
Biomass Estimation Models for Six Shrub Species in Hunshandake Sandy Land in Inner Mongolia, Northern China

Abstract
:1. Introduction
2. Materials and Methods
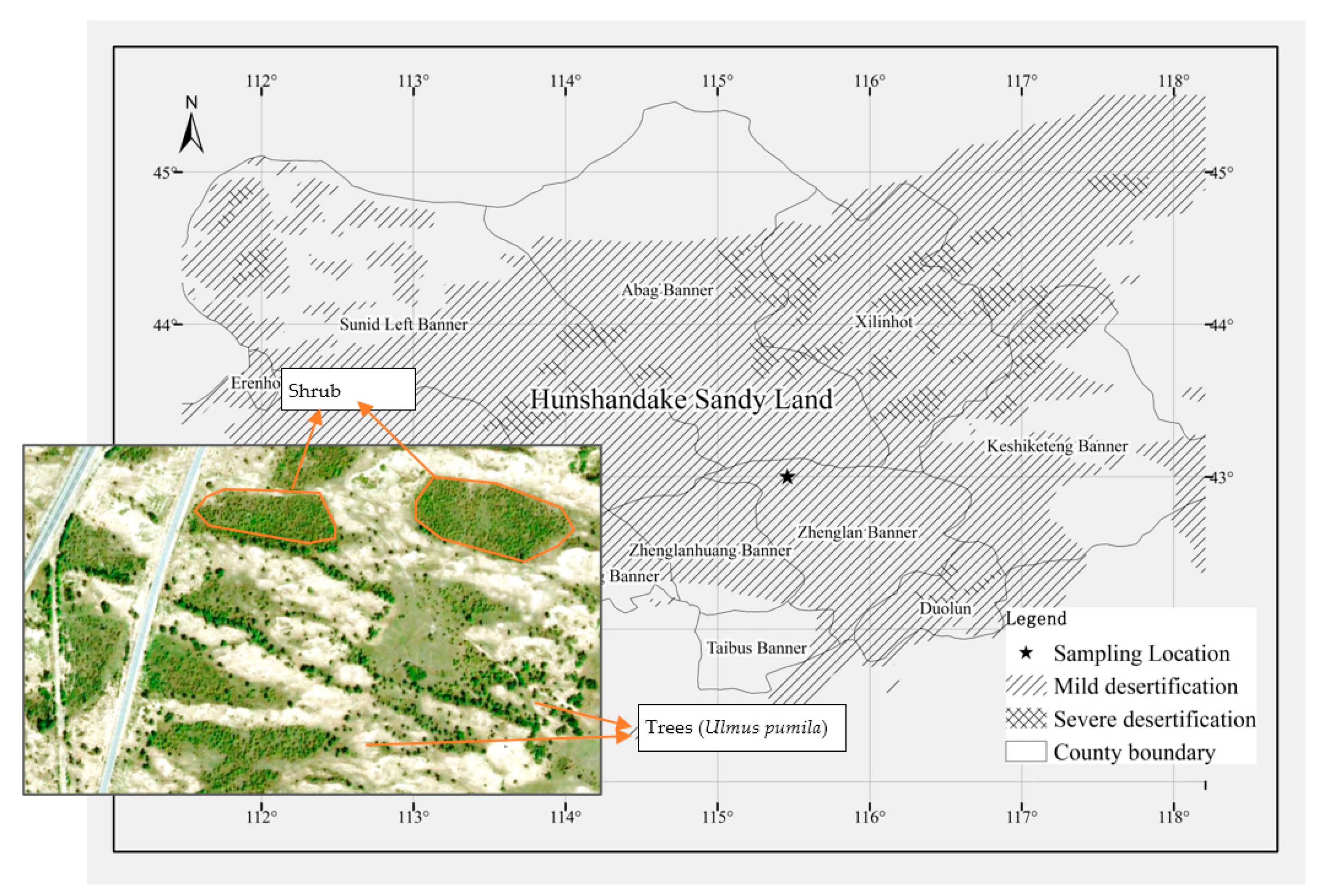
2.1. Study Area
2.2. Above-Ground Biomass Sampling
2.3. The Seven Models for Estimating Above-Ground Biomass of Shrub
2.4. Regression Fitting and Accuracy Evaluation of the Seven Models
3. Results
3.1. Correlation Analysis between Predictors and Shrub Biomass
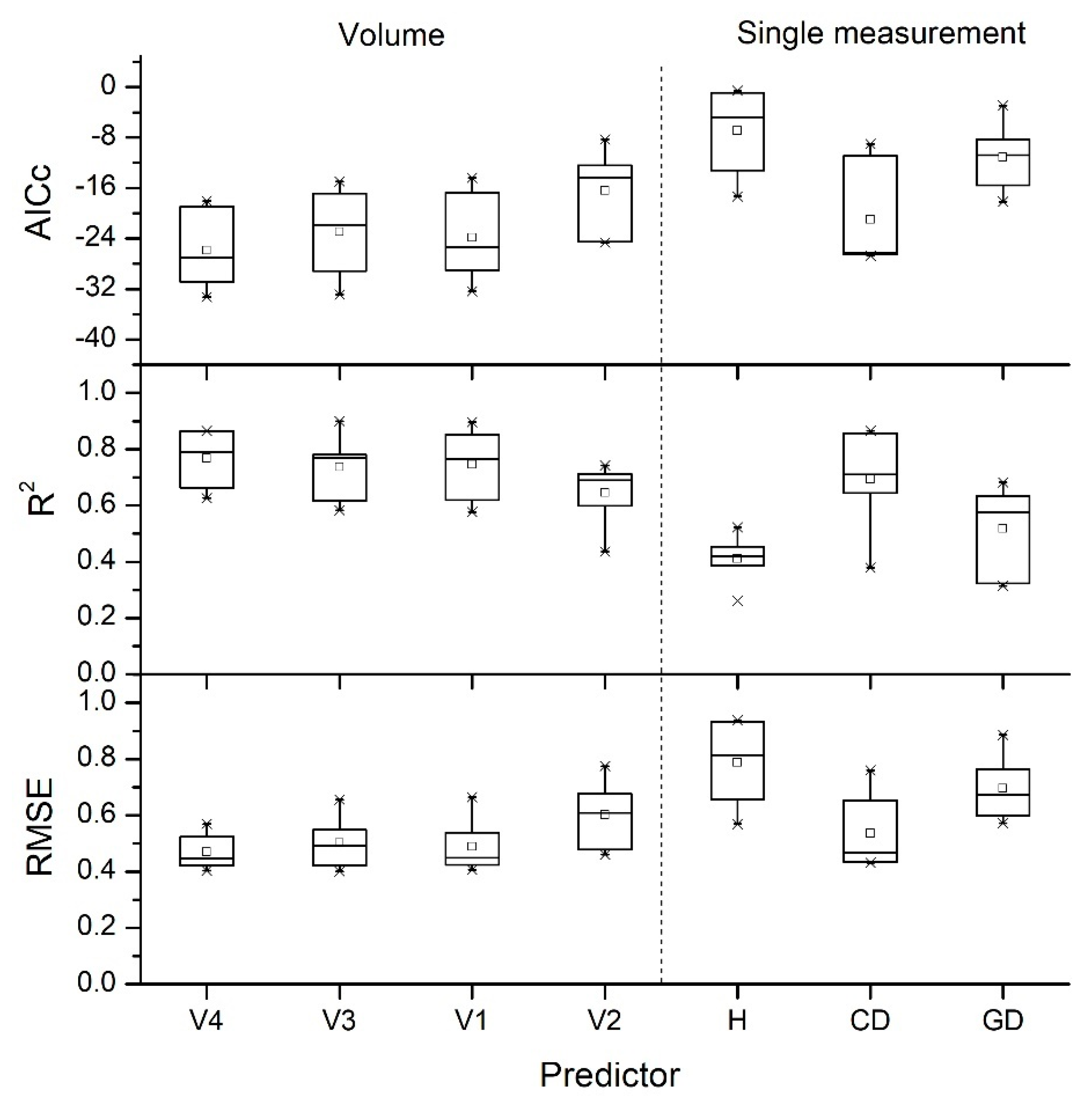
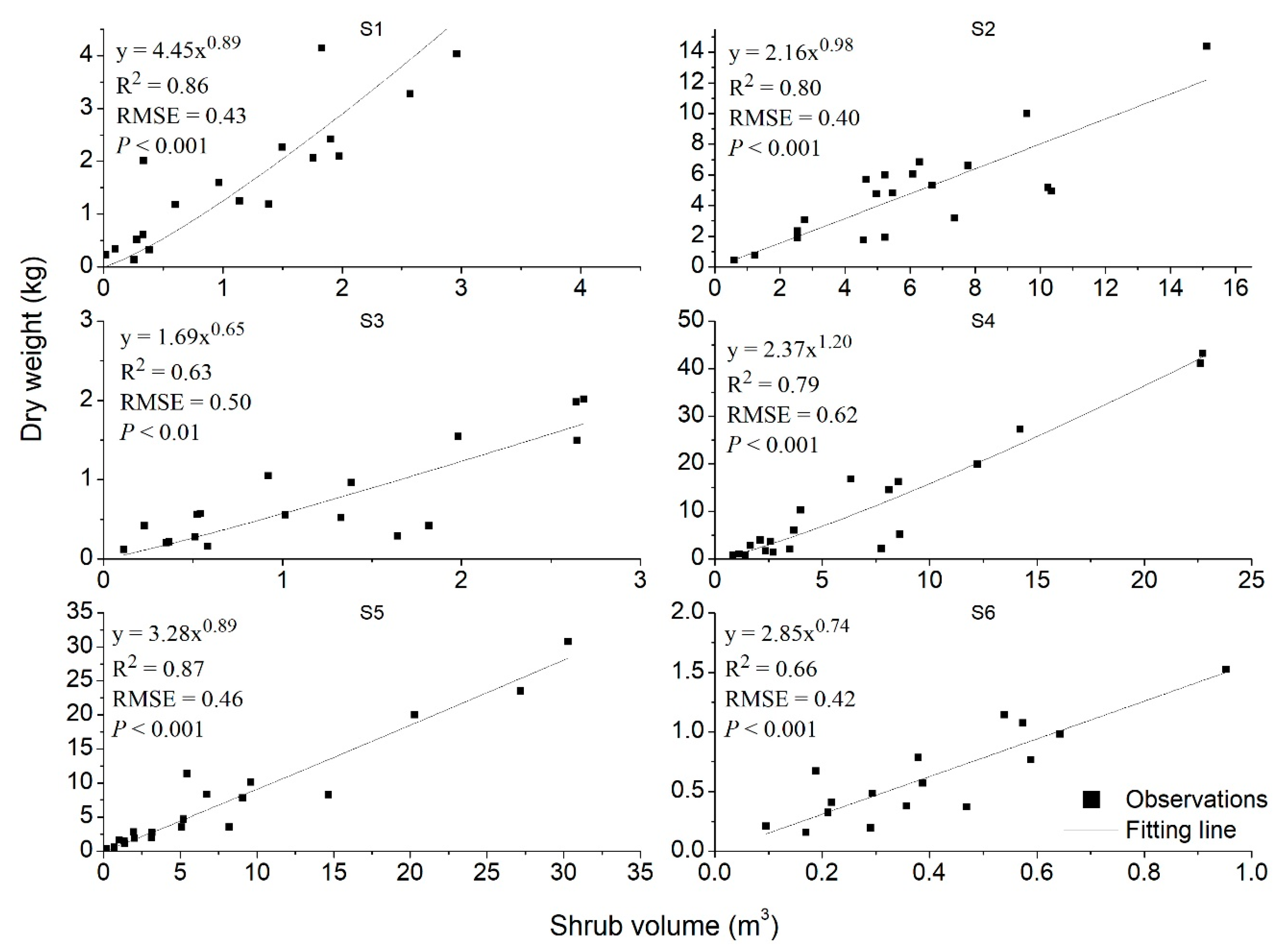
3.2. Fitting Accuracy Evaluation of the Seven Models
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- White, R.P.; Nackoney, J. Drylands, People, and Ecosystem Goods and Services: A Web-Based Geospatial Analysis; World Resources Institute: Washington, DC, USA, 2003. [Google Scholar]
- Hou, X.Y. Vegetation Map of the People’s Republic of China and Its Illustration; China Cartographic Publishing House: Beijing, China, 1982. (In Chinese) [Google Scholar]
- Cerrillo, R.M.N.; Oyonarte, P.B. Estimation of above-ground biomass in shrubland ecosystems of southern Spain. For. Syst. 2006, 15, 197–207. [Google Scholar]
- Daryanto, S.; Eldridge, D.J.; Throop, H.L. Managing semi-arid woodlands for carbon storage: Grazing and shrub effects on above- and belowground carbon. Agric. Ecosyst. Environ. 2013, 169, 1–11. [Google Scholar] [CrossRef]
- Pasalodostato, M.; Ruizpeinado, R.; Rio, M.D.; Montero, G. Shrub biomass accumulation and growth rate models to quantify carbon stocks and fluxes for the Mediterranean region. Eur. J. For. Res. 2015, 134, 537–553. [Google Scholar] [CrossRef]
- Ruizpeinado, R.; Moreno, G.; Juarez, E.; Montero, G.; Roig, S. The contribution of two common shrub species to above-ground and belowground carbon stock in Iberian dehesas. J. Arid. Environ. 2013, 91, 22–30. [Google Scholar] [CrossRef]
- Yang, H.; Wang, Z.; Tan, H.; Gao, Y. Allometric models for estimating shrub biomass in desert grassland in northern China. Arid Land Res. Manag. 2017, 31, 283–300. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, G.; Gao, L.; Niu, S.; Liu, M.; Yu, S.; Peng, Y. Impacts of human disturbance on elms-motte-veldt in Hunshandak sandland. Acta Phytoecol. Sin. 2003, 27, 829–834. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Schaffer, B.E.; Yang, Z.; Rodrigueziturbe, I. Probabilistic model predicts dynamics of vegetation biomass in a desert ecosystem in NW China. Proc. Natl. Acad. Sci. USA 2017, 114, 201703684. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Wang, M.; Ma, M.; Lin, Y. Aboveground Tree Biomass Estimation of Sparse Subalpine Coniferous Forest with UAV Oblique Photography. Remote Sens. 2018, 10, 1849–1868. [Google Scholar] [CrossRef] [Green Version]
- Gabrielle, N.B.; Emma, C.U.; Hugh, D.S. Estimating Biomass in California’s Chaparral and Coastal Sage Scrub Shrublands. Madroño 2018, 65, 28–46. [Google Scholar] [CrossRef]
- Murray, R.B.; Jacobson, M.Q. An Evaluation of Dimension Analysis for Predicting Shrub Biomass. J. Range Manag. 1982, 35, 451. [Google Scholar] [CrossRef]
- Rojo, V.; Arzamendia, Y.; Perez, C.; Baldo, J.L.; Vila, B.L. Double Sampling Methods in Biomass Estimates of Andean Shrubs and Tussocks. Rangel. Ecol. Manag. 2017, 70, 718–722. [Google Scholar] [CrossRef]
- Vora, R.S. Predicting biomass of five shrub species in northeastern California. J. Range Manag. 1988, 41, 63–65. [Google Scholar] [CrossRef]
- Ximenes, F.; Kathuria, A.; Mclean, M.; Coburn, R.; Sargeant, D.; Ryan, M.; Williams, J.; Boer, K.; Mo, M. Carbon in Mature Native Forests in Australia: The Role of Direct Weighing in the Derivation of Allometric Equations. Forests 2018, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, J.A.; Reynolds, J.F.; Whitson, P.D. Size-biomass Relationships of Several Chihuahuan Desert Shrubs. Am. Midl. Nat. 1975, 94, 451–461. [Google Scholar] [CrossRef]
- Mason, N.W.H.; Beets, P.N.; Payton, I.J.; Burrows, L.E.; Holdaway, R.J.; Carswell, F.E. Individual-Based Allometric Equations Accurately Measure Carbon Storage and Sequestration in Shrublands. Forests 2014, 5, 309–324. [Google Scholar] [CrossRef] [Green Version]
- Chave, J.; Rejoumechain, M.; Burquez, A.; Chidumayo, E.N.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C. Improved allometric models to estimate the above-ground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Ganeshamurthy, A.N.; Ravindra, V.; Venugopalan, R.; Mathiazhagan, M.; Bhat, R.M. Biomass Distribution and Development of Allometric Equations for Non-Destructive Estimation of Carbon Sequestration in Grafted Mango Trees. J. Agric. Sci. 2016, 8, 201. [Google Scholar] [CrossRef] [Green Version]
- Ploton, P.; Barbier, N.; Momo, S.T.; Rejoumechain, M.; Bosela, F.B.; Chuyong, G.; Dauby, G.; Droissart, V.; Fayolle, A.; Goodman, R.C. Closing a gap in tropical forest biomass estimation: Taking crown mass variation into account in pantropical allometries. Biogeosciences 2016, 13, 1571–1585. [Google Scholar] [CrossRef] [Green Version]
- Rutishauser, E.; Nooran, F.; Laumonier, Y.; Halperin, J.; Rufiie; Hergoualch, K.; Verchot, L.V. Generic allometric models including height best estimate forest biomass and carbon stocks in Indonesia. For. Ecol. Manag. 2013, 307, 219–225. [Google Scholar] [CrossRef]
- Jagodzinski, A.M.; Dyderski, M.K.; Gesikiewicz, K.; Horodecki, P. Tree- and Stand-Level Biomass Estimation in a Larix decidua Mill. Chronosequence. Forests 2018, 9, 587. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Cui, G.; Shen, W.; Liu, X. Cover as a simple predictor of biomass for two shrubs in Tibet. Ecol. Indic. 2016, 64, 266–271. [Google Scholar] [CrossRef]
- Conti, G.; Enrico, L.; Casanoves, F.; Díaz, S. Shrub biomass estimation in the semi-arid Chaco forest: A contribution to the quantification of an underrated carbon stock. Ann. For. Sci. 2013, 70, 515–524. [Google Scholar] [CrossRef] [Green Version]
- Huff, S.; Ritchie, M.W.; Temesgen, H. Allometric equations for estimating above-ground biomass for common shrubs in northeastern California. For. Ecol. Manag. 2017, 398, 48–63. [Google Scholar] [CrossRef]
- Yi, T.; Arshad, A.; Li, H. Bayesian model predicts the above-ground biomass of Caragana microphylla in sandy lands better than OLS regression models. J. Plant. Ecol. 2020, 13, 732–737. [Google Scholar]
- Picard, N.; Rutishauser, E.; Ploton, P.; Ngomanda, A.; Henry, M. Should tree biomass allometry be restricted to power models. For. Ecol. Manag. 2015, 353, 156–163. [Google Scholar] [CrossRef]
- Uso, J.L.; Mateu, J.; Karjalainen, T.; Salvador, P. Allometric regression equations to determine aerial biomasses of Mediterranean shrubs. Plant Ecol. 1997, 132, 59–69. [Google Scholar] [CrossRef]
- Sternberg, M.; Shoshany, M. Influence of slope aspect on Mediterranean woody formations: Comparison of a semi-arid and an arid site in Israel. Ecol. Res. 2001, 16, 335–345. [Google Scholar] [CrossRef]
- Ma, Y.; Li, G.; Pan, Y.; Ma, Y.; Wang, Q.; Zhang, C. Prediction model for biomass of 3 shrubs in Hunshandake sandy land. J. Arid. Lan. Res. Envi. 2017, 31, 198–201. [Google Scholar]
- Hierro, J.L.; Branch, L.C.; Villarreal, D.; Clark, K.L. Predictive equations for biomass and fuel characteristics of Argentine shrubs. J. Range Manag. 2000, 53, 617–621. [Google Scholar] [CrossRef]
- He, A.; Mcdermid, G.J.; Rahman, M.M.; Strack, M.; Saraswati, S.; Xu, B. Developing Allometric Equations for Estimating Shrub Biomass in a Boreal Fen. Forests 2018, 9, 569. [Google Scholar] [CrossRef] [Green Version]
- Flade, L.; Hopkinson, C.; Chasmer, L. Allometric Equations for Shrub and Short-Stature Tree Aboveground Biomass within Boreal Ecosystems of Northwestern Canada. Forests. 2020, 11, 1207. [Google Scholar] [CrossRef]
- Spiess, A.N.; Neumeyer, N. An evaluation of R2 as an inadequate measure for nonlinear models in pharmacological and biochemical research: A Monte Carlo approach. BMC Pharmacol. 2010, 10, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erik, V.; Kian, W.C. Carbon storage estimation of tropical urban trees by an improved allometric model for above-ground biomass based on terrestrial laser scanning. Urban. For. Urban. Gree. 2019, 44, 126387. [Google Scholar]
- IPCC. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. In Climate Change 2014; Synthesis Report; Core Writing Team, Pachauriand, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151.z. [Google Scholar]
- Brown, J.K. Estimating shrub biomass from basal stem diameters. Can. J. For. Res. 1976, 6, 153–158. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shrub Species | Height Range (cm) | Crown Diameter Range (cm) | Ground Diameter Range (cm) | Dried Biomass Range (g) |
|---|---|---|---|---|
| S1 | 80–190 | 43–250 | 13–107 | 130–7350 |
| S2 | 180–380 | 90–335 | 18–131 | 450–14,380 |
| S3 | 160–330 | 110–440 | 18–192 | 740–43,180 |
| S4 | 55–140 | 60–280 | 17–243 | 120–5180 |
| S5 | 55–170 | 60–215 | 22–84 | 160–1970 |
| S6 | 150–340 | 52–455 | 14–296 | 340–3080 |
| Code | Name | Formula |
|---|---|---|
| H | Height | H = h |
| CD | Crown Diameter | CD = (a1+ b1)/2 |
| GD | Ground Diameter | GD = (a2 +b2)/2 |
| V1 | Circular Cylinder | V1 = πCD2h/4 |
| V2 | Circular Cylinder | V2 = πGD2h/4 |
| V3 | Elliptical Cylinder | V3 = πa1b1h/4 |
| V4 | Inverted Cone Frustum | V4 = 1/12πh(CD2 + CD × GD + GD2) |
| Shrub Species | Predictors | ||||||
|---|---|---|---|---|---|---|---|
| H | CD | GD | V1 | V2 | V3 | V4 | |
| S1 | 0.54 * | 0.83 ** | 0.81 ** | 0.83 ** | 0.81 ** | 0.88 ** | 0.91 ** |
| S2 | 0.48 * | 0.68 ** | 0.47 * | 0.68 ** | 0.63 ** | 0.70 ** | 0.79 ** |
| S3 | 0.74 ** | 0.83 ** | 0.77 ** | 0.92 ** | 0.88 ** | 0.91 ** | 0.95 ** |
| S4 | 0.29 | 0.71 ** | 0.62 ** | 0.72 ** | 0.72 ** | 0.77 ** | 0.87 ** |
| S5 | 0.63 ** | 0.76 ** | 0.51 * | 0.66 ** | 0.71 ** | 0.70 ** | 0.71 ** |
| S6 | 0.63 ** | 0.85 ** | 0.75 ** | 0.92 ** | 0.82 ** | 0.94 ** | 0.89 ** |
| Average | 0.55 | 0.78 | 0.66 | 0.79 | 0.76 | 0.82 | 0.85 |
| Shrub Species | Predictors | a | b | AICc | R2 | RMSE | p-Value |
|---|---|---|---|---|---|---|---|
| S1 | H | 1.00 | 2.84 | −0.86 | 0.39 | 0.92 | < 0.01 |
| CD | 1.00 | 2.06 | −26.47 | 0.87 | 0.43 | <0.001 | |
| GD | 1.00 | 1.65 | −11.88 | 0.68 | 0.66 | <0.001 | |
| V1 | 2.16 | 0.87 | −25.01 | 0.85 | 0.45 | <0.001 | |
| V2 | 225.83 | 0.77 | −15.48 | 0.74 | 0.60 | <0.001 | |
| V3 | 2.50 | 0.71 | −18.23 | 0.78 | 0.55 | <0.001 | |
| V4 | 4.45 | 0.89 | −26.39 | 0.86 | 0.43 | <0.001 | |
| S2 | H | 1.00 | 3.10 | −12.98 | 0.45 | 0.66 | <0.01 |
| CD | 1.00 | 2.37 | −26.20 | 0.74 | 0.46 | <0.001 | |
| GD | 1.02 | 1.30 | −17.95 | 0.59 | 0.57 | <0.001 | |
| V1 | 1.42 | 1.00 | −28.76 | 0.77 | 0.42 | <0.001 | |
| V2 | 37.43 | 0.68 | −24.39 | 0.71 | 0.48 | <0.001 | |
| V3 | 1.43 | 1.00 | −28.89 | 0.77 | 0.42 | <0.001 | |
| V4 | 2.16 | 0.98 | −30.59 | 0.80 | 0.40 | <0.001 | |
| S3 | H | 1.00 | 1.92 | −8.03 | 0.26 | 0.71 | 0.052 |
| CD | 1.00 | 1.25 | −10.64 | 0.38 | 0.65 | <0.05 | |
| GD | 1.02 | 0.72 | −9.17 | 0.31 | 0.69 | <0.05 | |
| V1 | 1.47 | 0.78 | −16.42 | 0.58 | 0.54 | <0.01 | |
| V2 | 1.95 | 0.43 | −12.10 | 0.44 | 0.62 | <0.01 | |
| V3 | 1.47 | 0.78 | −16.57 | 0.58 | 0.54 | <0.01 | |
| V4 | 1.69 | 0.65 | −17.72 | 0.63 | 0.50 | <0.01 | |
| S4 | H | 1.00 | 4.29 | −0.70 | 0.52 | 0.93 | <0.001 |
| CD | 1.00 | 2.71 | −8.83 | 0.68 | 0.76 | <0.001 | |
| GD | 1.01 | 1.60 | −2.69 | 0.57 | 0.89 | <0.001 | |
| V1 | 1.44 | 1.21 | −14.17 | 0.76 | 0.67 | <0.001 | |
| V2 | 172.04 | 0.80 | −8.06 | 0.67 | 0.78 | <0.001 | |
| V3 | 1.44 | 1.22 | −14.75 | 0.76 | 0.66 | <0.001 | |
| V4 | 2.37 | 1.20 | −16.76 | 0.79 | 0.62 | <0.001 | |
| S5 | H | 1.00 | 3.51 | −0.31 | 0.45 | 0.94 | <0.01 |
| CD | 1.00 | 2.12 | −25.91 | 0.86 | 0.48 | <0.001 | |
| GD | 1.01 | 1.33 | −8.09 | 0.63 | 0.76 | <0.001 | |
| V1 | 1.88 | 0.96 | −32.11 | 0.90 | 0.41 | <0.001 | |
| V2 | 24.13 | 0.66 | −12.71 | 0.71 | 0.68 | <0.001 | |
| V3 | 1.91 | 0.96 | −32.63 | 0.90 | 0.40 | <0.001 | |
| V4 | 3.28 | 0.89 | −27.03 | 0.87 | 0.46 | <0.001 | |
| S6 | H | 1.00 | 1.44 | −17.04 | 0.39 | 0.57 | <0.01 |
| CD | 1.00 | 1.92 | −26.25 | 0.64 | 0.43 | <0.001 | |
| GD | 1.00 | 1.36 | −15.33 | 0.32 | 0.60 | <0.05 | |
| V1 | 1.90 | 0.66 | −25.13 | 0.62 | 0.45 | <0.001 | |
| V2 | 18.83 | 0.81 | −24.22 | 0.60 | 0.46 | <0.001 | |
| V3 | 1.91 | 0.65 | −24.95 | 0.62 | 0.45 | <0.001 | |
| V4 | 2.85 | 0.74 | −27.23 | 0.66 | 0.42 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, X.; Yang, G.; Wu, B.; Jiang, L.; Wang, F. Biomass Estimation Models for Six Shrub Species in Hunshandake Sandy Land in Inner Mongolia, Northern China. Forests 2021, 12, 167. https://doi.org/10.3390/f12020167
Yao X, Yang G, Wu B, Jiang L, Wang F. Biomass Estimation Models for Six Shrub Species in Hunshandake Sandy Land in Inner Mongolia, Northern China. Forests. 2021; 12(2):167. https://doi.org/10.3390/f12020167
Chicago/Turabian StyleYao, Xueling, Guojing Yang, Bo Wu, Lina Jiang, and Feng Wang. 2021. "Biomass Estimation Models for Six Shrub Species in Hunshandake Sandy Land in Inner Mongolia, Northern China" Forests 12, no. 2: 167. https://doi.org/10.3390/f12020167