
Aboveground Biomass Allocation of Boreal Shrubs and Short-Stature Trees in Northwestern Canada




Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Plant Destructive Sampling
2.3. In Situ Measurements and Plant Component Aboveground Biomass Allometric Equations
3. Results and Discussion
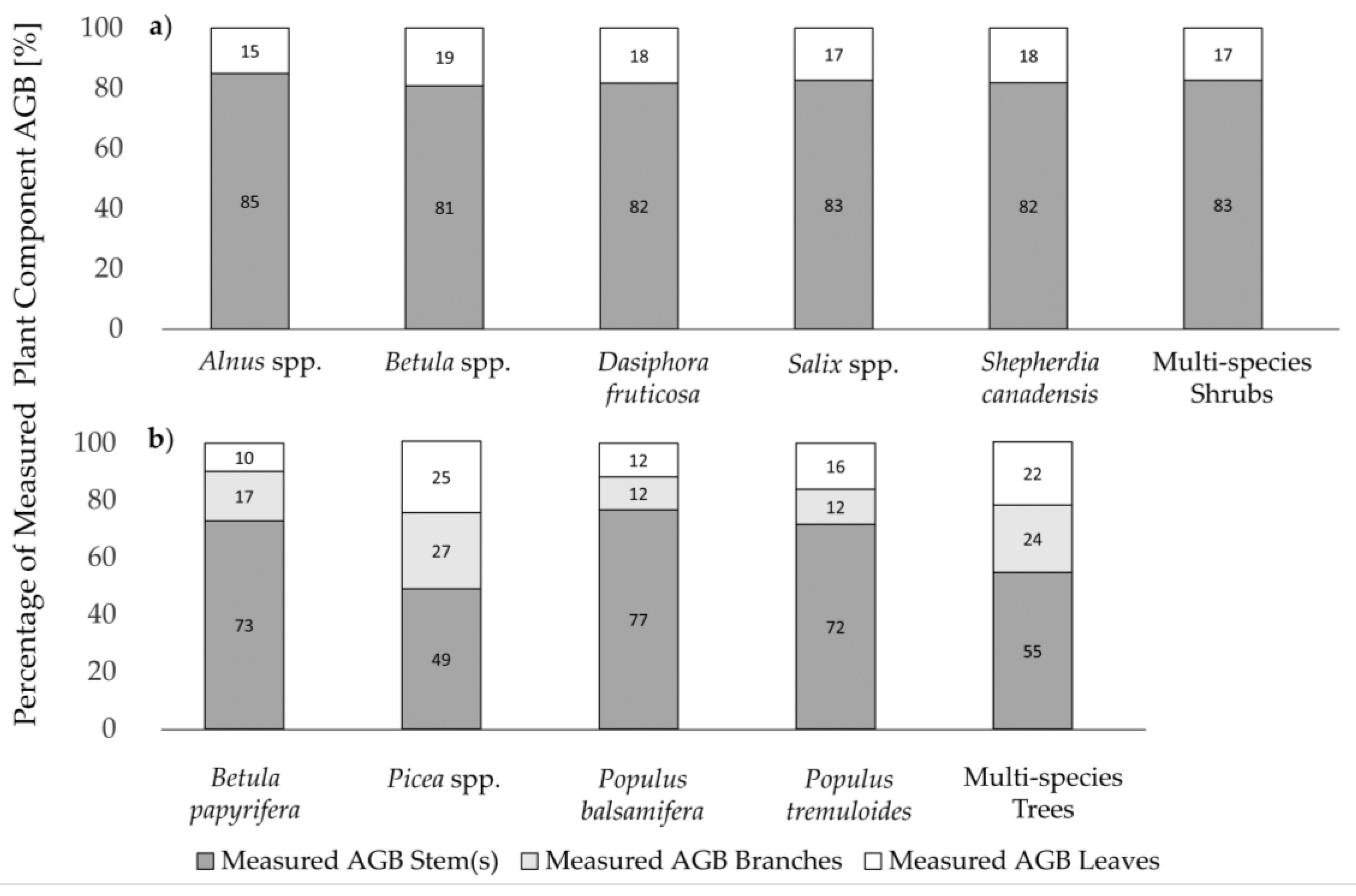
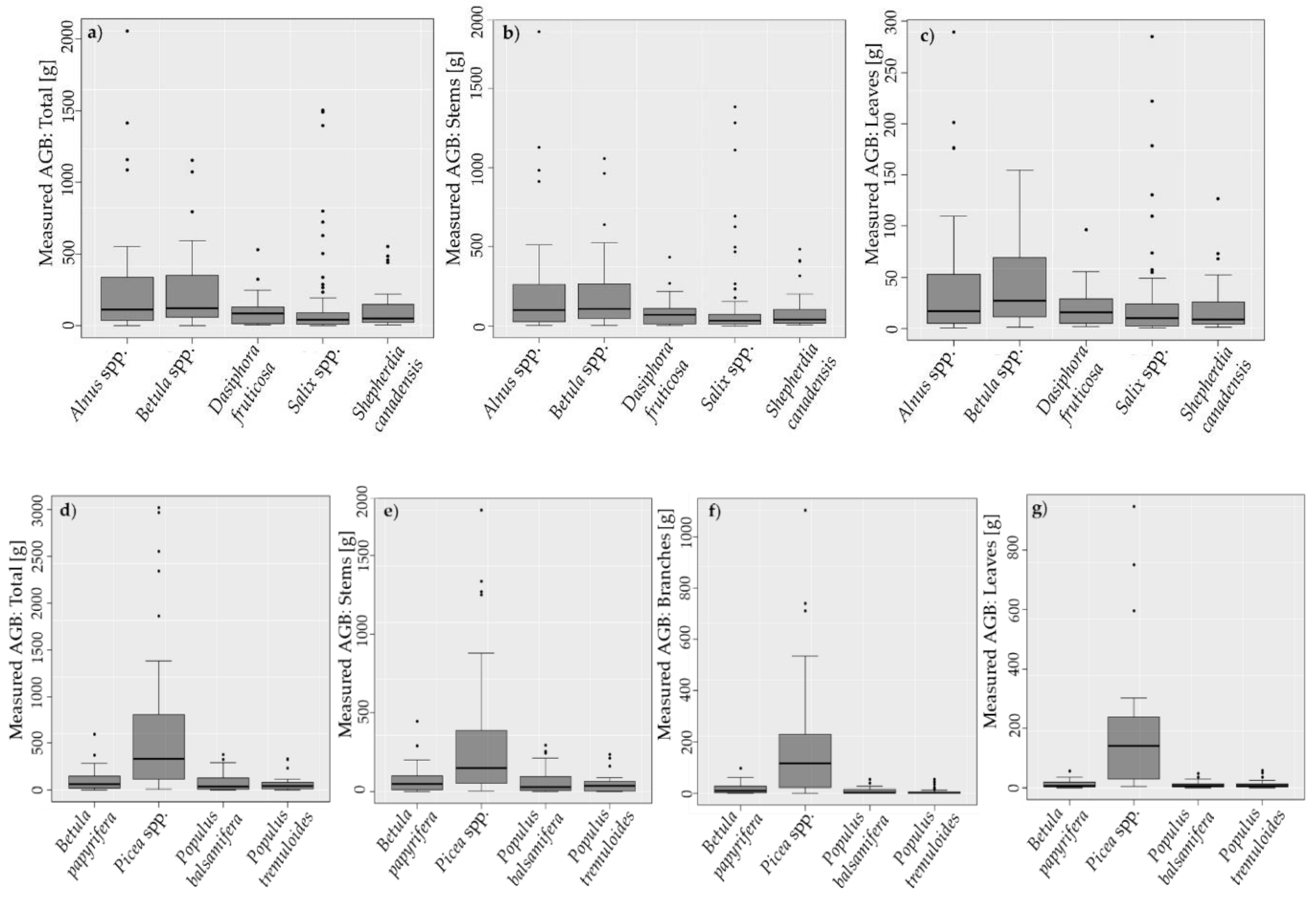
3.1. Measured Plant Component AGB
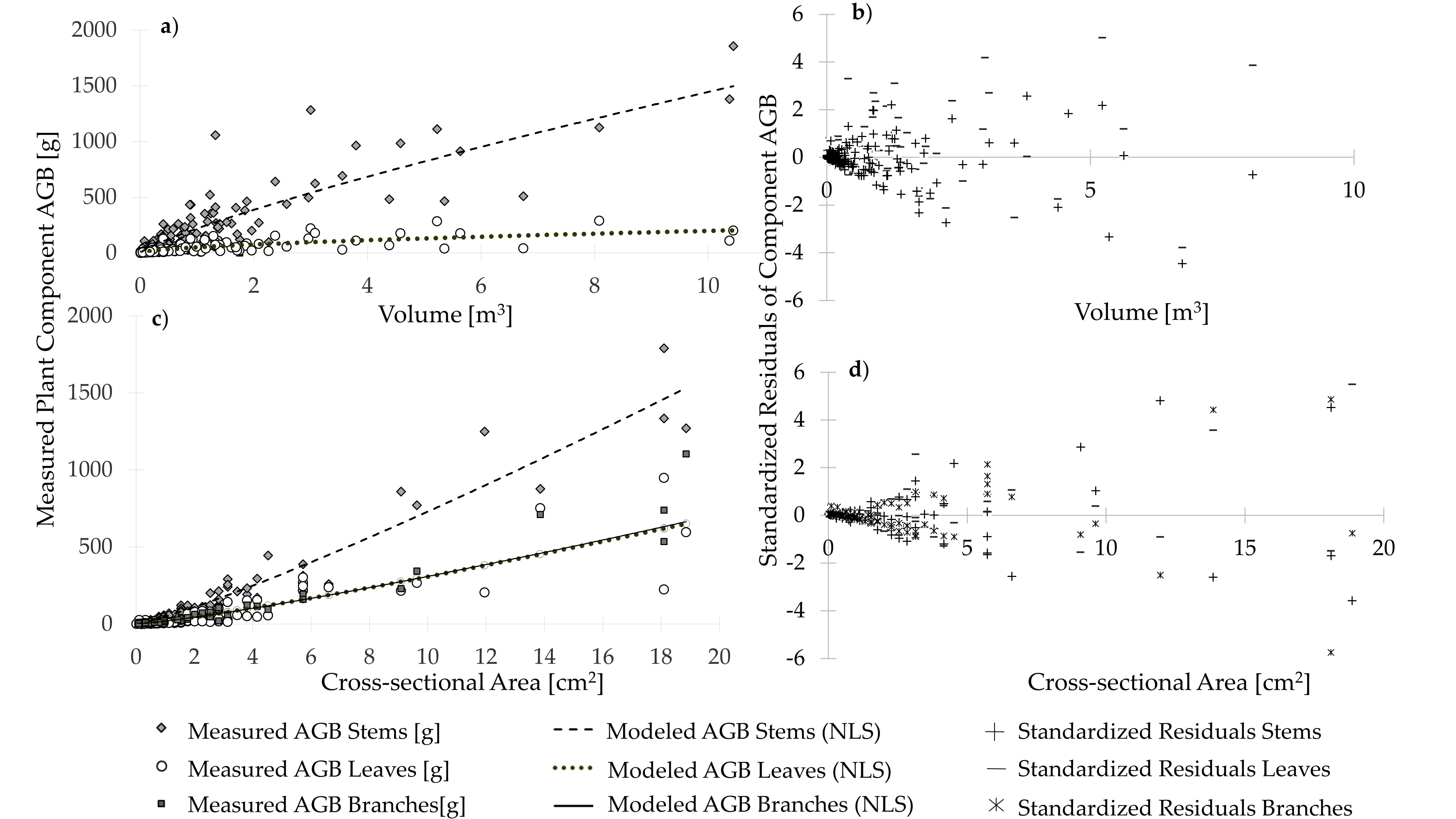
3.2. Modeled Plant Component AGB and Allometric Equations
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vitt, D.H.; Halsey, L.A.; Bauer, I.E.; Campbell, C. Spatial and temporal trends in carbon storage of peatlands of continental western Canada through the Holocene. Can. J. Earth Sci. 2000, 37, 683–693. [Google Scholar] [CrossRef]
- Kurz, W.A.; Shaw, C.H.; Boisvenue, C.; Stinson, G.; Metsaranta, J.; Leckie, D.; Dyk, A.; Smyth, C.; Neilson, E.T. Carbon in Canada’s boreal forest—A synthesis. Environ. Rev. 2013, 21, 260–292. [Google Scholar] [CrossRef]
- Bernier, P.; Kurz, W.A.; Lemprière, T.C.; Ste-Marie, C. A Blueprint for Forest Carbon Science in Canada: 2012–2020; Natural Resources Canada, Canadian Forest Service: Ottawa, ON, Canada, 2012; 52p. [Google Scholar]
- Romero-Lankao, P.; Smith, J.B.; Davidson, D.J.; Diffenbaugh, N.S.; Kinney, P.L.; Kirshen, P.; Kovacs, P.; Villers Ruiz, L. North America. In Climate Change 2014: Impacts, Adaption, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Barros, V.R., Field, C.B., Dokken, D.J., Mastrandrea, M.D., Mach, K.J., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 1439–1498. [Google Scholar]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate Change and Forest Disturbances. Bioscience 2001, 51, 723. [Google Scholar] [CrossRef] [Green Version]
- Quinton, W.L.; Hayashi, M.; Chasmer, L. Peatland Hydrology of Discontinuous Permafrost in the Northwest Territories: Overview and Synthesis. Can. Water Resour. J. 2009, 34, 311–328. [Google Scholar] [CrossRef]
- Baltzer, J.; Veness, T.; Chasmer, L.; Sniderhan, A.E.; Quinton, W.L. Forests on Thawing Permafrost: Fragmentation, Edge Effects, and Net Forest Loss. Glob. Chang. Biol. 2014, 20, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Chasmer, L.; Hopkinson, C. Threshold Loss of Discontinuous Permafrost and Landscape Evolution. Glob. Chang. Biol. 2017, 23, 2672–2686. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Forbes, B.C.; Wilmking, M.; Hallinger, M.; Lantz, T.C.; Blok, D.; Tape, K.D.; Macias-Fauria, M.; Sass-Klaassen, U.; Lévesque, E.; et al. Shrub expansion in tundra ecosystems: Dynamics, impacts and research priorities. Environ. Res. Lett. 2011, 6, 045509. [Google Scholar] [CrossRef] [Green Version]
- Myers-Smith, I.H.; Kerby, J.T.; Phoenix, G.K.; Bjerke, J.W.; Epstein, H.E.; Assmann, J.J.; John, C.; Andreu-Hayles, L.; Angers-Blondin, S.; Beck, P.S.A.; et al. Complexity revealed in the greening of the Arctic. Nat. Clim. Chang. 2020, 10, 106–117. [Google Scholar] [CrossRef] [Green Version]
- Helbig, M.; Chasmer, L.E.; Kljun, N.C.; Quinton, W.L.; Treat, C.C.; Sonnentag, O. The positive net radiative greenhouse gas forcing of increasing methane emissions from a thawing boreal forest-wetland landscape. Glob. Chang. Biol. 2017, 23, 2413–2427. [Google Scholar] [CrossRef] [Green Version]
- Goulden, M.L.; Mcmillan, A.M.S.; Winston, G.C.; Rocha, A.V.; Manies, K.L.; Harden, J.W.; Bond-Lamberty, B.P. Patterns of NPP, GPP, respiration, and NEP during boreal forest succession. Glob. Chang. Biol. 2011, 17, 855–871. [Google Scholar] [CrossRef] [Green Version]
- Goetz, S.J.; MacK, M.C.; Gurney, K.R.; Randerson, J.T.; Houghton, R.A. Ecosystem responses to recent climate change and fire disturbance at northern high latitudes: Observations and model results contrasting northern Eurasia and North America. Environ. Res. Lett. 2007, 2, 045031. [Google Scholar] [CrossRef]
- Amiro, B.; Orchansky, A.; Barr, A.; Black, T.; Chambers, S.; Iii, F.C.; Goulden, M.; Litvak, M.; Liu, H.; McCaughey, J.; et al. The effect of post-fire stand age on the boreal forest energy balance. Agric. For. Meteorol. 2006, 140, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Beck, P.S.A.; Goetz, S.J. Satellite observations of high northern latitude vegetation productivity changes between 1982 and 2008: Ecological variability and regional differences. Environ. Res. Lett. 2011, 6, 045501. [Google Scholar] [CrossRef]
- Flade, L.; Hopkinson, C.; Chasmer, L. Allometric equations for shrub and short-stature tree aboveground biomass within boreal ecosystems of northwestern Canada. Forests 2020, 11, 1207. [Google Scholar] [CrossRef]
- Ecosystem Classification Group. Ecological Regions of the Northwest. Territories—Taiga Plains. Department of Environment and Natural Resources; Government of the Northwest Territories: Yellowknife, NT, Canada, 2007.
- Ecosystem Classification Group. Ecological Regions of the Northwest. Territories—Taiga Shield. Department of Environment and Natural Resources; Government of the Northwest Territories: Yellowknife, NT, Canada, 2008.
- Baskerville, G.L. Use of logarithmic regression in the examination of plant biomass. Can. J. For. 1972, 2, 49–53. [Google Scholar]
- Mascaro, J.; Labs, P.; Litton, C.M.; Schnitzer, S. Minimizing Bias in Biomass Allometry: Model Selection and Log—Transformation of Data. Biotropica 2011, 43, 649–653. [Google Scholar] [CrossRef] [Green Version]
- Konôpka, B.; Pajtík, J.; Moravčík, M.; Lukac, M. Biomass partitioning and growth efficiency in four naturally regenerated forest tree species. Basic Appl. Ecol. 2010, 11, 234–243. [Google Scholar] [CrossRef] [Green Version]
- Petersson, H.; Holm, S.; Ståhl, G.; Alger, D.; Fridman, J.; Lehtonen, A.; Lundström, A.; Mäkipää, R. Individual tree biomass equations or biomass expansion factors for assessment of carbon stock changes in living biomass—A comparative study. For. Ecol. Manag. 2012, 270, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Johansson, T. Biomass production of Norway spruce (Picea abies (L.) Karst) growing on abandoned farmland. Silva. Fenn. 1999, 33, 261–280. [Google Scholar] [CrossRef] [Green Version]
- He, A.; McDermid, G.J.; Rahman, M.M.; Strack, M.; Saraswati, S.; Xu, B. Developing allometric equations for estimating shrub biomass in a boreal fen. Forests 2018, 9, 569. [Google Scholar] [CrossRef] [Green Version]
- Lantz, T.C.; Marsh, P.; Kokelj, S. Recent Shrub Proliferation in the Mackenzie Delta Uplands and Microclimatic Implications. Ecosystems 2013, 16, 47–59. [Google Scholar] [CrossRef]
- Moffat, N.D.; Lantz, T.C.; Fraser, R.H.; Olthof, I. Recent vegetation change (1980–2013) in the tundra ecosystems of the Tuktoyaktuk Coastlands, NWT, Canada. Arct. Antarct. Alp. Res. 2016, 48, 581–597. [Google Scholar] [CrossRef] [Green Version]
- Gower, S.T.; Vogel, J.G.; Norman, M.; Kucharik, C.J.; Steele, S.J. Carbon distribution and aboveground net primary production in aspen, jack pine, and black spruce stands in Saskatchewan and Manitoba, Canada. J. Geophys. Res. 1997, 29. [Google Scholar] [CrossRef]
- Bonan, G. Ecosystems. In Ecological Climatology; Cambridge University Press: Cambridge, UK, 2016; pp. 328–357. [Google Scholar] [CrossRef]
- Baldocchi, D.; Vogel, C. Energy and CO2 flux densities above and below a temperate broad-leaved forest and a boreal pine forest. Tree Physiol. 1996, 16, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Thomas, H.J.D.; Bjorkman, A.D.; Myers-Smith, I.H.; Elmendorf, S.C.; Kattge, J.; Diaz, S.; Vellend, M.; Blok, D.; Cornelissen, J.H.C.; Forbes, B.C.; et al. Global plant trait relationships extend to the climatic extremes of the tundra biome. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef]
- Lambert, M.-C.; Ung, C.-H.; Raulier, F. Canadian national tree aboveground biomass equations. Can. J. For. Res. 2005, 35, 1996–2018. [Google Scholar] [CrossRef]
- Ung, C.H.; Bernier, P.; Guo, X.J. Canadian national biomass equations: New parameter estimates that include British Columbia data. Can. J. For. Res. 2008, 38, 1123–1132. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Genus/Species | No. of Samples | Maximum Height [m] | Total AGB [g] | AGB Stems [g] | AGB Branches [g] | AGB Leaves/Needles [g] |
|---|---|---|---|---|---|---|
| Alnus spp. | 33 | [0.2; 3.2] 1.3 ± 0.7 | [1.3; 2057.1] 311.4 ± 470.9 | [0.6; 1856] 264.8 ± 401.8 | - | [0.4; 289.7] 46.7 ± 68.0 |
| Betula spp. | 46 | [0.2; 2.1] 1.1 ± 0.4 | [4.0; 1154.1] 232.7 ± 261.1 | [2.2; 1057.2] 188.2 ± 228.4 | - | [1.3; 154.7] 44.5 ± 42.3 |
| Dasiphora fruticosa | 20 | [0.2; 0.9] 0.6 ± 0.4 | [5.1; 530.8] 117.6 ± 127.9 | [3.6; 434.6] 96.2 ± 105.8 | - | [1.5; 96.2] 21.4 ± 22.7 |
| Salix spp. | 79 | [0.3; 2.8] 0.9 ± 0.5 | [0.8; 1503.7] 143.3 ± 302.5 | [0.4; 1381.4] 118.4 ± 261.1 | - | [0.4; 284.8] 24.9 ± 47.0 |
| Shepherdia canadensis | 28 | [0.3; 1.7] 0.8 ± 0.4 | [7.1; 552.0] 121.5 ± 158.9 | [5.7; 484.0] 99.5 ± 134.9 | - | [1.1; 127.0] 22.0 ± 28.6 |
| Betula papyrifera | 15 | [0.7, 3.4] 2.0 ± 0.8 | [4.2; 596.2] 127.7 ± 162.2 | [2.6; 444.9] 93.0 ± 122.4 | [0.8; 95.7] 22.0 ± 26.1 | [0.8; 55.6] 12.7 ± 14.8 |
| Picea glauca | 14 | [0.4; 3.8] 1.8 ± 1.2 | [10.5; 3021.8] 865.0 ± 992.4 | [3.4; 1789.6] 426.1 ± 545.6 | [1.2; 739.4] 212.5 ± 247.8 | [5.9; 947.8] 226.4 ± 270.5 |
| Picea mariana | 15 | [0.4; 3.6] 1.6 ± 0.9 | [12.5; 2968.9] 668.5 ± 801.2 | [4.3; 1269.6] 326.0 ± 412.2 | [3.1; 1103.6] 195.2 ± 270.4 | [5.1; 595.7] 157.1 ± 152.2 |
| Populus balsamifera | 31 | [0.2; 4.2] 1.7 ± 1.1 | [1.0; 380.9] 85.7 ± 103.3 | [0.7; 294.9] 65.7 ± 81.3 | [0.3; 53.1] 11.3 ± 13.7 | [0.04; 47.0] 10.1 ± 11.1 |
| Populus tremuloides | 30 | [0.4; 3.9] 1.8 ± 0.9 | [1.1; 329.2] 67.8 ± 84.3 | [0.5; 233.7] 50.2 ± 58.7 | [0.04; 52.4] 8.6 ± 13.2 | [0.2; 58.1] 10.9 ± 14.0 |
| Model | LN (β) | β | SE (β) | α | SE (α) | CF | RMSE [g] | R2 | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Alnus spp. | Stems | LLRC | 5.104 | 164.6793 | 0.9474 | 1.2166 | 163.59 | 0.882 | ||
| NLS | 146.3720 | 30.0176 | 1.0210 | 0.1021 | 137.23 | 0.885 | ||||
| Leaves | LLRC | 3.418 | 30.5083 | 0.7862 | 1.1792 | 35.04 | 0.735 | |||
| NLS | 37.0392 | 8.5131 | 0.7805 | 0.1213 | 35.01 | 0.735 | ||||
| Betula spp. | Stems | LLRC | 5.415 | 224.7525 | 0.8135 | 1.1766 | 137.05 | 0.651 | ||
| NLS | 275.7010 | 28.2444 | 0.8980 | 0.1222 | 134.39 | 0.654 | ||||
| Leaves | LLRC | 3.977 | 53.3567 | 0.6370 | 1.2068 | 27.51 | 0.578 | |||
| NLS | 64.8446 | 5.1663 | 0.6047 | 0.1019 | 27.48 | 0.579 | ||||
| Dasiphora fruticosa | Stems | LLRC | 5.350 | 210.6083 | 0.7564 | 1.1608 | 60.72 | 0.672 | ||
| NLS | 255.6900 | 31.3893 | 0.8490 | 0.2042 | 60.33 | 0.675 | ||||
| Leaves | LLRC | 3.714 | 41.0175 | 0.6228 | 1.1800 | 13.20 | 0.676 | |||
| ILS | 55.9038 | 6.56778 | 0.8269 | 0.1911 | 12.62 | 0.691 | ||||
| Salix spp. | Stems | LLRC | 5.161 | 174.3387 | 0.8857 | 1.1364 | 112.78 | 0.814 | ||
| NLS | 210.9940 | 22.3109 | 0.8320 | 0.0588 | 111.90 | 0.817 | ||||
| Leaves | LLRC | 3.664 | 39.0171 | 0.7380 | 1.1767 | 33.75 | 0.519 | |||
| ILS | 48.6847 | 5.6819 | 0.5734 | 0.0728 | 31.76 | 0.546 | ||||
| Shepherdia canadensis | Stems | LLRC | 5.073 | 159.6526 | 0.7601 | 1.1018 | 62.16 | 0.789 | ||
| NLS | 192.057 | 18.0495 | 0.6690 | 0.0849 | 60.68 | 0.801 | ||||
| Leaves | LLRC | 3.504 | 33.2482 | 0.6807 | 1.2342 | 23.74 | 0.380 | |||
| NLS | 38.9539 | 5.84277 | 0.4535 | 0.1263 | 21.77 | 0.427 | ||||
| Multi- species Shrubs | Stems | LLRC | 5.240 | 188.6701 | 0.8642 | 1.1842 | 123.31 | 0.795 | ||
| NLS | 220.1460 | 12.7277 | 0.8170 | 0.0337 | 120.10 | 0.796 | ||||
| Leaves | LLRC | 3.692 | 40.1250 | 0.7151 | 1.2335 | 31.52 | 0.586 | |||
| NLS | 50.4742 | 2.9920 | 0.5945 | 0.0393 | 30.12 | 0.600 | ||||
| Model | LN (β) | β | SE (β) | α | SE (α) | CF | RMSE [g] | R2 | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Betula papyrifera | Stems | LLRC | 3.970 | 52.9845 | 1.2370 | 1.0660 | 40.88 | 0.898 | ||
| NLS | 41.4994 | 11.9728 | 1.5494 | 0.2193 | 36.63 | 0.913 | ||||
| Branches | LLRC | 2.634 | 13.9294 | 1.1440 | 1.0542 | 8.33 | 0.900 | |||
| NLS | 12.7152 | 2.95996 | 1.2921 | 0.1826 | 8.08 | 0.905 | ||||
| Leaves | LLRC | 2.124 | 8.3645 | 1.0140 | 8.6012 | 70.00 | 0.882 | |||
| NLS | 7.8627 | 2.1302 | 1.2143 | 0.2150 | 5.70 | 0.853 | ||||
| Picea spp. | Stems | LLRC | 3.894 | 49.1069 | 1.0670 | 1.0782 | 170.97 | 0.928 | ||
| NLS | 42.8437 | 11.9427 | 1.2201 | 0.1030 | 129.29 | 0.929 | ||||
| Branches | LLRC | 3.430 | 30.8766 | 1.0230 | 1.1129 | 94.07 | 0.892 | |||
| NLS | 19.1842 | 6.6497 | 1.2866 | 0.1272 | 80.78 | 0.904 | ||||
| Leaves | LLRC | 3.821 | 45.6498 | 0.8059 | 1.1088 | 123.21 | 0.702 | |||
| NLS | 42.2998 | 17.9815 | 0.9148 | 0.1635 | 119.89 | 0.704 | ||||
| Populus balsamifera | Stems | LLRC | 3.805 | 44.9253 | 1.1320 | 1.1648 | 25.46 | 0.919 | ||
| NLS | 47.3512 | 5.5776 | 1.3235 | 0.1053 | 22.43 | 0.924 | ||||
| Branches | LLRC | 1.773 | 5.8885 | 1.1140 | 1.4758 | 6.88 | 0.736 | |||
| NLS | 7.6872 | 1.8698 | 1.2340 | 0.2211 | 6.83 | 0.737 | ||||
| Leaves | LLRC | 2.028 | 7.5989 | 1.0370 | 1.4577 | 5.81 | 0.832 | |||
| ILS | 8.31954 | 1.1695 | 1.1007 | 0.1313 | 4.53 | 0.834 | ||||
| Populus tremuloides | Stems | LLRC | 3.770 | 43.3801 | 1.0670 | 1.1627 | 13.64 | 0.950 | ||
| NLS | 41.7058 | 3.0681 | 1.2658 | 0.0680 | 11.72 | 0.960 | ||||
| Branches | LLRC | 1.682 | 5.3763 | 1.0910 | 1.5380 | 6.18 | 0.816 | |||
| NLS | 4.5910 | 1.0036 | 1.8127 | 0.1858 | 4.42 | 0.888 | ||||
| Leaves | LLRC | 2.255 | 9.5353 | 0.8919 | 1.1365 | 6.50 | 0.854 | |||
| NLS | 8.07253 | 1.0364 | 1.4362 | 0.1149 | 4.11 | 0.917 | ||||
| Hardwood Trees | Stems | LLRC | 3.830 | 46.0625 | 1.1220 | 1.1482 | 30.87 | 0.887 | ||
| NLS | 41.4212 | 3.9694 | 1.4431 | 0.0807 | 26.48 | 0.904 | ||||
| Branches | LLRC | 1.927 | 6.8689 | 1.1270 | 1.4893 | 8.7 | 0.760 | |||
| NLS | 6.4825 | 1.1607 | 1.5966 | 0.1475 | 8.00 | 0.789 | ||||
| Leaves | LLRC | 2.138 | 8.4825 | 0.9655 | 1.2434 | 5.57 | 0.826 | |||
| NLS | 8.28683 | 0.8242 | 1.2225 | 0.0872 | 4.44 | 0.841 | ||||
| Multi- species Trees | Stems | LLRC | 3.833 | 46.2009 | 1.1090 | 1.1296 | 80.19 | 0.939 | ||
| NLS | 49.1790 | 5.0772 | 1.1712 | 0.0393 | 73.17 | 0.940 | ||||
| Branches | LLRC | 2.336 | 10.3398 | 1.2690 | 1.6568 | 51.61 | 0.919 | |||
| NLS | 12.7920 | 2.2283 | 1.4315 | 0.0639 | 45.08 | 0.923 | ||||
| Leaves | LLRC | 2.584 | 13.2500 | 1.1070 | 1.5055 | 73.11 | 0.768 | |||
| NLS | 20.2891 | 4.6584 | 1.1807 | 0.0872 | 67.96 | 0.767 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flade, L.; Hopkinson, C.; Chasmer, L. Aboveground Biomass Allocation of Boreal Shrubs and Short-Stature Trees in Northwestern Canada. Forests 2021, 12, 234. https://doi.org/10.3390/f12020234
Flade L, Hopkinson C, Chasmer L. Aboveground Biomass Allocation of Boreal Shrubs and Short-Stature Trees in Northwestern Canada. Forests. 2021; 12(2):234. https://doi.org/10.3390/f12020234
Chicago/Turabian StyleFlade, Linda, Christopher Hopkinson, and Laura Chasmer. 2021. "Aboveground Biomass Allocation of Boreal Shrubs and Short-Stature Trees in Northwestern Canada" Forests 12, no. 2: 234. https://doi.org/10.3390/f12020234
APA StyleFlade, L., Hopkinson, C., & Chasmer, L. (2021). Aboveground Biomass Allocation of Boreal Shrubs and Short-Stature Trees in Northwestern Canada. Forests, 12(2), 234. https://doi.org/10.3390/f12020234