
Early Growth Responses of Larix kaempferi (Lamb.) Carr. Seedling to Short-Term Extreme Climate Events in Summer


Abstract
:1. Introduction
2. Materials and Methods
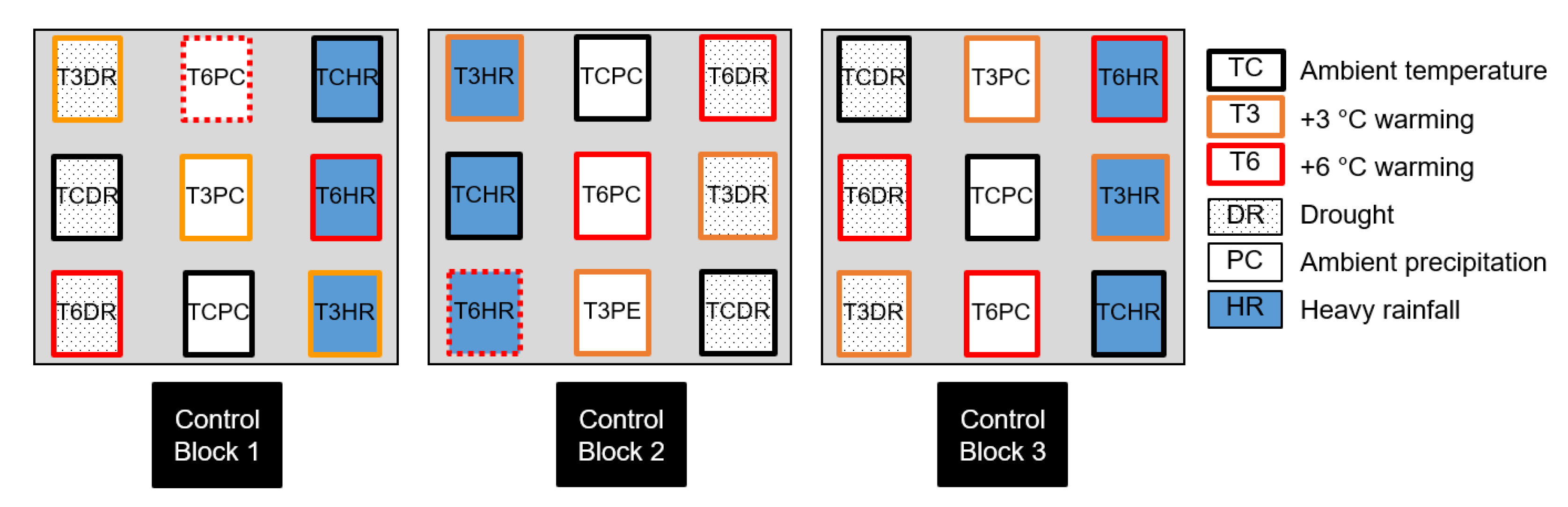
2.1. Experimental Design
2.2. Seedling Measurements
2.3. Data Analyses
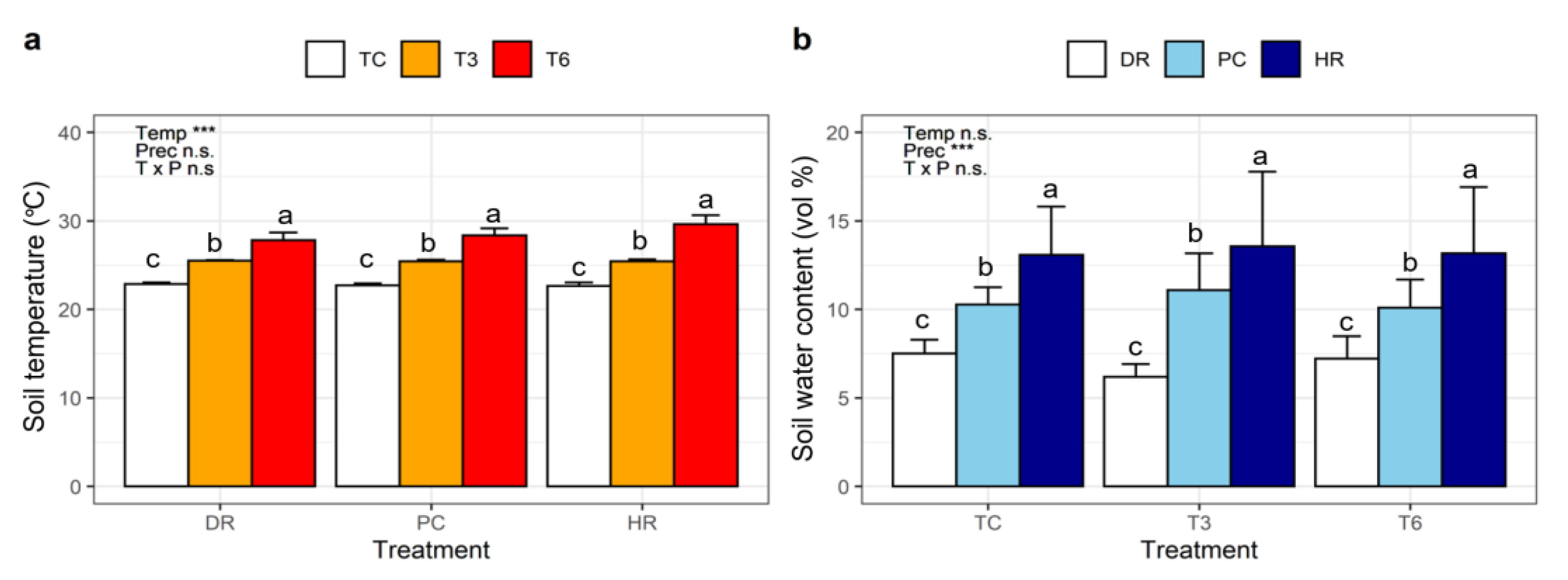
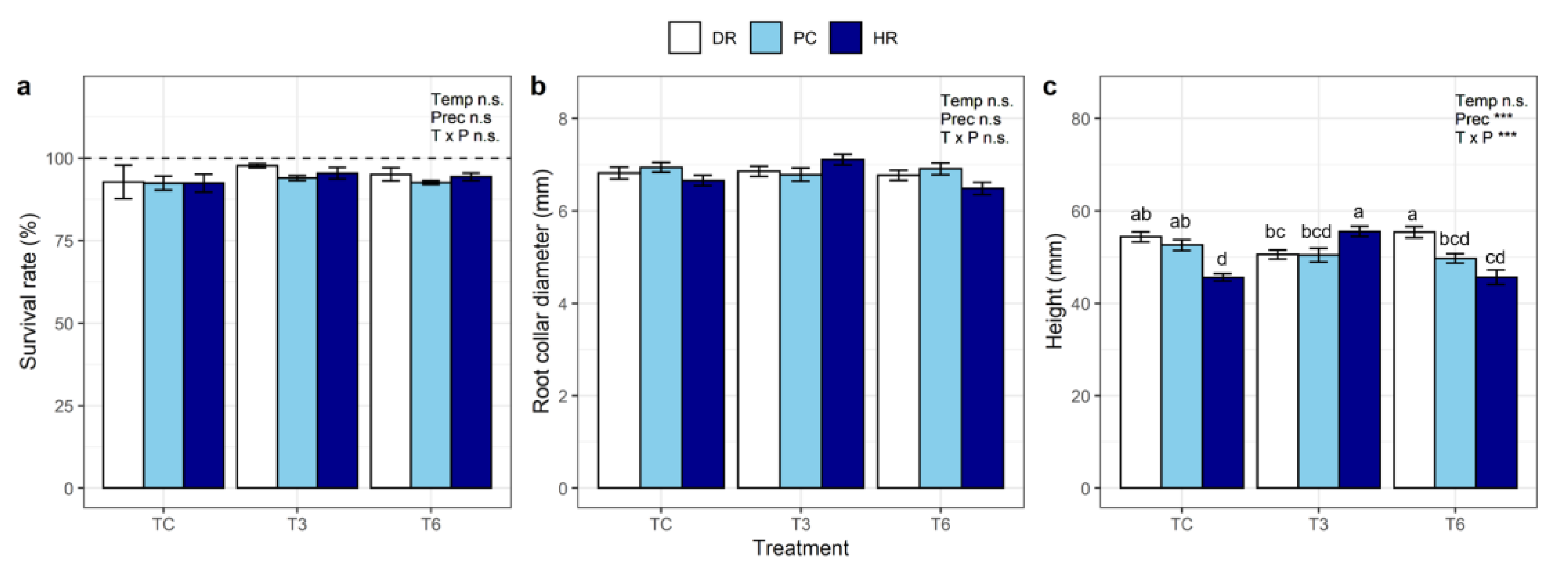
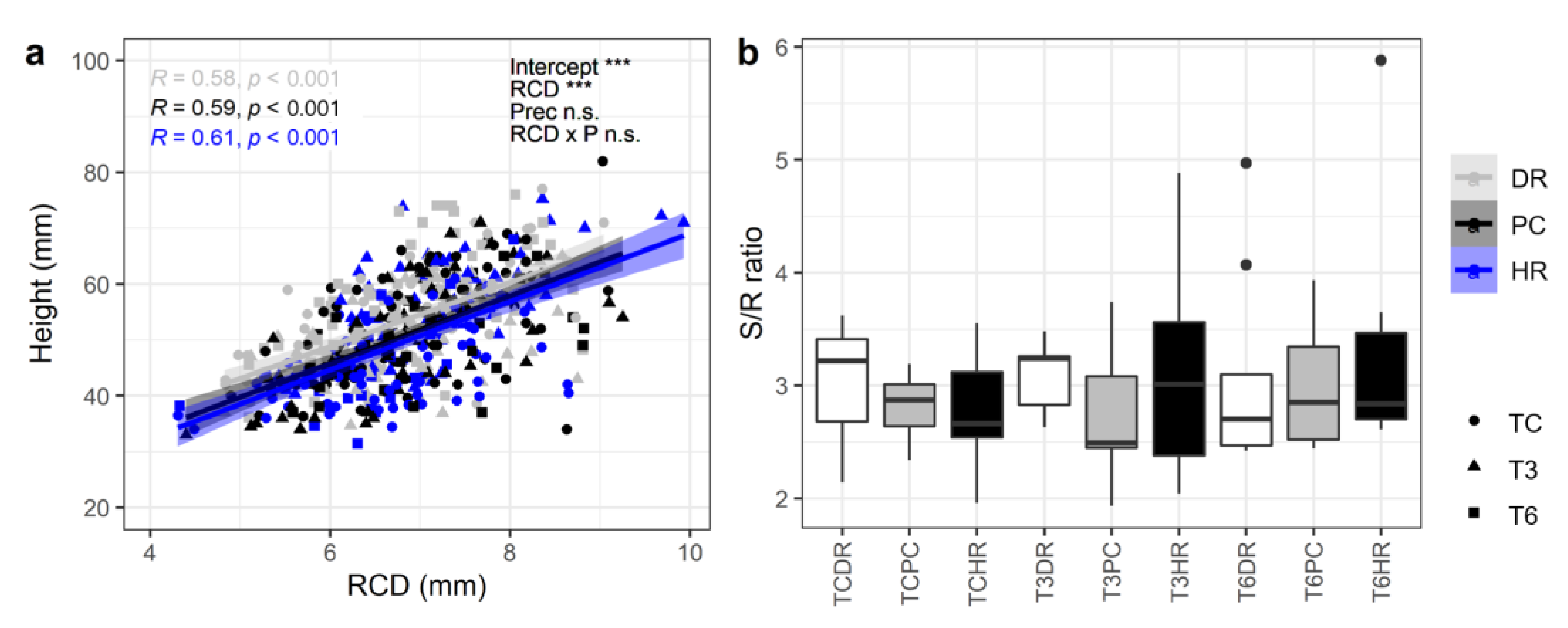
3. Results
3.1. Effects of Temperature and Precipitation Treatments on Seedling Development
3.2. S/R Ratios and SQI
4. Discussion
4.1. Survival
4.2. Plant Growth
4.3. Biomass Allocation and Seedling Quality
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate change 2013: The physical science basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Ed.; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Mazdiyasni, O.; AghaKouchak, A. Substantial increase in concurrent droughts and heatwaves in the United States. Proc. Natl. Acad. Sci. USA 2015, 37, 11484–11489. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Jiang, D.; Lang, X. Future extreme climate changes linked to global warming Intensity. Sci. Bull. 2017, 62, 1673–1680. [Google Scholar] [CrossRef] [Green Version]
- Orlowsky, B.; Seneviratne, S.I. Global changes in extreme events: Regional and seasonal Dimension. Clim. Chang. 2012, 110, 669–696. [Google Scholar] [CrossRef] [Green Version]
- Myhre, G.; Alterskjær, K.; Stjern, C.W.; Hodnebrog, Ø.; Marelle, L.; Samset, B.H.; Sillmann, J.; Schaller, N.; Fischer, E.; Schulz, M.; et al. Frequency of extreme precipitation increases extensively with event rareness under global warming. Sci. Rep. 2019, 9, 16063. [Google Scholar] [CrossRef] [Green Version]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.; Luo, Y.; Li, D.; Cao, S.; Xia, J.; Li, J.; Smith, M.D. Plant growth and mortality under climate extremes: An overview. Environ. Exp. Bot. 2014, 98, 13–19. [Google Scholar] [CrossRef]
- Reyer, C.P.O.; Leuzinger, S.; Rammig, A.; Wolf, A.; Bartholomeus, R.P.; Bonfante, A.; de Lorenzi, F.; Dury, M.; Gloning, P.; Abou Jaoude, R.; et al. A plant’s perspective of extremes: Terrestrial plant responses to changing climatic variability. Glob. Chang. Biol. 2013, 19, 75–89. [Google Scholar] [CrossRef]
- Fisichelli, N.; Wright, A.; Rice, K.; Mau, A.; Buschena, C.; Reich, P.B. First-year seedlings and climate change: Species-specific responses of 15 North American tree species. Oikos 2014, 123, 1331–1340. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Manzanedo, R.D.; D’Orangeville, L.; Lv, P.; Wang, C.; Xu, C.; Hou, M.; Huang, X.; Rademacher, T. High risk of growth cessation of planted larch under extreme drought. Environ. Res. Lett. 2021, 16, 014040. [Google Scholar] [CrossRef]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef]
- Ruehr, N.K.; Grote, R.; Mayr, S.; Arneth, A. Beyond the extreme: Recovery of carbon and water relations in woody plants following heat and drought stress. Tree Physiol. 2019, 39, 1285–1299. [Google Scholar] [CrossRef] [PubMed]
- Bauweraerts, I.; Wertin, T.M.; Ameye, M.; McGuire, M.A.; Teskey, R.O.; Steppe, K. The Effect of heat waves, elevated [CO2] and low soil water availability on northern red oak (Quercus rubra L.) seedlings. Glob. Change. Biol. 2013, 19, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Kuster, T.M.; Arend, M.; Bleuler, P.; Günthardt-Goerg, M.S.; Schulin, R. Water regime and growth of young oak stands subjected to air-warming and drought on two different forest soils in a model ecosystem experiment. Plant Biol. 2012, 15, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Kreyling, J.; Beier, C. Complexity in climate change manipulation experiments. Bioscience 2013, 63, 763–767. [Google Scholar] [CrossRef] [Green Version]
- Matías, L.; Castro, J.; Villar-Salvador, P.; Quero, J.L.; Jump, A.S. Differential impact of hotter drought on seedling performance of five ecologically distinct pine species. Plant Ecol. 2017, 218, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Terazawa, K.; Maruyama, Y.; Morikawa, Y. Photosynthetic and stomatal responses of larix kaempferi seedlings to short-term waterlogging. Ecol. Res. 1992, 7, 193–197. [Google Scholar] [CrossRef]
- Tei, S.; Sugimoto, A.; Yonenobu, H.; Kotani, A.; Maximov, T.C. Effects of extreme drought and wet events for tree mortality: Insights from tree-ring width and carbon isotope ratio in a siberian larch forest. Echohydrology 2019, 12, e2143. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the anthropocene. Ecosphere 2015, 6, 129. [Google Scholar] [CrossRef]
- Saccone, P.; Sylvain, D.; Pagés, J.-P.; Brun, J.-J.; Michalet, R. The role of biotic interactions in altering tree seedling responses to an extreme climate event. J. Veg. Sci. 2009, 20, 403–414. [Google Scholar] [CrossRef]
- Andivia, E.; Villar-Salvador, P.; Oliet, J.A.; Puértolas, J.; Dumroese, R.K.; Ivetić, V.; Molina-Venegas, R.; Arellano, E.C.; Li, G.; Ovalle, J.F. Climate and species stress resistance modulate the higher survival of large seedlings in forest restorations worldwide. Ecol. Appl. 2021, 31, e02394. [Google Scholar] [CrossRef] [PubMed]
- Grossnickle, S.C.; MacDonald, J.E. Seedling quality: History, application, and plant attributes. Forests 2018, 9, 283. [Google Scholar] [CrossRef] [Green Version]
- Riikonen, J.; Luoranen, J. Seedling production and the field performance of seedlings. Forests 2018, 9, 740. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.S.; Hwang, J.; Yang, A.; Han, S.; Son, Y. Seed germination and seedling survival rate of pinus densiflora and abies holophylla in open-field experimental warming using the infrared lamp. J. Kor. For. Soc. 2014, 103, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Kwon, B.; Cho, M.S.; Yang, A.-R.; Chang, H.; An, J.; Son, Y. The early growth performances of pinus densiflora and larix kaempferi seedlings under open-field experimental warming and precipitation manipulation. J. Korean Soc. For. Sci. 2020, 109, 31–40. [Google Scholar]
- Corcobado, T.; Cubera, E.; Juarez, E.; Moreno, G.; Solla, A. Drought events determine performance of quercus ilex seedlings and increase their susceptibility to phytophthora cinnamomic. Agiric. For. Meteorol. 2014, 192, 1–8. [Google Scholar]
- Hou, Q.Q.; Chen, B.M.; Peng, S.L.; Chen, L.Y. Effects of extreme temperature on seedling establishment of nonnative invasive plants. Biol. Invasions 2014, 16, 2049–2061. [Google Scholar] [CrossRef]
- Sukhbaatar, G.; Ganbaatar, B.; Jamsran, T.; Purevragchaa, B.; Nachin, B.; Gradel, A. Assessment of early survival and growth of planted scots pine (Pinus sylvestris) seedlings under extreme continental climate conditions of northern mongolia. J. For. Res. 2020, 31, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Takata, K.; Kurinobu, K.; Koisumi, A.; Yasue, K.; Tamai, Y.; Kisanuki, M. Bibliography on Japanese Larch (Larix kaempferi (Lamb.) Carr.). Eurasian J. For. Res. 2005, 8, 111–126. [Google Scholar]
- Korea Forest Service. Statistical Yearbook of Forestry; Korea Forest Service: Daejeon, Korea, 2020. (In Korean) [Google Scholar]
- Dulamsuren, C.; Hauck, M.; Leuschner, C. Recent drought stress leads to growth reductions in Larix sibirica in the Western Khentey, Mongolia. Glob. Change Biol. 2010, 16, 3024–3035. [Google Scholar] [CrossRef]
- Lévesque, M.; Saurer, M.; Siegwolf, R.; Eilmann, B.; Brang, P.; Bugmann, H.; Rigling, A. Drought response of five conifer species under contrasting water availability suggests high vulnerability of Norway Spruce and European larch. Glob. Change Biol. 2013, 19, 3184–3199. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, D.; Shen, J.; Sun, X.; Zhang, S. Estimating the distribution and productivity characters of larix kaempferi in response to climate change. J. Environ. Manag. 2021, 280, 111633. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Chang, H.; Park, M.J.; Han, S.H.; Hwang, J.H.; Cho, M.S.; Son, Y. Effect of experimental warming on physiological and growth responses of larix kaempferi seedlings. J. Clim. Chang. Res. 2016, 7, 77–84. [Google Scholar] [CrossRef]
- Korea Meteorological Administration. Climate Data Open Portal. 2021. Available online: Data.kma.go.kr/cmmn/main.do (accessed on 30 September 2021).
- Korea Forest Service. Guidelines for Seed and Seedlings Management; Korea Forest Service: Daejeon, Korea, 2020. [Google Scholar]
- Alexander, L.V.; Zhang, X.; Peterson, T.C.; Caesar, J.; Gleason, B.; Klein Tank, A.M.G.; Haylock, M.; Collins, D.; Trewin, B.; Rahimzadeh, R.; et al. Global Observed changes in daily climate extremes of temperature and precipitation. J. Geophys. Res. 2006, 111, D05109. [Google Scholar] [CrossRef] [Green Version]
- Grant, K.; Kreyling, J.; Heilmeier, H.; Beierkuhnlein, C.; Jentsch, C. Extreme weather events and plant-plant interactions: Shifts between competition and facilitation among grassland species in the face of drought and heavy rainfall. Ecol. Res. 2014, 29, 991–1001. [Google Scholar] [CrossRef]
- Zakaria, R.; Radi, N.F.A.; Satari, S.Z. Extraction method of extreme rainfall Data. J. Phys. Conf. Ser. 2017, 890, 012154. [Google Scholar] [CrossRef]
- Pendergrass, A.G. What precipitation is extreme? Science 2018, 360, 1072–1073. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.J.; Jo, H.; Kim, H.; Cho, M.S.; Noh, N.J.; Chang, H.; Kim, H.S.; Son, Y. Open-field temperature and precipitation manipulations system design to simulate extreme climate events for plants and soils. For. Sci. Tech. 2021. under review. [Google Scholar]
- Pinto, J.R.; Marshall, J.D.; Dumroese, R.K.; Davis, A.S.; Cobos, D.R. Establishment and growth of container seedlings for reforestation: A function of stocktype and edaphic conditions. For. Ecol. Manag. 2011, 261, 1876–1884. [Google Scholar] [CrossRef]
- Deans, J.; Mason, W.; Cannell, M.; Sharpe, A.; Sheppard, L. Growing regimes for bare-root stock of sitka spruce, douglas-fir and scots pine. 1. morphology at the end of the nursery phase. Forestry 1989, 62, 53–60. [Google Scholar]
- Cho, M.S.; Lee, S.W.; Park, B.B. Effects of fertilization methods on the growth and physiological characteristics of larix kaempferi seedlings in the container nursery system. J. Bio-Environ. Con. 2012, 21, 57–65. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects models. R Package Version 3, 2018, pp 1–137. Available online: http://CRAN.R-project.org/package=nlme (accessed on 1 September 2021).
- R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria, 2018. Available online: http://www.R-project.org (accessed on 1 September 2021).
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate forest trees and stands under severe drought: A review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef] [Green Version]
- Weemstra, M.; Eilmann, B.; Sass-Klaassen, U.G.W.; Sterch, F.J. Summer droughts limit tree growth across 10 temperate species on a productive forest site. For. Ecol. Manag. 2013, 306, 142–149. [Google Scholar] [CrossRef]
- Gazol, A.; Ribas, M.; Gutiérrez, E.; Camarero, J.J. Aleppo pine forests from across Spain show drought-induced growth decline partial recovery. Agric. For. Meteorol. 2017, 232, 186–194. [Google Scholar] [CrossRef]
- South, D.B.; Harris, S.W.; Barnett, J.P.; Hainds, M.J.; Gjerstad, D.H. Effect of container type and seedling size on survival and early height growth of pinus palustris seedlings in Alabama, U.S.A. For. Ecol. Manag. 2005, 204, 385–398. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; El-Kassaby, Y.A. Bareroot versus container stocktypes: A performance comparison. New For. 2016, 47, 1–51. [Google Scholar] [CrossRef]
- Adams, H.D.; Collins, A.D.; Briggs, S.P.; Vennetier, M.; Dickman, L.T.; Sevanto, S.A.; Garcia-Forner, N.; Powers, H.H.; Mcdowell, N.G. Experimental drought and heat can delay phenological development and reduce foliar and shoot growth in semiarid trees. Glob. Chang. Biol. 2015, 21, 4210–4220. [Google Scholar] [CrossRef] [PubMed]
- Bigler, C.; Gavin, D.G.; Gunning, C.; Veblen, T.T. Drought induces lagged tree mortality in a subalpine forest in the rocky mountains. Oikos 2007, 116, 1983–1994. [Google Scholar] [CrossRef]
- Hoeppner, S.S.; Dukes, J.S. Interactive responses of open-field plant growth and composition to warming and precipitation. Glob. Chang. Biol. 2012, 18, 1754–1786. [Google Scholar] [CrossRef]
- Taeger, S.; Spark, T.H.; Menzel, A. Effects of temperature and drought manipulations on seedlings of scots pine provenances. Plant Biol. 2015, 17, 361–372. [Google Scholar] [CrossRef]
- Harter, D.E.; Nagy, L.; Backhaus, S.; Beierkuhnlein, C.; Fuss, B.; Huber, G.; Jentsch, A.; Konnert, M.; Thiel, D.; Kreyling, J.A. Comparison of genetic diversity and phenotypic plasticity among european beech (Fagus sylvatica L.) populations from Bulgaria and Germany under drought and temperature manipulation. Int. J. Plant Sci. 2015, 176, 232–244. [Google Scholar] [CrossRef]
- Noh, N.J.; Lee, S.J.; Jo, W.; Han, S.; Yoon, T.K.; Chung, H.; Muraoka, H.; Son, Y. Effects of experimental warming on soil respiration and biomass in Quercus variabilis blume and Pinus densiflora zieb. and zucc. seedling. Ann. For. Sic. 2016, 73, 533–545. [Google Scholar] [CrossRef] [Green Version]
- Rustad, L.E.; Campbell, J.L.; Marion, G.M.; Norby, R.J.; Mitchell, M.J.; Hartley, A.E.; Cornelissen, J.H.C.; Gurevitch, J. A Meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 2001, 126, 543–562. [Google Scholar] [CrossRef]
- Lu, M.; Zhou, X.; Yang, Q.; Li, H.; Luo, Y.; Fang, C.; Chen, J.; Yang, X.; Li, B. Responses of ecosystem carbon cycle to experimental warming: A meta-analysis. Ecology 2013, 94, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogee, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2003, 437, 529–533. [Google Scholar] [CrossRef]
- Voesenek, L.A.C.J.; Colmer, T.D.; Pierik, R.; Millenaar, F.F.; Peeters, A.J.M. How plants cope with complete submergence. N. Phytol. 2006, 170, 213–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, H.; Chang, H.; An, J.; Cho, M.S.; Son, Y. Species specific physiological responses of pinus densiflora and larix kaempferi seedlings to open-field experimental warming and precipitation manipulation. For. Sci. Tech. 2019, 15, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Vitasse, Y.; Bottero, A.; Cailleret, M.; Bigler, C.; Fonti, P.; Gessler, A.; Levesque, M.; Rohner, B.; Weber, P.; Rigling, A.; et al. Contrasting resistance and resilience to extreme drought and late spring frost in five major European tree species. Glob. Chang Biol. 2019, 25, 3781–3792. [Google Scholar] [CrossRef] [PubMed]
- Matisons, R.; Krišāns, O.; Jansons, Ā.; Kondratovičs, T.; Elferts, D.; Ievinsh, G. Norway spruce seedlings from an Eastern Baltic provenance show tolerance to simulated drought. Forests 2021, 12, 82. [Google Scholar] [CrossRef]
- Way, D.A.; Oren, R. Differential Responses to Changes in Growth Temperature between trees from different functional groups and biomes: A review and synthesis of data. Tree Physiol. 2010, 30, 669–688. [Google Scholar] [CrossRef] [Green Version]
- Arend, M.; Kuster, T.; Günthardt-Goerg, M.S.; Dobbertin, M. Provenance-specific growth responses to drought and air warming in three european oak species (Quercus robur, Q. pertaea and Q. pubescens). Tree Physiol. 2011, 31, 287–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noh, N.J.; Crous, K.Y.; Li, J.; Choury, Z.; Barton, C.V.M.; Arndt, S.K.; Reich, P.B.; Tjoelker, M.G.; Pendall, E. Does root respiration in australian rainforest tree seedlings acclimate to experimental warming? Tree Physiol. 2020, 40, 1192–1204. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity. Acta Physiol. Plant 2015, 37, 9. [Google Scholar] [CrossRef]
- Aaltonen, H.; Lindén, A.; Heinonsalo, J.; Biasi, C.; Pumpanen, J. Effect of prolonged drought stress on scots pine seedling carbon allocation. Tree Physiol. 2016, 37, 418–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Oravec, M.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; Parella, T.; Peñuelas, J. Warming differentially influences the effects of drought on stoichiometry and metabolomics in shoots and roots. New Phytol. 2015, 207, 591–603. [Google Scholar] [CrossRef]
- Granda, E.; Camarero, J.J.; Gimeno, T.E.; Martínez-Fernández, J.; Valladares, F. Intensity and timing of warming and drought differentially affect growth patterns of co-occuring mediterranean tree species. Eur. J. For. Res. 2013, 132, 469–480. [Google Scholar] [CrossRef]
- Grossiord, C.; Sevanto, S.; Dawson, T.E.; Adams, H.D.; Collins, A.D.; Dickman, L.T.; Newman, B.D.; Stockton, E.A.; McDowell, N.G. Warming combined with more extreme precipitation regimes modifies the water sources used by trees. New Phytol. 2017, 213, 584–596. [Google Scholar] [CrossRef]
- Liu, Y.; Li, P.; Xiao, L.; Wang, W.; Yu, K.; Shi, P. Heterogeneity in short-term allocation of carbon to roots of Pinus tabuliformis seedlings and root respiration under drought stress. Plant Soil 2020, 452, 359–378. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | d.f. 1 | Growth Responses | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Survival | RCD | Height | Volume | H/D | Total Biomass | S/R 2 | SQI 3 | ||
| Temperature (T) | 2 | 0.305 | 0.258 | 0.270 | 0.176 | 0.373 | 0.881 | 0.419 | 0.498 |
| Precipitation (P) | 2 | 0.576 | 0.598 | <0.001 | 0.155 | <0.001 | 0.955 | 0.390 | 0.145 |
| T × P | 4 | 0.967 | 0.035 | <0.001 | <0.001 | <0.001 | 0.785 | 0.788 | 0.562 |
| Treatment | Leaf Biomass (g) | Stem Biomass (g) | Root Biomass (g) | Total Biomass (g) | SQI | |
|---|---|---|---|---|---|---|
| TC | DR | 4.06 ± 0.42 | 7.03 ± 1.27 | 3.76 ± 0.74 | 14.87 ± 2.00 | 1.23 ± 0.23 |
| PC | 4.80 ± 0.90 | 7.17 ± 1.43 | 4.21 ± 1.07 | 15.88 ± 3.25 | 1.37 ± 0.29 | |
| HR | 4.37 ± 0.71 | 6.30 ± 1.08 | 3.90 ± 0.92 | 14.57 ± 2.50 | 1.37 ± 0.29 | |
| T3 | DR | 4.39 ± 1.10 | 7.11 ± 1.82 | 3.74 ± 0.75 | 15.24 ± 3.47 | 1.24 ± 0.20 |
| PC | 4.38 ± 1.28 | 6.88 ± 1.73 | 4.12 ± 0.73 | 15.38 ± 3.35 | 1.34 ± 0.22 | |
| HR | 4.17 ± 1.10 | 7.12 ± 1.46 | 3.84 ± 1.42 | 15.13 ± 3.48 | 1.21 ± 0.32 | |
| T6 | DR | 4.21 ± 0.55 | 6.87 ± 1.38 | 3.84 ± 1.23 | 14.93 ± 2.95 | 1.20 ± 0.29 |
| PC | 5.03 ± 1.33 | 6.64 ± 1.22 | 4.08 ± 1.43 | 15.75 ± 3.78 | 1.42 ± 0.47 | |
| HR | 4.09 ± 0.58 | 6.03 ± 1.05 | 3.12 ± 0.55 | 13.24 ± 1.27 | 1.09 ± 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noh, N.-J.; Kim, G.-J.; Son, Y.; Cho, M.-S. Early Growth Responses of Larix kaempferi (Lamb.) Carr. Seedling to Short-Term Extreme Climate Events in Summer. Forests 2021, 12, 1595. https://doi.org/10.3390/f12111595
Noh N-J, Kim G-J, Son Y, Cho M-S. Early Growth Responses of Larix kaempferi (Lamb.) Carr. Seedling to Short-Term Extreme Climate Events in Summer. Forests. 2021; 12(11):1595. https://doi.org/10.3390/f12111595
Chicago/Turabian StyleNoh, Nam-Jin, Gwang-Jung Kim, Yowhan Son, and Min-Seok Cho. 2021. "Early Growth Responses of Larix kaempferi (Lamb.) Carr. Seedling to Short-Term Extreme Climate Events in Summer" Forests 12, no. 11: 1595. https://doi.org/10.3390/f12111595
APA StyleNoh, N.-J., Kim, G.-J., Son, Y., & Cho, M.-S. (2021). Early Growth Responses of Larix kaempferi (Lamb.) Carr. Seedling to Short-Term Extreme Climate Events in Summer. Forests, 12(11), 1595. https://doi.org/10.3390/f12111595