Keren, S.; Svoboda, M.; Diaci, J.; Boncina, A.; Paulic, V. The natural disturbance regime in forests of the Dinaric Mountains: A synthesis of evidence. For. Ecol. Manag. 2017, 388, 29–42. [Google Scholar] [CrossRef]
- Paquette, A.; Messier, C. The effect of biodiversity on tree productivity: From temperate to boreal forests. Glob. Ecol. Biogeogr. 2011, 20, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, 8957. [Google Scholar] [CrossRef] [Green Version]
- Van Der Plas, F.; Manning, P.; Allan, E.; Scherer-Lorenzen, M.; Verheyen, K.; Wirth, C.; Zavala, M.A.; Hector, A.; Ampoorter, E.; Baeten, L.; et al. Jack-of-all-trades effects drive biodiversity-ecosystem multifunctionality relationships in European forests. Nat. Commun. 2016, 7, 11109. [Google Scholar] [CrossRef] [Green Version]
- Bohn, F.J.; Huth, A. The importance of forest structure to biodiversity–productivity relationships. R. Soc. Open Sci. 2016, 4, 160521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Benito, P.; Ratcliffe, S.; Jump, A.S.; Gómez-Aparicio, L.; Madrigal-González, J.; Wirth, C.; Kändler, G.; Lehtonen, A.; Dahlgren, J.; Kattge, J.; et al. Functional diversity underlies demographic responses to environmental variation in European forests. Glob. Ecol. Biogeogr. 2017, 26, 128–141. [Google Scholar] [CrossRef] [Green Version]
- Ammer, C. Diversity and forest productivity in a changing climate. New Phytol. 2019, 221, 50–66. [Google Scholar] [CrossRef] [Green Version]
- Brun, P.; Zimmermann, N.E.; Graham, C.H.; Lavergne, S.; Pellissier, L.; Münkemüller, T.; Thuiller, W. The productivity-biodiversity relationship varies across diversity dimensions. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–76. [Google Scholar] [CrossRef]
- Loreau, M.; Hector, A. Not even wrong: Comment by Loreau and Hector. Ecology 2019, 100, e02794. [Google Scholar] [CrossRef] [PubMed]
- Pillai, P.; Gouhier, T.C. Not even wrong: The spurious measurement of biodiversity’s effects on ecosystem functioning. Ecology 2019, 100, e02645. [Google Scholar] [CrossRef] [Green Version]
- Pillai, P.; Gouhier, T.C. Not even wrong: Reply to Loreau and Hector. arXiv 2019, arXiv:1910.13563. [Google Scholar]
- Pillai, P.; Gouhier, T.C. Not even wrong: Reply to Wagg et al. arXiv 2019, arXiv:1910.13670. [Google Scholar]
- Wagg, C.; Barry, K.E.; O’Brien, M.J.; McKenzie-Gopsill, A.; Roscher, C.; Eisenhauer, N.; Schmid, B. Not even wrong: Comment by Wagg et al. Ecology 2019, 100, e02805. [Google Scholar] [CrossRef]
- Naeem, S.; Duffy, J.E.; Zavaleta, E. The functions of biological diversity in an age of extinction. Science 2012, 336, 1401–1406. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Benito, P.; Gómez-Aparicio, L.; Paquette, A.; Messier, C.; Kattge, J.; Zavala, M.A. Diversity increases carbon storage and tree productivity in Spanish forests. Glob. Ecol. Biogeogr. 2014, 23, 311–322. [Google Scholar] [CrossRef]
- Dellasala, D.A.; Kormos, C.F.; Keith, H.; Mackey, B.; Young, V.; Rogers, B.; Mittermeier, R.A. Primary Forests Are Undervalued in the Climate Emergency. Bioscience 2020, 70, 445. [Google Scholar] [CrossRef]
- Vilà, M.; Carrillo-Gavilán, A.; Vayreda, J.; Bugmann, H.; Fridman, J.; Grodzki, W.; Haase, J.; Kunstler, G.; Schelhaas, M.; Trasobares, A. Disentangling Biodiversity and Climatic Determinants of Wood Production. PLoS ONE 2013, 8, e53530. [Google Scholar] [CrossRef] [Green Version]
- Ratcliffe, S.; Wirth, C.; Jucker, T.; Plas, F. van der; Scherer-Lorenzen, M.; Verheyen, K.; Allan, E.; Benavides, R.; Bruelheide, H.; Ohse, B.; et al. Biodiversity and ecosystem functioning relations in European forests depend on environmental context. Ecol. Lett. 2017, 20, 1414–1426. [Google Scholar] [CrossRef]
- Baritz, R.; Seufert, G.; Montanarella, L.; Van Ranst, E. Carbon concentrations and stocks in forest soils of Europe. For. Ecol. Manag. 2010, 260, 262–277. [Google Scholar] [CrossRef]
- Luyssaert, S.; Marie, G.; Valade, A.; Chen, Y.Y.; Njakou Djomo, S.; Ryder, J.; Otto, J.; Naudts, K.; Lansø, A.S.; Ghattas, J.; et al. Trade-offs in using European forests to meet climate objectives. Nature 2018, 562, 259–262. [Google Scholar] [CrossRef]
- Bodin, P.; Wiman, B.L.B. The usefulness of stability concepts in forest management when coping with increasing climate uncertainties. For. Ecol. Manag. 2007, 242, 541–552. [Google Scholar] [CrossRef]
- Oikonomakis, N.; Ganatsas, P. Land cover changes and forest succession trends in a site of Natura 2000 network (Elatia forest), in northern Greece. For. Ecol. Manag. 2012, 285, 153–163. [Google Scholar] [CrossRef]
- Silva Pedro, M.; Rammer, W.; Seidl, R. Tree species diversity mitigates disturbance impacts on the forest carbon cycle. Oecologia 2014, 177, 619–630. [Google Scholar] [CrossRef]
- Silva Pedro, M.; Rammer, W.; Seidl, R. Disentangling the effects of compositional and structural diversity on forest productivity. J. Veg. Sci. 2017, 28, 649–658. [Google Scholar] [CrossRef]
- Dănescu, A.; Albrecht, A.T.; Bauhus, J. Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany. Oecologia 2016, 182, 319–333. [Google Scholar] [CrossRef]
- Tahvonen, O.; Pukkala, T.; Laiho, O.; Lähde, E.; Niinimäki, S. Optimal management of uneven-aged Norway spruce stands. For. Ecol. Manag. 2010, 260, 106–115. [Google Scholar] [CrossRef]
- Schall, P.; Schulze, E.D.; Fischer, M.; Ayasse, M.; Ammer, C. Relations between forest management, stand structure and productivity across different types of Central European forests. Basic Appl. Ecol. 2018, 32, 39–52. [Google Scholar] [CrossRef]
- Schall, P.; Gossner, M.M.; Heinrichs, S.; Fischer, M.; Boch, S.; Prati, D.; Jung, K.; Baumgartner, V.; Blaser, S.; Böhm, S.; et al. The impact of even-aged and uneven-aged forest management on regional biodiversity of multiple taxa in European beech forests. J. Appl. Ecol. 2018, 55, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhu, Y.; Wang, J.; Ma, W.; Meng, J. Species association of the dominant tree species in an old-growth forest and implications for enrichment planting for the restoration of natural degraded forest in subtropical China. Forests 2019, 10, 957. [Google Scholar] [CrossRef] [Green Version]
- Ammer, C. Unraveling the Importance of Inter- and Intraspecific Competition for the Adaptation of Forests to Climate Change. In Progress in Botany; Cánovas, F.M., Lüttge, U.M.R., Eds.; Springer: Cham, Switzerland, 2016; pp. 345–367. [Google Scholar]
- Ehbrecht, M.; Schall, P.; Ammer, C.; Seidel, D. Quantifying stand structural complexity and its relationship with forest management, tree species diversity and microclimate. Agric. For. Meteorol. 2017, 242, 1–9. [Google Scholar] [CrossRef]
- Gough, C.M.; Atkins, J.W.; Fahey, R.T.; Hardiman, B.S. High rates of primary production in structurally complex forests. Ecology 2019, 100, e02864. [Google Scholar] [CrossRef] [Green Version]
- Jucker, T.; Avăcăritei, D.; Bărnoaiea, I.; Duduman, G.; Bouriaud, O.; Coomes, D.A. Climate modulates the effects of tree diversity on forest productivity. J. Ecol. 2016, 104, 12522. [Google Scholar] [CrossRef] [Green Version]
- Duduman, G.; Roibu, C.-C.; Duduman, M.-L.; Miron-Onciul, M. The influence of competition and dimensional- spatial characteristics of trees on their radial growth in Old-Growth Slătioara forest, Romania. Adv. Environ. Sci.–Int. J. Bioflux Soc. 2010, 2, 215–230. [Google Scholar]
- Fyllas, N.M.; Bentley, L.P.; Shenkin, A.; Asner, G.P.; Atkin, O.K.; Díaz, S.; Enquist, B.J.; Farfan-Rios, W.; Gloor, E.; Guerrieri, R.; et al. Solar radiation and functional traits explain the decline of forest primary productivity along a tropical elevation gradient. Ecol. Lett. 2017, 20, 730–740. [Google Scholar] [CrossRef]
- Leathwick, J.R.; Burns, B.R.; Clarkson, B.D. Environmental correlates of tree alpha-diversity in New Zealand primary forests. Ecography 1998, 21, 235–246. [Google Scholar] [CrossRef]
- Stevens, G.C. The Latitudinal Gradient in Geographical Range: How so Many Species Coexist in the Tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- Zhu, Z.X.; Nizamani, M.M.; Sahu, S.K.; Kunasingam, A.; Wang, H.F. Tree abundance, richness, and phylogenetic diversity along an elevation gradient in the tropical forest of Diaoluo Mountain in Hainan, China. Acta Oecol. 2019, 101, 103481. [Google Scholar] [CrossRef]
- Leuschner, C.; Moser, G.; Bertsch, C.; Röderstein, M.; Hertel, D. Large altitudinal increase in tree root/shoot ratio in tropical mountain forests of Ecuador. Basic Appl. Ecol. 2007, 8, 219–230. [Google Scholar] [CrossRef]
- Ensslin, A.; Rutten, G.; Pommer, U.; Zimmermann, R.; Hemp, A.; Fischer, M. Effects of elevation and land use on the biomass of trees, shrubs and herbs at Mount Kilimanjaro. Ecosphere 2015, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Khadanga, S.S.; Jayakumar, S. Tree biomass and carbon stock: Understanding the role of species richness, elevation, and disturbance. Trop. Ecol. 2020, 61, 128–141. [Google Scholar] [CrossRef]
- Måren, I.E.; Karki, S.; Prajapati, C.; Yadav, R.K.; Shrestha, B.B. Facing north or south: Does slope aspect impact forest stand characteristics and soil properties in a semiarid trans-Himalayan valley? J. Arid Environ. 2015, 121, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Xue, R.; Yang, Q.; Miao, F.; Wang, X.; Shen, Y.; Xue, R.; Yang, Q.; Miao, F.; Wang, X.; Shen, Y. Slope aspect influences plant biomass, soil properties and microbial composition in alpine meadow on the Qinghai-Tibetan plateau. J. Soil Sci. Plant Nutr. 2018, 18, 1–12. [Google Scholar] [CrossRef]
- den Dubbelden, K.C.; Knops, J.M.H. The Effect of Competition and Slope Inclination on Aboveground Biomass Allocation of Understorey Ferns in Subtropical Forest. Oikos 1993, 67, 285. [Google Scholar] [CrossRef]
- Duduman, G.; Duduman, M.-L.; Avăcăriței, D.; Barnoaiea, I.; Barbu, C.-O.; Ciornei, I.; Clinovschi, F.; Coșofreț, V.C.; Cotos, M.-G.; Dănilă, G.; et al. A Permanent Research Platform for Ecological Studies in Intact Temperate Mountainous Forests from Slătioara UNESCO Site and Its Surroundings, Romania. Forests 2020, 11, 1004. [Google Scholar] [CrossRef]
- Doniţã, N.; Paucã-Comãnescu, M.; Popescu, A.; Mihãilescu, S.; Biriş, I.-A. Habitatele din România; Editura Tehnicã Silvicã Bucureşti: Bucharest, Romania, 2005; ISBN 973-96-0-014-X. [Google Scholar]
- European Commission. Interpretation Manual of European Union Habitats—EUR 27; European Commission: Brussels, Belgium, 2007. [Google Scholar]
- Gafta, D.; Mountford, O. Manual de Interpretare a Habitatelor Natura 2000 din România; Gafta, D., Mountford, O., Eds.; Risoprint: Cluj-Napoca, Romania, 2008; ISBN 978-973-751-697-8. [Google Scholar]
- Anonymus. Anonymus. Forest Management Plan of Forest Management Unit VI Chiril. ICAS; Crucea Forest District, Suceava State Forest Administration: Bistrita, Romania, 2010. [Google Scholar]
- Anonymus. Forest Management Plan of Forest Management Unit I Rarău. ICAS; Pojorâta Forest District, Suceava State Forest Administration: Bistrita, Romania, 2014. [Google Scholar]
- Anonymus. Forest Management Plan of Forest Management Unit VIII Slătioara. NCDS „Marin Drăcea”; Stulpicani Forest District, Suceava State Forest Administration: Bistrita, Romania, 2016. [Google Scholar]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423, 623–656. [Google Scholar] [CrossRef] [Green Version]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Lexerød, N.L.; Eid, T. An evaluation of different diameter diversity indices based on criteria related to forest management planning. For. Ecol. Manag. 2006, 222, 17–28. [Google Scholar] [CrossRef]
- O’Hara, K.L.; Hasenauer, H.; Kindermann, G. Sustainability in multi-aged stands: An analysis of long-term plenter systems. Forestry 2007, 80, 163–181. [Google Scholar] [CrossRef] [Green Version]
- Duduman, G. A forest management planning tool to create highly diverse uneven-aged stands. Forestry 2011, 84, 301–314. [Google Scholar] [CrossRef]
- Gini, C. Variabilità e mutabilità. In Memorie di Metodologica Statistica; Pizetti, E.S.T., Ed.; Libreria Eredi Virgilio Veschi: Rome, Italy, 1995. [Google Scholar]
- Gini, C. Measurement of Inequality of Incomes. Econ. J. 1921, 31, 124. [Google Scholar] [CrossRef]
- Lorenz, M.O. Methods of Measuring the Concentration of Wealth. Publ. Am. Stat. Assoc. 1905, 9, 209. [Google Scholar] [CrossRef]
- Valbuena, R.; Packalen, P.; Mehtätalo, L.; García-Abril, A.; Maltamo, M. Characterizing forest structural types and shelterwood dynamics from Lorenz-based indicators predicted by airborne laser scanning. Can. J. For. Res. 2013, 43, 1063–1074. [Google Scholar] [CrossRef]
- Duduman, G. An Ecological Approach for Establishing the Allowable Cut in Forests Where Single Tree Selection System is Applied; Editura Universităţii Suceava: Suceava, Romania, 2009. (In Romanian) [Google Scholar]
- Baker, J.B.; Cain, M.D.; Guldin, J.M.; Murphy, P.A.; Shelton, M.G. Uneven-Aged Silviculture for the Loblolly and Shortleaf Pine Forest Cover Types; General Technical Report SO-1 18; USDA: Washington, DC, USA, 1996. [Google Scholar]
- Giurgiu, V.; Drăghiciu, D. Metode și Tabele Dendrometrice; Ceres Publishing House: Bucharest, Romania, 2004. [Google Scholar]
- Giurgiu, V.; Decei, I.; Drăghiciu, D. Modele Matematico-Auxologice şi Tabele de Producţie Pentru Arborete; Ceres Publishing House: Bucharest, Romania, 2004. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson Prentice Hall: Hoboken, NJ, USA, 2010. [Google Scholar]
- A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013.
- Kobler, J.; Zehetgruber, B.; Dirnböck, T.; Jandl, R.; Mirtl, M.; Schindlbacher, A. Effects of aspect and altitude on carbon cycling processes in a temperate mountain forest catchment. Landsc. Ecol. 2019, 34, 325–340. [Google Scholar] [CrossRef] [Green Version]
- Potter, K.M.; Woodall, C.W. Does biodiversity make a difference? Relationships between species richness, evolutionary diversity, and aboveground live tree biomass across U.S. forests. For. Ecol. Manag. 2014, 321, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Sabatini, F.M.; Zanini, M.; Dowgiallo, G.; Burrascano, S. Multiscale heterogeneity of topsoil properties in southern European old-growth forests. Eur. J. For. Res. 2015, 134, 911–925. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Penttinen, A.; Leinonen, K.; Nygren, M. Statistical opportunities for comparing stand structural heterogeneity in managed and primeval forests: An example from boreal spruce forest in southern Finland. Silva Fenn. 1996, 30, 315–328. [Google Scholar] [CrossRef] [Green Version]
- Schütz, J.P. Close-to-nature silviculture: Is this concept compatible with species diversity? Forestry 1999, 72, 359–366. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H. Individual size inequality links forest diversity and above-ground biomass. J. Ecol. 2015, 103, 1245–1252. [Google Scholar] [CrossRef]
- Fotis, A.T.; Murphy, S.J.; Ricart, R.D.; Krishnadas, M.; Whitacre, J.; Wenzel, J.W.; Queenborough, S.A.; Comita, L.S. Above-ground biomass is driven by mass-ratio effects and stand structural attributes in a temperate deciduous forest. J. Ecol. 2018, 106, 561–570. [Google Scholar] [CrossRef]
- Krupková, L.; Marková, I.; Havránková, K.; Pokorný, R.; Urban, O.; Šigut, L.; Pavelka, M.; Cienciala, E.; Marek, M.V. Comparison of different approaches of radiation use efficiency of biomass formation estimation in Mountain Norway spruce. Trees 2017, 31, 325–337. [Google Scholar] [CrossRef]
- Salinas-Melgoza, M.A.; Skutsch, M.; Lovett, J.C. Predicting aboveground forest biomass with topographic variables in human-impacted tropical dry forest landscapes. Ecosphere 2018, 9, e02063. [Google Scholar] [CrossRef]
- Lieberman, D.; Lieberman, M.; Peralta, R.; Hartshorn, G.S. Tropical Forest Structure and Composition on a Large-Scale Altitudinal Gradient in Costa Rica. J. Ecol. 1996, 84, 137–152. [Google Scholar] [CrossRef]
- Marshall, A.R.; Willcock, S.; Platts, P.J.; Lovett, J.C.; Balmford, A.; Burgess, N.D.; Latham, J.E.; Munishi, P.K.T.; Salter, R.; Shirima, D.D.; et al. Measuring and modelling above-ground carbon and tree allometry along a tropical elevation gradient. Biol. Conserv. 2012, 154, 20–33. [Google Scholar] [CrossRef]
- Day, M.; Baldauf, C.; Rutishauser, E.; Sunderland, T.C.H. Relationships between tree species diversity and above-ground biomass in Central African rainforests: Implications for REDD. Environ. Conserv. 2014, 41, 64–72. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species (i)/Code | Coefficients for Equation (7) | WID-j (t·m−3) | ||||
|---|---|---|---|---|---|---|
| a0 | a1 | a2 | a3 | a4 | ||
| Acer pseudoplatanus L. /AP | −4.06012 | 1.81478 | 0.07283 | 0.76688 | 0.006155 | 0.530 |
| Abies alba Mill. /AA | −4.46414 | 2.19479 | −0.12498 | 1.04645 | −0.016848 | 0.335 |
| Betula pendula Roth /BP | −4.16999 | 2.27038 | −0.21540 | 0.30765 | 0.368258 | 0.530 |
| Fagus sylvatica L. /FS | −4.11122 | 1.30216 | 0.23636 | 1.26562 | −0.079661 | 0.545 |
| Larix decidua Mill. /LD | −4.59667 | 2.26066 | −0.13256 | 1.02582 | 0.007491 | 0.460 |
| Populus tremula L. /PT | −4.22131 | 1.76256 | 0.05900 | 1.04105 | −0.009430 | 0.390 |
| Picea abies (L.) H. Karst./PA | −4.18161 | 2.08131 | −0.11819 | 0.70119 | 0.148181 | 0.353 |
| Pinus sylvestris L. /PS | −3.84672 | 1.82103 | −0.04107 | 0.35677 | 0.334910 | 0.406 |
| Sorbus aucuparia L. /SA | −4.31485 | 2.58064 | −0.21693 | 0.55092 | 0.025773 | 0.530 |
| Taxus baccata L. /TB | −4.46414 | 2.19479 | −0.12498 | 1.04645 | −0.016848 | 0.460 |
| Ulmus glabra Huds. /UG | −4.49118 | 2.18244 | −0.10324 | 1.20293 | −0.124978 | 0.530 |
| Tree Species Code | No. of Plots Per Habitat Types | Living Trees Per Habitat Types | Mean AgB Per SP and Habitat Types (Tonnes) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 9110 | 91V0 | 9410 | NN | Total Area | 9110 | 91V0 | 9410 | NN | Total Area | 9110 | 91V0 | 9410 | NN | Total Area | |
| AP | 7 | 36 | 7 | 4 | 54 | 14 | 102 | 12 | 12 | 140 | 0.49 | 0.52 | 1.44 | 0.45 | 0.63 |
| AA | 35 | 99 | 20 | 8 | 162 | 792 | 1755 | 415 | 110 | 3072 | 4.20 | 4.56 | 3.39 | 6.23 | 4.42 |
| BP | 2 | 2 | 3 | - | 7 | 3 | 2 | 16 | - | 21 | 0.06 | 0.17 | 0.22 | - | 0.16 |
| FS | 34 | 100 | 16 | 6 | 156 | 498 | 1586 | 107 | 24 | 2215 | 4.46 | 5.78 | 1.06 | 1.37 | 4.84 |
| LD | - | - | - | 1 | 1 | - | - | - | 10 | 10 | - | - | - | 6.05 | 6.05 |
| PT | 1 | 1 | - | 1 | 3 | 1 | 1 | - | - | 3 | 0.08 | 0.77 | - | 0.10 | 0.32 |
| PA | 35 | 95 | 42 | 15 | 187 | 327 | 688 | 1325 | 458 | 2798 | 5.17 | 3.59 | 7.75 | 7.96 | 5.17 |
| PS | - | - | - | 1 | 1 | - | - | - | 9 | 9 | - | - | - | 3.59 | 3.59 |
| SA | - | - | 5 | 1 | 6 | - | - | 9 | 4 | 13 | - | - | 0.07 | 0.13 | 0.08 |
| TB | 1 | 3 | - | - | 4 | 1 | 3 | - | - | 4 | 0.004 | 0.03 | - | - | 0.02 |
| UG | - | 8 | - | - | 8 | - | 11 | - | - | 11 | - | 0.28 | - | - | 0.28 |
| Total | 35 | 101 | 42 | 15 | 193 | 1636 | 4148 | 1884 | 628 | 8296 | 13.81 | 13.79 | 10.03 | 12.61 | 12.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duduman, G.; Barnoaiea, I.; Avăcăriței, D.; Barbu, C.-O.; Coșofreț, V.-C.; Dănilă, I.-C.; Duduman, M.-L.; Măciucă, A.; Drăgoi, M. Aboveground Biomass of Living Trees Depends on Topographic Conditions and Tree Diversity in Temperate Montane Forests from the Slătioara-Rarău Area (Romania). Forests 2021, 12, 1507. https://doi.org/10.3390/f12111507
Duduman G, Barnoaiea I, Avăcăriței D, Barbu C-O, Coșofreț V-C, Dănilă I-C, Duduman M-L, Măciucă A, Drăgoi M. Aboveground Biomass of Living Trees Depends on Topographic Conditions and Tree Diversity in Temperate Montane Forests from the Slătioara-Rarău Area (Romania). Forests. 2021; 12(11):1507. https://doi.org/10.3390/f12111507
Chicago/Turabian StyleDuduman, Gabriel, Ionuț Barnoaiea, Daniel Avăcăriței, Cătălina-Oana Barbu, Vasile-Cosmin Coșofreț, Iulian-Constantin Dănilă, Mihai-Leonard Duduman, Anca Măciucă, and Marian Drăgoi. 2021. "Aboveground Biomass of Living Trees Depends on Topographic Conditions and Tree Diversity in Temperate Montane Forests from the Slătioara-Rarău Area (Romania)" Forests 12, no. 11: 1507. https://doi.org/10.3390/f12111507