
Variations in Organic Carbon Content and Dehydrogenases Activity in Post-Agriculture Forest Soils: A Case Study in South-Western Pomerania


Abstract
1. Introduction
2. Materials and Methods
2.1. Description of the Research Area
2.2. Soil Sampling and Preparation
2.3. Laboratory Methods and Analyses
2.4. Statistical Analysis
3. Results
3.1. Organic Carbon Content in Topsoil Horizons
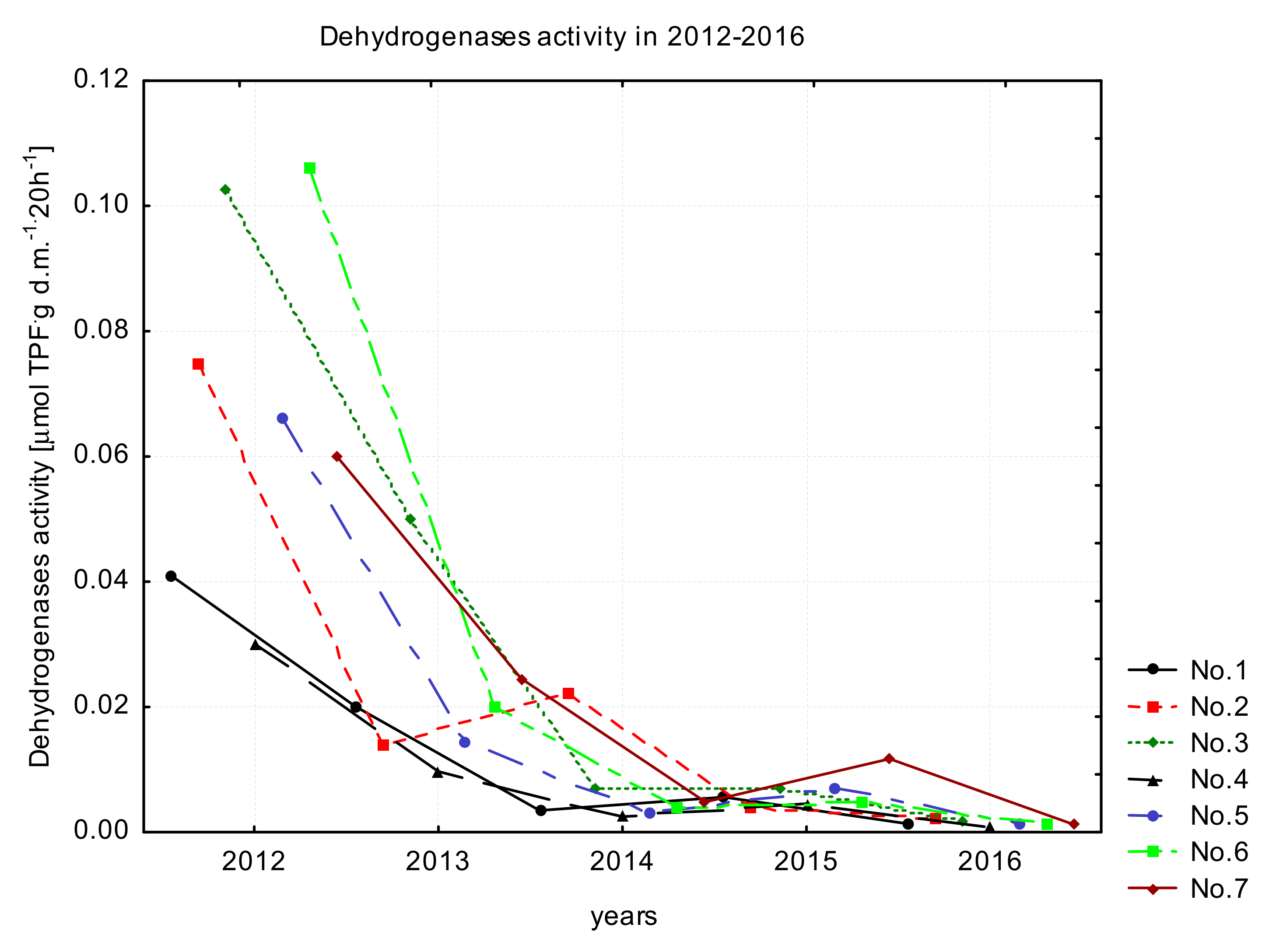
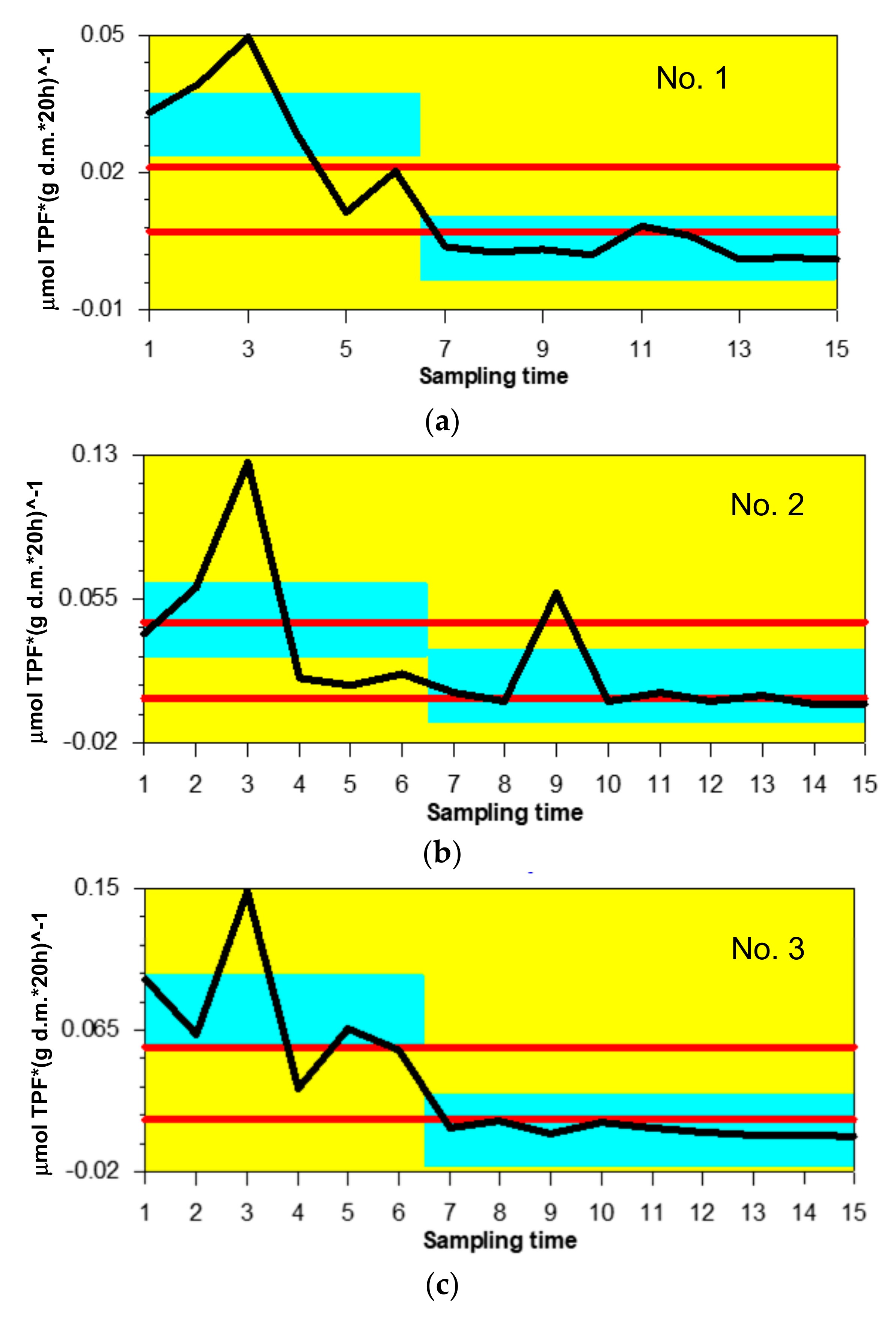
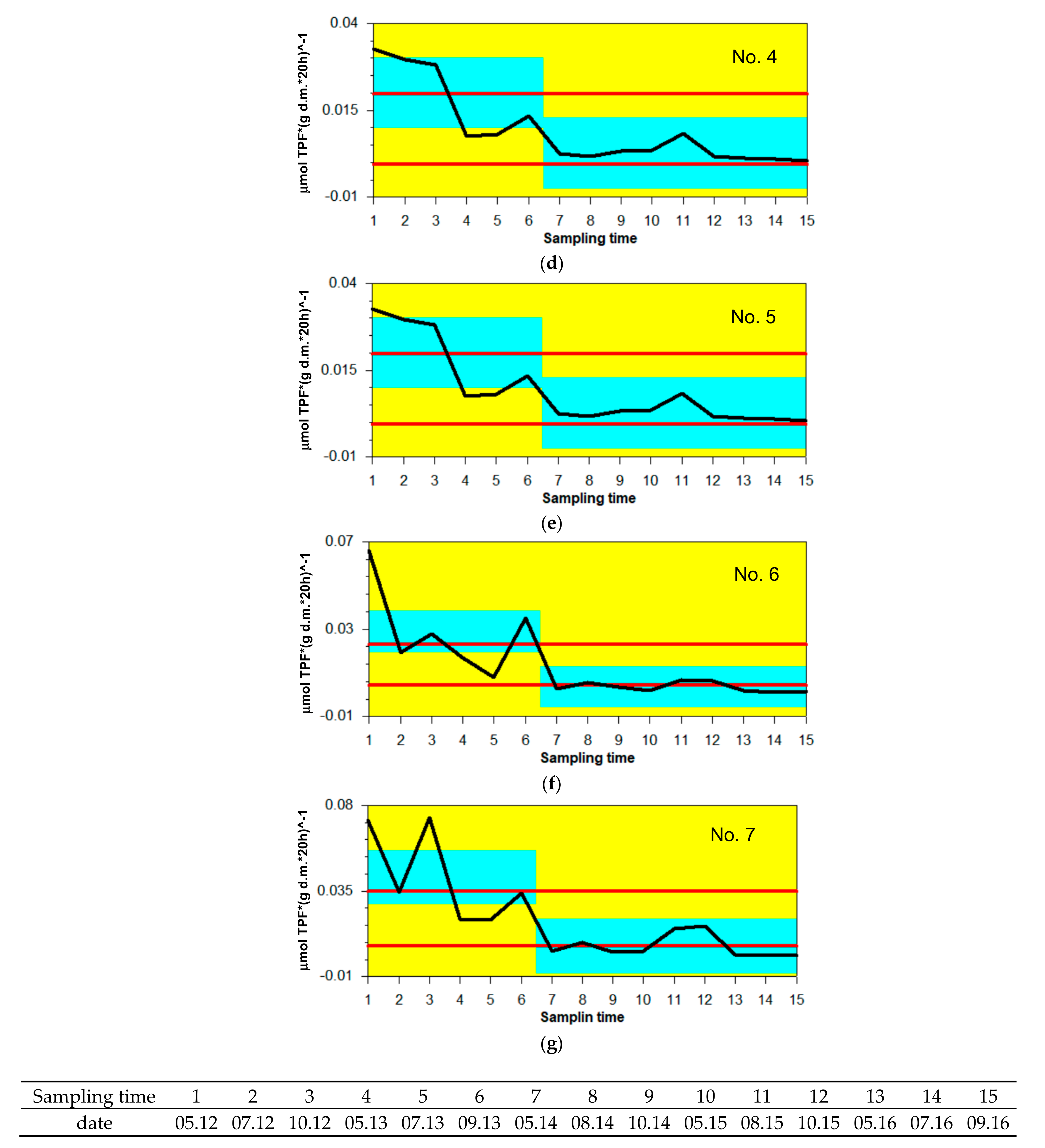
3.2. Variability in OC Content and DHA
4. Discussion
4.1. Factors Controling OC Content
4.2. Variations in OC Content and DHA
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alemu, B. The Role of Forest and Soil Carbon Sequestrations on Climate Change Mitigation. J. Agric. Environ. Manag. 2014, 3, 492–505. [Google Scholar]
- Lal, R. Soil carbon sequestration Impacts on global climate change and food security. Science 2004, 304, 1623. [Google Scholar] [CrossRef] [PubMed]
- Lal, R. Soil carbon sequestration to mitigate climate change. Geoderma 2004, 123, 1–22. [Google Scholar] [CrossRef]
- Lal, R. Carbon sequestration. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 815–830. [Google Scholar] [CrossRef] [PubMed]
- Paul, K.I.; Polglase, P.J.; Nyakuengama, J.G.; Khanna, P.K. Change in soil carbon following afforestation. For. Ecol. Manag. 2002, 168, 241–257. [Google Scholar] [CrossRef]
- Deng, L.; Zhu, G.Y.; Tang, Z.S.; Shangguan, Z.P. Global patterns of the effects of land-use changes on soil carbon stocks. Glob. Ecol. Conserv. 2016, 5, 127–138. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef]
- Shi, S.; Zhang, W.; Zhang, P.; Yu, Y.; Fan, D. A synthesis of change in deep soil organic carbon stores with afforestation of agricultural soils. For. Ecol. Manag. 2013, 296, 53–63. [Google Scholar] [CrossRef]
- Christensen, B.T. Decomposability of organic matter in particle size fractions from field soils with straw incorporation. Soil Biol. Biochem. 1987, 4, 429–435. [Google Scholar] [CrossRef]
- Ontl, T.A.; Schulte, L.A. Soil Carbon Storage. Nat. Educ. Knowl. 2012, 3, 35. [Google Scholar]
- Sun, X.; Tang, Z.; Ryan, M.G.; You, Y.; Sun, O.J. Changes in soil organic carbon contents and fractionations of forests along a climatic gradient in China. For. Ecosyst. 2019, 6, 1. [Google Scholar] [CrossRef]
- Lützow, M.; Kögel-Knabner, I.; Ekschmitt, K.; Matzner, E.; Guggenberger, G.; Marschner, B.; Flessa, H. Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions—A review. Eur. J. Soil Sci. 2006, 57, 426–445. [Google Scholar] [CrossRef]
- Gower, S.T. Patterns and mechanisms of the forest carbon cycle. Annu. Rev. Environ. Resour. 2003, 28, 169–204. [Google Scholar] [CrossRef]
- IPCC. Summary for policy makers. In Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 1–18. [Google Scholar]
- McKinley, D.; Ryan, M.; Birdsey, R.; Giardina, C.; Harmon, M.; Heath, L.; Houghton, R.; Jackson, R.; Morrison, J.; Murray, B. A synthesis of current knowledge on forests and carbon storage in the United States. Ecol. Appl. 2011, 21, 1902–1924. [Google Scholar] [CrossRef]
- Smal, H.; Ligeza, S.; Olszewska, M. The effect of afforestation of sandy post-arable soils on the quality of organic matter and chemical composition of soils solution. Soil Sci. Annu. 2004, 55, 139–148. [Google Scholar]
- Dovydenko, N. Contents of carbon in soil of selected pine and spruce stands occurring on post-agricultural lands. For. Res. Inst. 2004, 2, 49–66. [Google Scholar]
- Degórski, M. Influence of forest use on soil organic carbon reserves. Monit. Nat. Environ. 2005, 6, 75–83. [Google Scholar]
- Smal, H.; Olszewska, M. The effect of afforestation with Scots pine (Pinus silvestris L.) of sandy post-arable soils on their selected properties. II. Reaction, carbon, nitrogen and phosphorous. Plant Soil 2008, 305, 171–187. [Google Scholar] [CrossRef]
- Li, T.; Ren, B.; Wang, D.; Liu, G. Spatial variation in the storages and age-related dynamics of forest carbon sequestration in different climate zones—evidence from black locust plantations on the Loess Plateau of China. PLoS ONE 2015, 10, e0121862. [Google Scholar] [CrossRef]
- Wiesmeier, M.; Prietzel, J.; Barthold, F.; Spörlein, P.; Geuß, U.; Hangen, E.; Reischl, A.; Schilling, B.; von Lützow, M.; Kögel-Knabner, I. Storage and drivers of organic carbon in forest soils of southeast Germany (Bavaria)—Implications for carbon sequestration. For. Ecol. Manag. 2013, 295, 162–172. [Google Scholar] [CrossRef]
- Powers, R.F.; Busse, M.D.; McFarlane, K.J.; Zhang, J.; Young, D.H. Long-term effects of silviculture on soil carbon storage: Does vegetation control make a difference? Forestry 2013, 86, 47–58. [Google Scholar] [CrossRef]
- Kondracki, J. Regional Geography of Poland; PWN: Warszawa, Poland, 2002. [Google Scholar]
- Krogulec, E.; Wierchowiec, J.; Bujakowska, K.; Bojakowska, I.; Kwecko, P.; Pasieczna, A.; Tomassi-Morawiec, H. Explanations to the Geo-Environmental Map of Poland 1: 50,000; Warsaw PGI-NRI: Warszawa, Poland, 2009. [Google Scholar]
- Ziemblińska, K.; Urbaniak, M.; Chojnicki, B.H.; Black, T.A.; Niu, S.; Olejnik, J. Net ecosystem productivity and its environmental controls in a mature Scots pine stand in north-western Poland. Agric. For. Meteorol. 2016, 228–229, 60–72. [Google Scholar] [CrossRef]
- World reference base for soil resources 2014. In International Soil Classification System for Naming Soils and Creating Legends for Soil Maps Update; FAO: Rome, Italy, 2015.
- Komisarek, J.; Wiatrowska, K. Changes in organic carbon content in post-agricultural forest soils in Tuczno reaserch area. In The Role of the Forest in Absorbing Carbon Dioxide from the Atmosphere; Olejnik, J., Małek, S., Eds.; University Publisher, Poznan University of Life Sciences: Poznan, Poland, 2020; pp. 271–291. [Google Scholar]
- Schoeneberger, P.J.; Wysocki, D.A.; Benham, E.C. Soil Survey Staff. In Field Book for Describing and Sampling Soils, Version 3.0; Natural Resources Conservation Service, National Soil Survey Center: Lincoln, NE, USA, 2012. [Google Scholar]
- Schumacher, B.A. Methods for the Determination of Total Organic Carbon (TOC) in Soils and Sediments; United States Environmental Protection Agency, Environmental Sciences Division National, Exposure Research Laboratory: Washington, DC, USA, 2002.
- Santi, C.; Certini, G.; D’Acqui, L.P. Direct Determination of Organic Carbon by Dry Combustion in Soils with Carbonates. Commun. Soil Sci. Plan. 2006, 37, 155–162. [Google Scholar] [CrossRef]
- Rebecca, B. (Ed.) Soil Survey Laboratory Methods Manual. In Soil Survey Investigations Report No. 42 version 4.0 November 2004; NRCS Natural Resources Conservations Service: Washington, DC, USA, 2004. [Google Scholar]
- Ostrowska, A.; Gawliński, S.; Szczubiałka, Z. Methods of Analysis and Assessment of the Soil and Plants; State Institute of Environmental Protection: Warszawa, Poland, 1991. [Google Scholar]
- Tabatai, M.A.; Bremner, J.M. Soil enzymes. In Methods of Soil Analysis Part 2, 2nd ed.; American Society of Agronomy Inc.: Madison, WI, USA, 1969; pp. 903–968. [Google Scholar]
- Siti, N.A.M.A.; Mohamad, F.M.M.; Azuraliza, A.B.; Abdul, R.H.; Sharifah, M.; Syed, A. Change point analysis: A statistical approach to detect potential abrupt change. J. Teknol. 2017, 79, 147–159. [Google Scholar]
- Beaulieu, C.; Chen, J.; Sarmiento, J.L. Change Point Analysis as a Tool to Detect Abrupt Climate Variations. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2012, 370, 1228–1249. [Google Scholar] [CrossRef] [PubMed]
- Zak, D.R.; Grigal, D.F.; Gleeson, S.; Tilman, D. Carbon and nitrogen cycling during old-field succession: Constraints on plant and microbial biomass. Biogeochemistry 1990, 11, 111–129. [Google Scholar] [CrossRef]
- Vesterdal, L.; Ritter, E.; Gundersen, P. Change in soil organic carbon following afforestation of former arable land. For. Ecol. Manage. 2002, 169, 137–147. [Google Scholar] [CrossRef]
- Ritter, E. Carbon, nitrogen and phosphorus in volcanic soils following afforestation with native birch (Betula pubescens) and introduced larch (Larix sibirica) in Iceland. Plant Soil 2007, 295, 239–251. [Google Scholar] [CrossRef]
- Laganiere, J.; Angers, D.A.; Pare, D. Carbon accumulation in agricultural soils after afforestation: A meta-analysis. Glob. Chang. Biol. 2010, 16, 439–453. [Google Scholar] [CrossRef]
- Kang, H.; Gao, H.; Yu, W.; Yi, Y.; Wang, Y.; Ning, M. Changes in soil microbial community structure and function after afforestation depend on species and age: Case study in a subtropical alluvial island. Sci. Total Environ. 2018, 625, 1423–1432. [Google Scholar] [CrossRef]
- Dłużewski, P.; Wiatrowska, K.; Kozłowski, M. Seasonal changes in organic carbon content in post-arable forest soils. Soil Sci. Annu. 2019, 70, 3–12. [Google Scholar] [CrossRef]
- Six, J.; Frey, S.D.; Thiet, R.K.; Batten, K.M. Bacterial and fungal contribution to carbon sequestration in agrosystems. Soil Sci. Soc. Am. J. 2006, 70, 555–569. [Google Scholar] [CrossRef]
- Busse, M.D.; Sanchez, F.G.; Ratcliff, A.W.; Butnor, J.R.; Carter, E.A.; Powers, R.F. Soil carbon sequestration and changes in fungal and bacterial biomass following incorporation of forest residue. Soil Biol. Biochem. 2019, 41, 220–227. [Google Scholar] [CrossRef]
- Gu, Y.; Wag, P.; Kong, C. Urease, Invertase, Dehydrogenase and Polyphenoloxidase Activities in Paddy Soils Influenced By Allelophatic Rice variety. Europ. J. Soil Biol. 2009, 45, 436–441. [Google Scholar] [CrossRef]
- Quilchano, C.; Marañon, T. Dehydrogenase activity in mediterranean forest soils. Biol. Fertil. Soils 2002, 35, 102–107. [Google Scholar] [CrossRef]
- Salazar, S.; Sanchez, L.; Alvarez, J.; Valverde, A.; Galindo, P.; Igual, J.; Peix, A.; Santa-Regina, I. Correlation Among Soil Enzyme Activities Under Different Forest System Management Practices. Ecol. Eng. 2011, 37, 1123–1131. [Google Scholar] [CrossRef]
- Moeskops, B.; Buchan, D.; Sleutel, S.; Herawaty, L.; Husen, E.; Saraswati, R.; Setyorini, D.; De Neve, S. Soil Microbial Communities and Activities Under Intensive Organic and Conventional Vegetable Farming in West Java, Indonesia. Appl. Soil Ecol. 2010, 45, 112–120. [Google Scholar] [CrossRef]
- Zhao, B.; Chen, J.; Zhang, J.; Qin, S. Soil Microbial Biomass and Activity Response to Repeated Drying-Rewetting Cycles Along a Soil Fertility Gradient Modified by Long-Term Fertilization Management Practices. Geoderma 2010, 160, 218–224. [Google Scholar] [CrossRef]
- Yuan, B.; Yue, D. Soil Microbial and Enzymatic Activities Across a Chronosequence of Chinese Pine Plantation Development on the Loess Plateau of China. Pedosphere 2012, 22, 1–12. [Google Scholar] [CrossRef]
- Geisseler, D.; Horwath, W.; Scow, K. Soil Moisture and Plant Residue Addition Interact in Their Effect on Extracellular Enzyme Activity. Pedobiologia 2011, 54, 71–78. [Google Scholar] [CrossRef]
- Wolińska, A.; Stępniewska, Z. Dehydrogenase activity in the soil environment. Dehydrogenases 2012, 10, 183–210. [Google Scholar] [CrossRef]
- Bolat, I. Seasonal Changes in Microbial Biomass Carbon (CMIC), Nitrogen (NMIC), Phosphorus (PMIC), and Microbial Respiration of Forest Floor and Top Soil under Beech, Fir and Mixed Fir-Beech Forest Stands. Ph.D. Thesis, Graduate School of Natural and Applied Sciences, Department of Forestry Engineering, Bartın University, Bartın, Turkey, 2011. [Google Scholar]
- Babur, E.; Dindaroglu, T. Seasonal changes of soil organic carbon and microbial biomass carbon in different forest ecosystems. Environ. Factors Affect. Hum. Health 2020, 115–123. [Google Scholar] [CrossRef]
- Gubler, A.; Wächter, D.; Schwab, P.; Müller, M.; Keller, A. Twenty-five years of observations of soil organic carbon in Swiss cropland showing stability overall but with some divergent trends. Environ. Monit. Asess. 2019, 191, 277. [Google Scholar] [CrossRef] [PubMed]
- Leinweber, P.; Schulten, H.R.; Körschens, M. Seasonal variations of soil organic matter in a long-term agricultural experiment. Plant Soil 1994, 160, 225–235. [Google Scholar] [CrossRef]
- Wuest, S. Seasonal variation in soil organic carbon. Soil Sci. Soc. Am. J. 2014, 78, 1442. [Google Scholar] [CrossRef]
- Zhang, N.; He, X.; Gao, Y.; Li, Y.; Wang, H.; Ma, D.; Zhang, R.; Yang, S. Pedogenic Carbonate and Soil Dehydrogenase Activity in Response to Soil Organic Matter in Artemisia ordosica Community. Pedosphere 2010, 20, 229–235. [Google Scholar] [CrossRef]
- Wolińska, A.; Bennicelli, R. Dehydrogenase Activity Response to Soil Reoxidation Process Described as Varied Condition of Water Potential, Air Porosity and Oxygen Availability. Pol. J. Environ. Stud. 2010, 19, 651–657. [Google Scholar]
- Wolińska, A.; Stępniewska, Z. Microorganisms Abundance and Dehydrogenase Activity as a Consequence of Soil Reoxidation Process. In Soil Tillage & Microbial Activities; Miransari, M., Ed.; Research Singpost: Kerala, India, 2011; pp. 111–143. [Google Scholar]
- Piotrowska, A.; Długosz, J. Spatio–temporal variability of microbialbiomass content and activities related to some physicochemicalproperties of Luvisols. Geoderma 2012, 173–174, 199–208. [Google Scholar] [CrossRef]
- Dłużewski, P. Determination of the Variation in the Content of Organic Carbon and Enzymatic Activity in Brunic Arenosols in Different Forest Stand Age of Pinus sp. Ph.D. Thesis, Poznan University of Life Science, Poznań, Poland, 2019. [Google Scholar]
- Zhang, M.; Dong, L.G.; Fei, S.X.; Zhang, J.W.; Jiang, X.M.; Wang, Y.; Yu, X. Responses of Soil Organic Carbon Mineralization and Microbial Communities to Leaf Litter Addition under Different Soil Layers. Forests 2021, 12, 170. [Google Scholar] [CrossRef]
- Urbaniak, M.; Ziemblińska, K.; Dukat, P.; Olejnik, J. Carbon dioxide exchange streams in pine stands around measuring sites: Tuczno, Mezyk, tlen1 and Tlen2. In The Role of the Forest in Absorbing Carbon Dioxide from the Atmosphere; Olejnik, J., Małek, S., Eds.; University Publisher, Poznan University of Life Sciences: Poznań, Poland, 2020; pp. 187–269. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Research Plot | Descriptive Statistics | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Plot Number/Age of the Stand | Altitude [m a.s.l.] | O Horizon (n = 28) * | A Horizon (n = 75) * | ||||||
| Average | Minimum | Maximum | Standard Deviation | Average | Minimum | Maximum | Standard Deviation | ||
| No. 1 52 year-old | 106 | 221.3 | 120.0 | 392.1 | 75.86 | 9.40 | 5.16 | 17.02 | 2.71 |
| No. 2 60 year-old | 106 | 386.1 | 157.0 | 538.0 | 109.9 | 12.71 | 6.85 | 31.45 | 4.82 |
| No. 3 60 year-old | 105 | 357.0 | 124.2 | 557.0 | 119.5 | 15.79 | 6.25 | 37.91 | 6.37 |
| No. 4 50 year-old | 106 | 312.2 | 120.0 | 492.8 | 109.2 | 11.16 | 6.41 | 23.93 | 3.29 |
| No. 5 60 year-old | 106 | 307.6 | 156.0 | 485.2 | 97.96 | 9.61 | 5.28 | 17.76 | 2.89 |
| No. 6 60 year-old | 103 | 312.2 | 120 | 484.4 | 99.43 | 10.73 | 5.44 | 20.51 | 3.79 |
| No. 7 60 year-old | 108 | 412.6 | 248.2 | 520.7 | 85.49 | 11.06 | 6.91 | 18.49 | 3.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiatrowska, K.; Komisarek, J.; Olejnik, J. Variations in Organic Carbon Content and Dehydrogenases Activity in Post-Agriculture Forest Soils: A Case Study in South-Western Pomerania. Forests 2021, 12, 459. https://doi.org/10.3390/f12040459
Wiatrowska K, Komisarek J, Olejnik J. Variations in Organic Carbon Content and Dehydrogenases Activity in Post-Agriculture Forest Soils: A Case Study in South-Western Pomerania. Forests. 2021; 12(4):459. https://doi.org/10.3390/f12040459
Chicago/Turabian StyleWiatrowska, Katarzyna, Jolanta Komisarek, and Janusz Olejnik. 2021. "Variations in Organic Carbon Content and Dehydrogenases Activity in Post-Agriculture Forest Soils: A Case Study in South-Western Pomerania" Forests 12, no. 4: 459. https://doi.org/10.3390/f12040459
APA StyleWiatrowska, K., Komisarek, J., & Olejnik, J. (2021). Variations in Organic Carbon Content and Dehydrogenases Activity in Post-Agriculture Forest Soils: A Case Study in South-Western Pomerania. Forests, 12(4), 459. https://doi.org/10.3390/f12040459