
Long-Term Soil Fertility and Site Productivity in Stem-Only and Whole-Tree Harvested Stands in Boreal Forest of Quebec (Canada)


Abstract
:1. Introduction
- WTH sites have smaller soil element pools than SOH sites;
- Soil responses to biomass harvesting intensity differ among soil regions;
- Forest stands grow less following WTH than following SOH.
2. Materials and Method
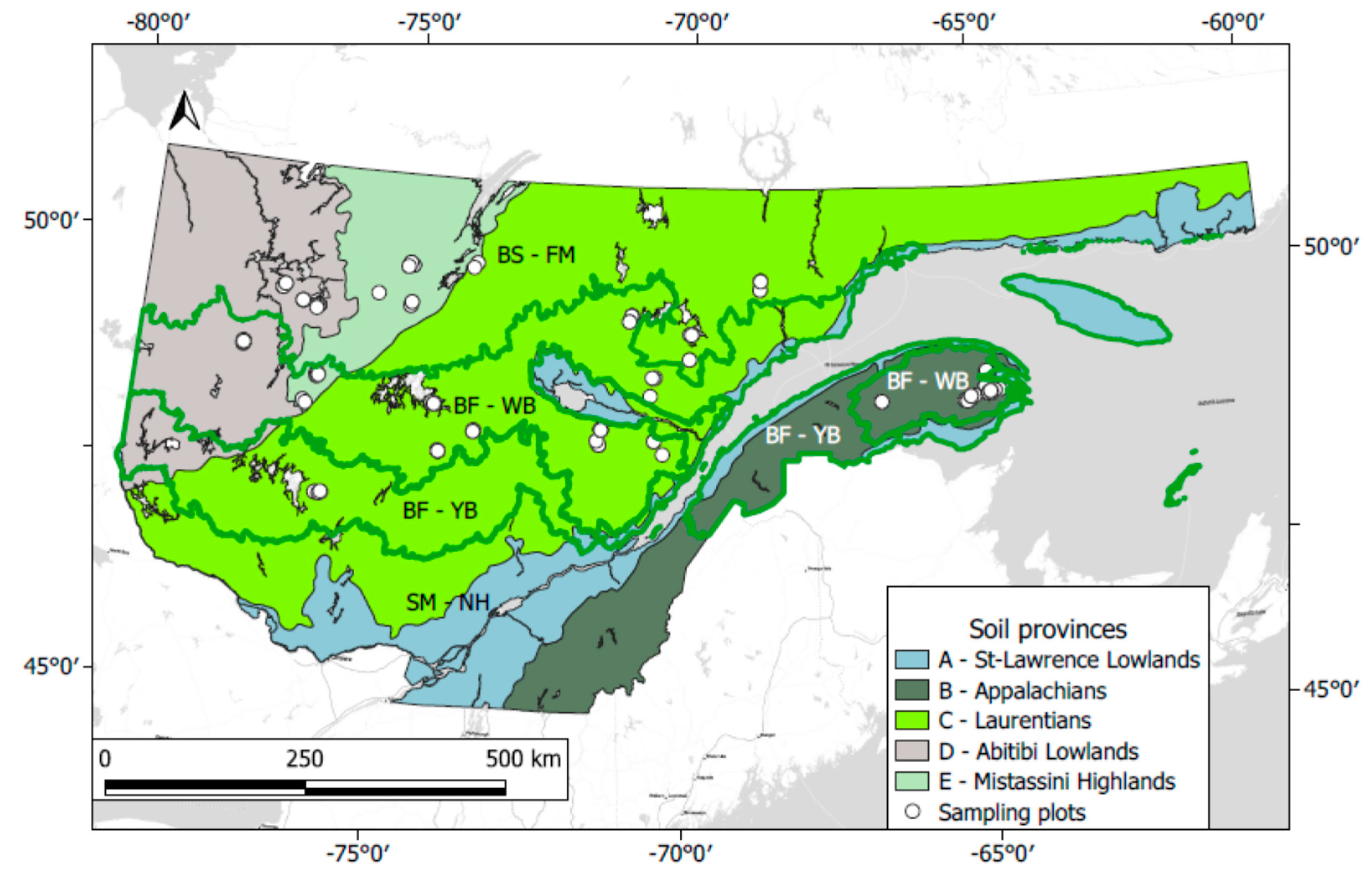
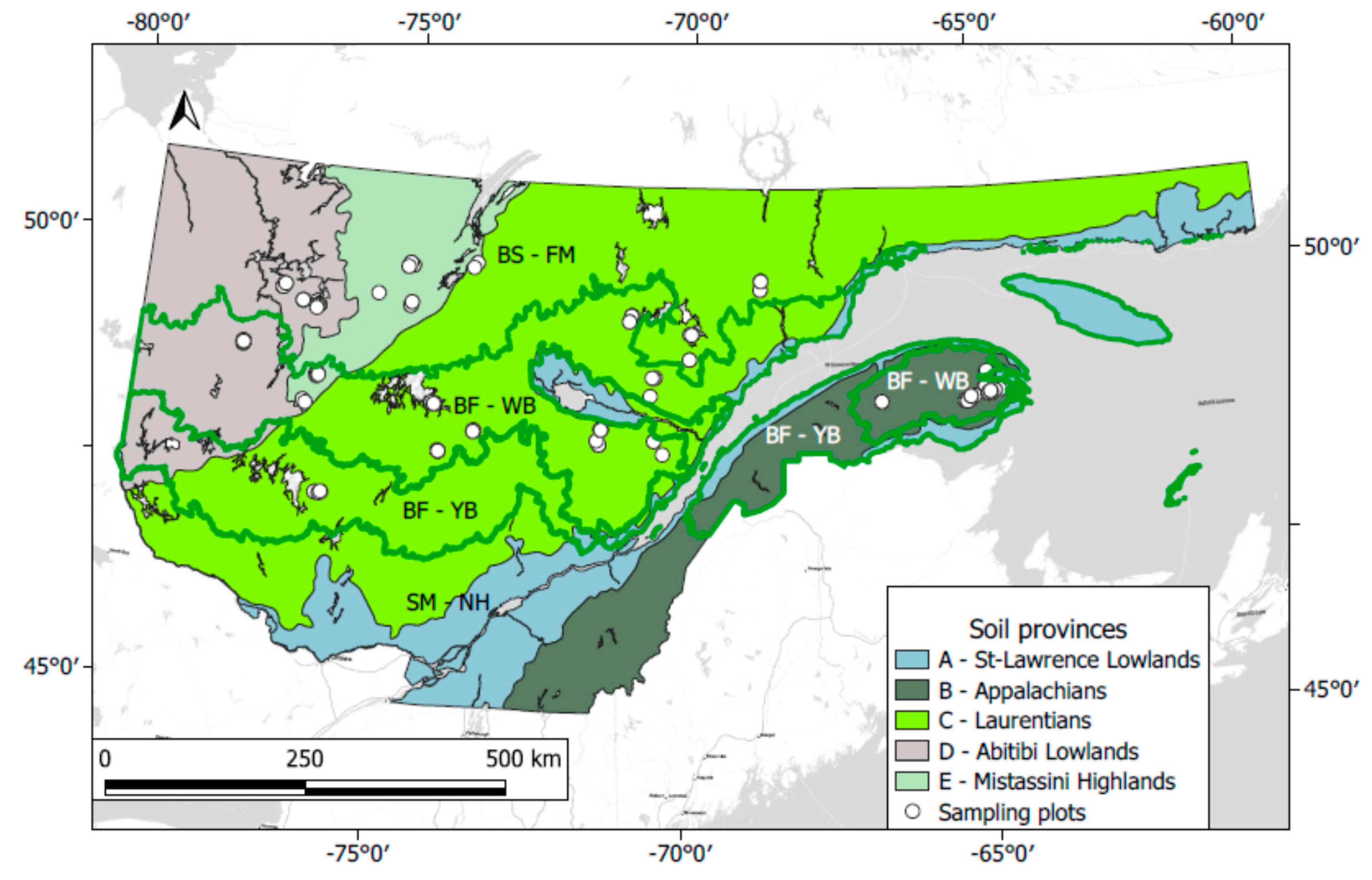
2.1. Study Area
2.2. Experimental Design
2.3. Field Sampling and Measurements
2.4. Laboratory Analyses
2.5. Computations
- OM is organic matter content (Mg·ha−1),
- M is oven-dry sample mass (g), and
- A is core sampling area (50.265 cm2).
- QH is element content in the forest floor (kg·ha−1), and
- [x] is element concentration in the forest floor (mg·kg−1).
- Qm is element content in the mineral soil (kg·ha−1),
- [x] is element concentration (mg·kg−1) on an oven-dry basis,
- Db is bulk density (g·cm−3), and
- Ee is effective thickness of the layer (cm), i.e., the corrected thickness of the soil layer without fragments (f).
- Ee is effective thickness of the layer (cm),
- E is measured thickness of the layer (cm), and
- f is the coarsest fraction of the volumetric sample (>2 mm) (%/100).
2.6. Statistical Analysis
3. Results
3.1. Harvested Biomass and Mineralomass
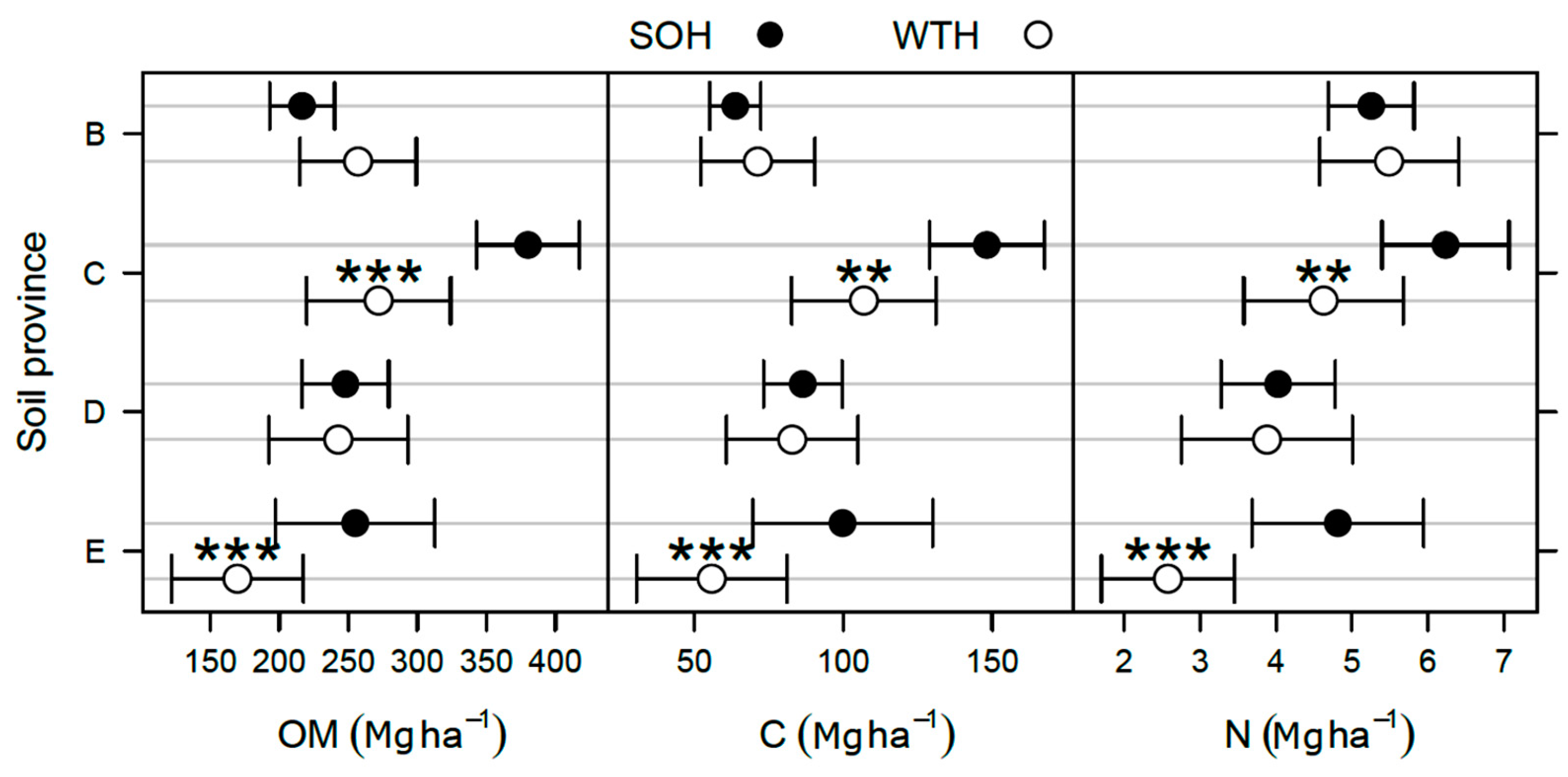
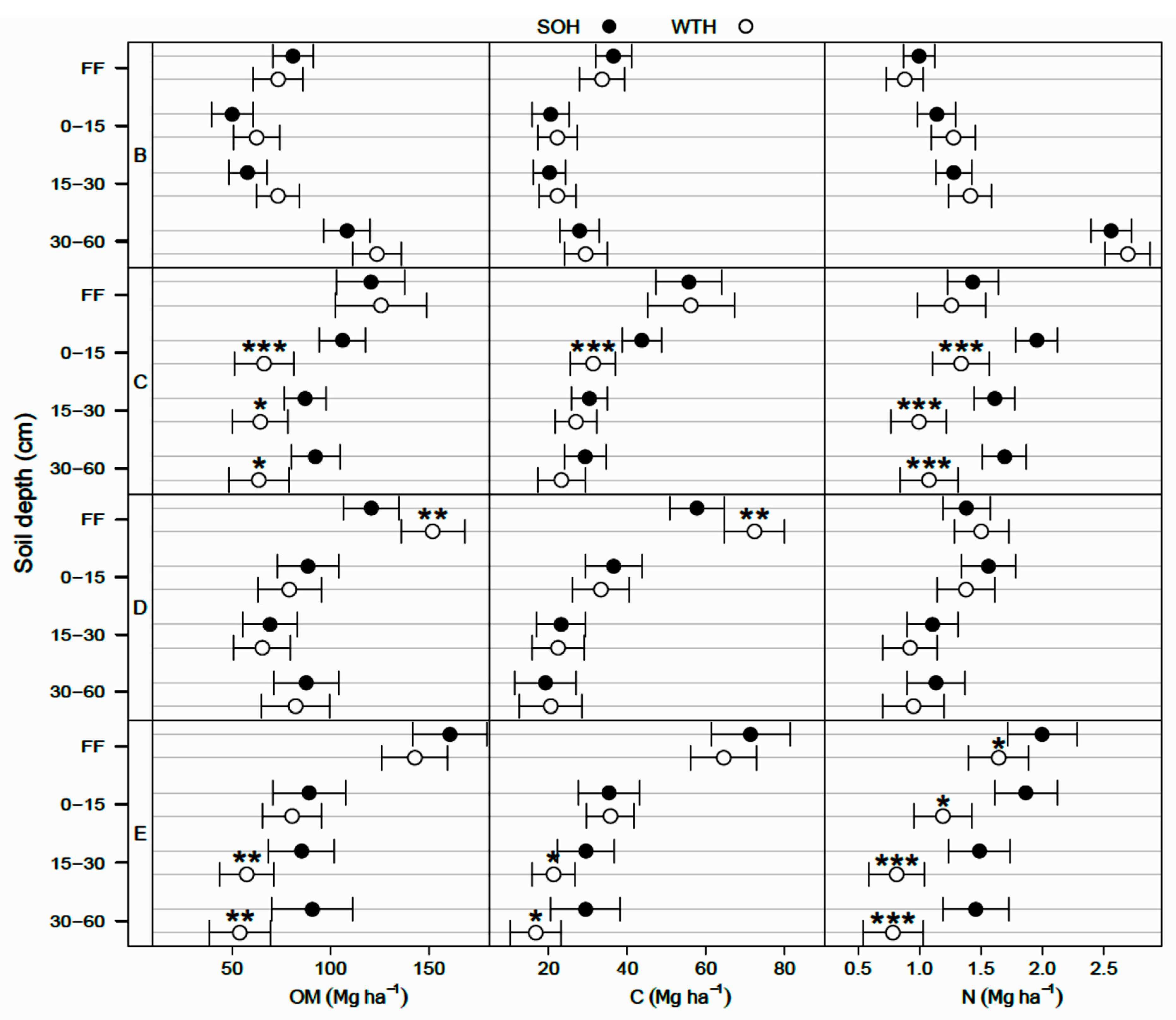
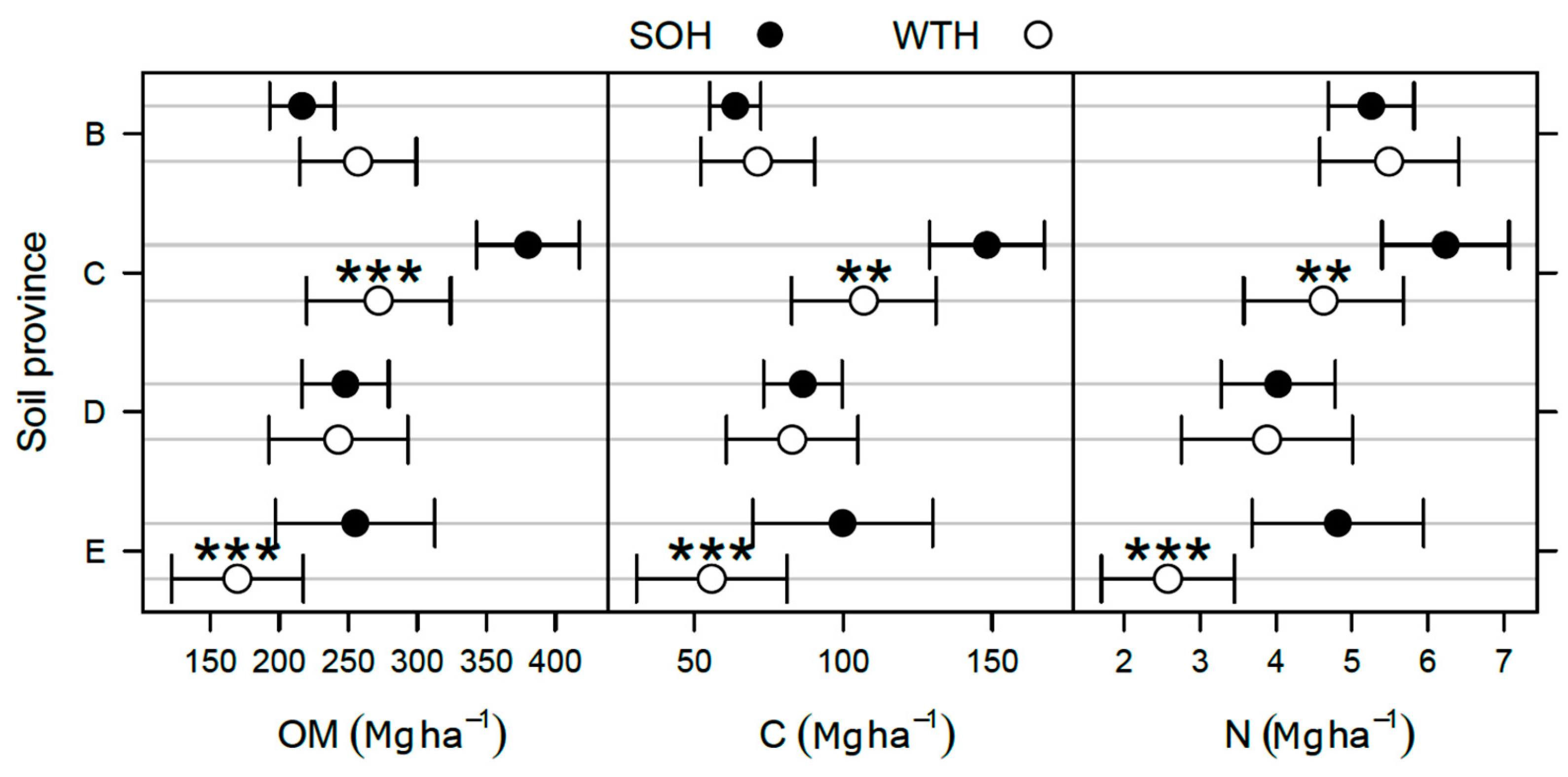
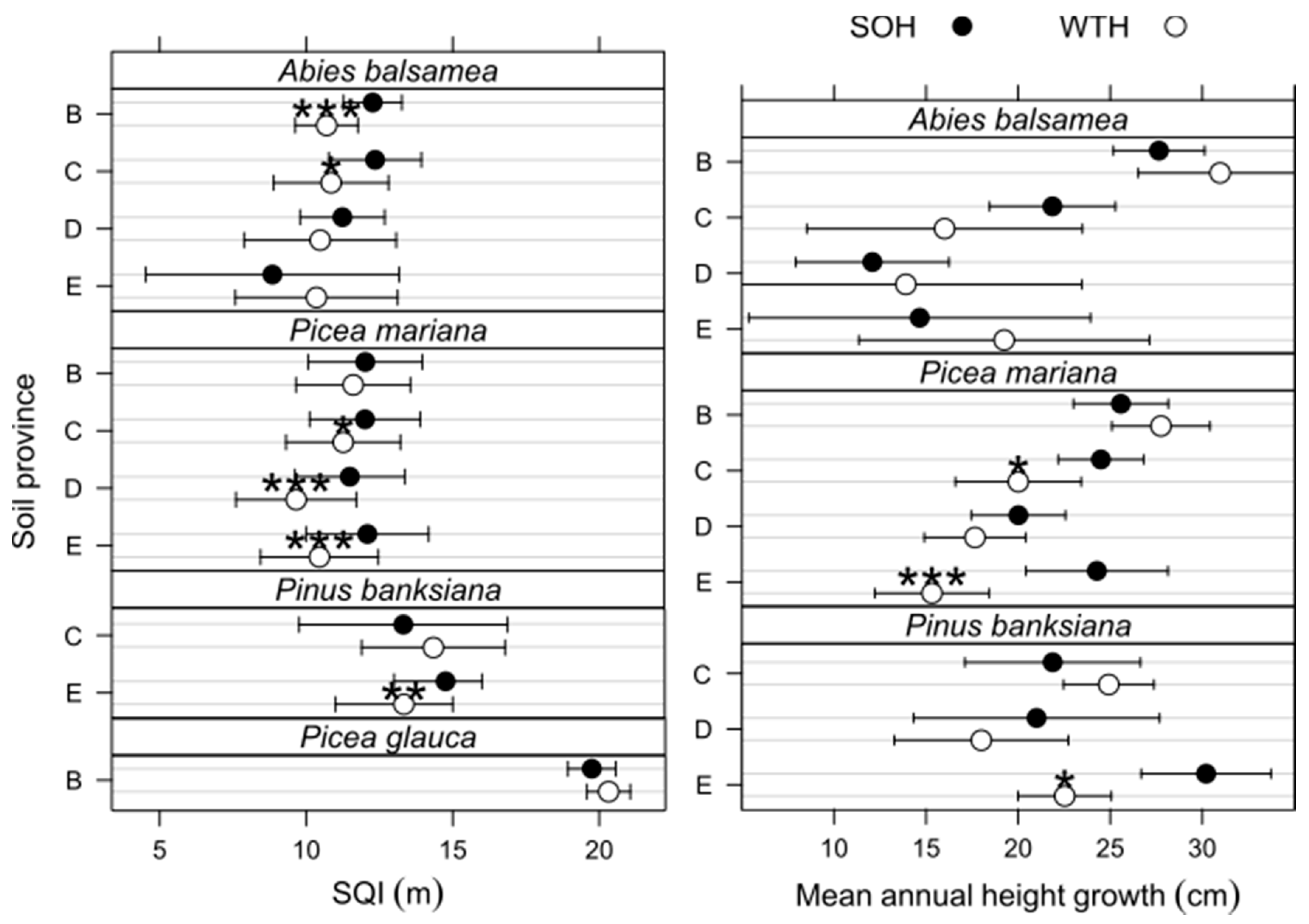
3.2. Differences in the Forest Floor after 30 Years
- -
- In the Appalachians soil province (B), the total Mn pool was 26% smaller in WTH sites (p = 0.026; Table 4).
- -
- In the Laurentians soil province (C), BS was 16% lower in WTH sites than in SOH sites (p = 0.003; Table 3).
- -
- In the Abitibi Lowlands soil province (D), OM and organic C pools were about 25% greater in WTH sites (OM: 152 ± 8 Mg·ha−1; C: 72 ± 4 Mg·ha−1) than in SOH sites (OM: 121 ± 7 Mg·ha−1; C: 58 ± 3 Mg·ha−1; p = 0.004; Figure 2). In the forest floor, total P pool was nearly 20% larger (p = 0.021; Table 4), total Mn pool was 40% smaller (p = 0.026; Table 4), and exchangeable Mg and CEC pools were respectively 41% and 28% larger in WTH sites than in SOH sites (p ≤ 0.007; Table 3). Forest floor acidity status—as expressed by BS, exchangeable Al, exchangeable Fe, and exchangeable acidity pools—was 71% to 118% greater (19% smaller in the case of BS) in WTH sites than in SOH sites (p ≤ 0.014).
- -
- In the Mistassini Highlands soil province (E), no differences in OM and organic C pools in the forest floor were detected between WTH and SOH sites (p ≥ 0.119), but total N pool was 18% smaller in WTH sites (1.64 ± 0.12 Mg·ha−1) than in SOH sites (2.00 ± 0.14 Mg·ha−1, p = 0.050; Figure 2). Total K pool in the forest floor also was 16% smaller (p = 0.018; Table 4), while total Fe pool was 35% larger in WTH sites than in SOH sites (p = 0.002). This was the only instance where total K and Fe pools in the forest floor differed between harvesting treatments.
3.3. Differences in the Mineral Soil after 30 Years
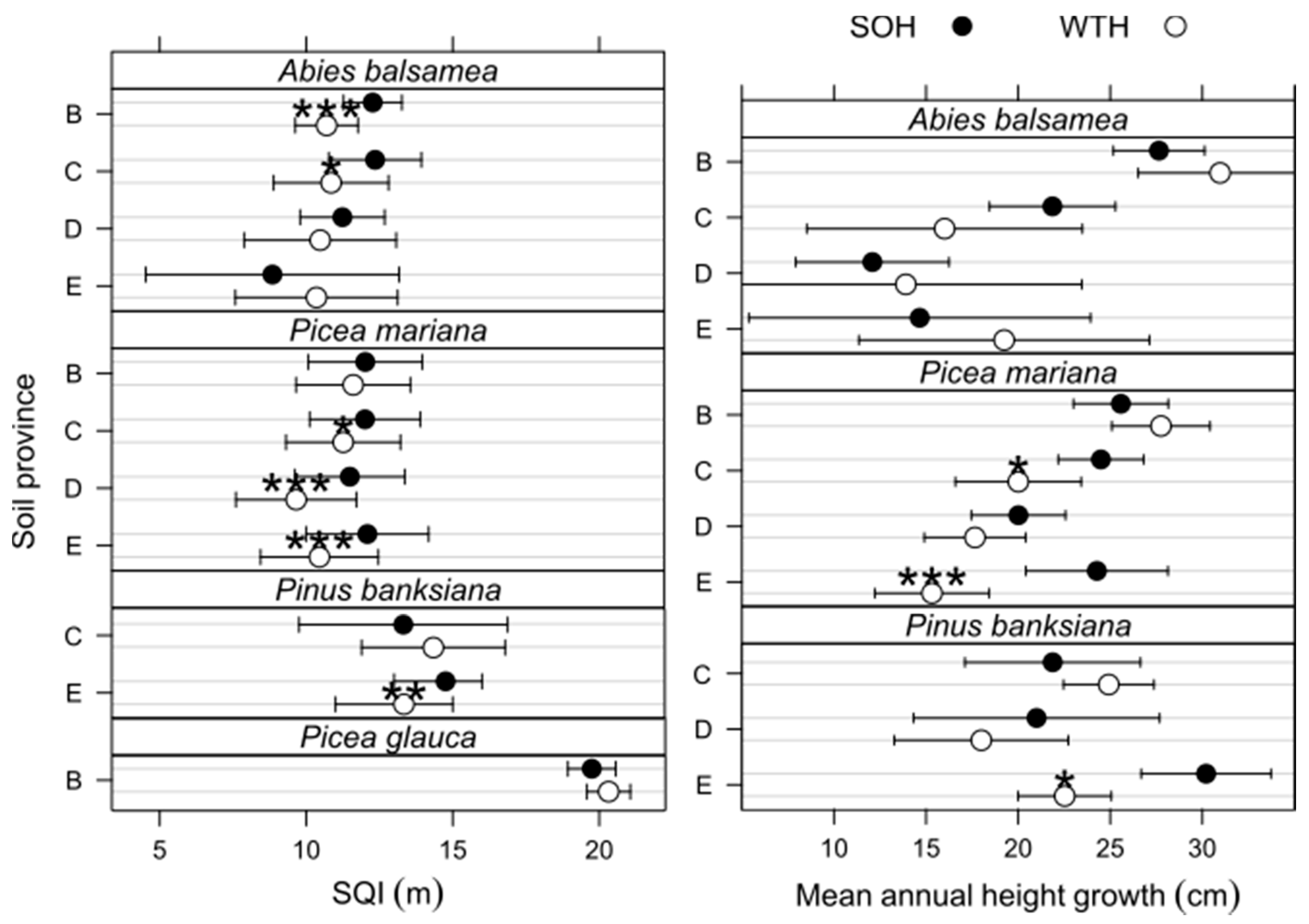
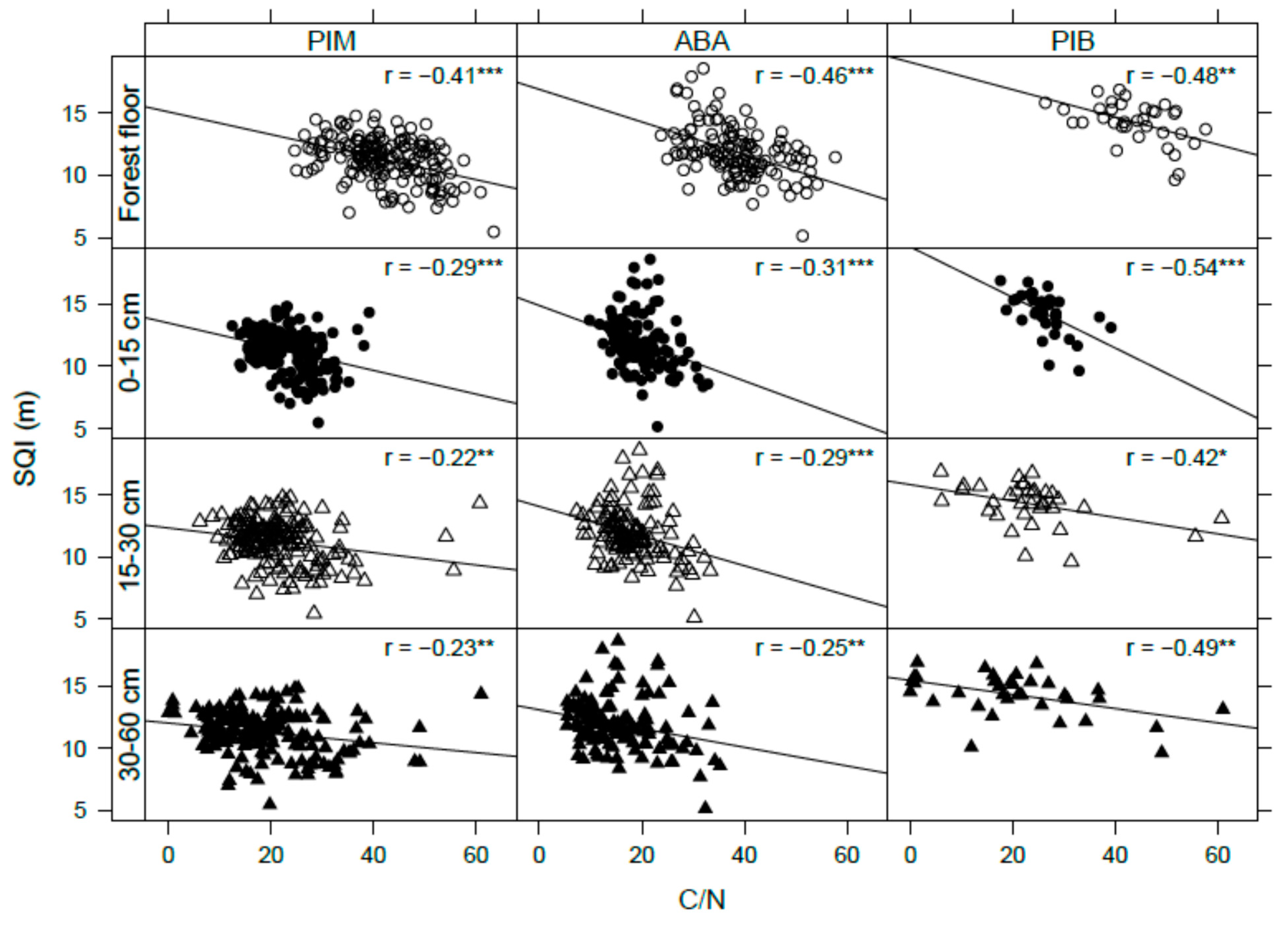
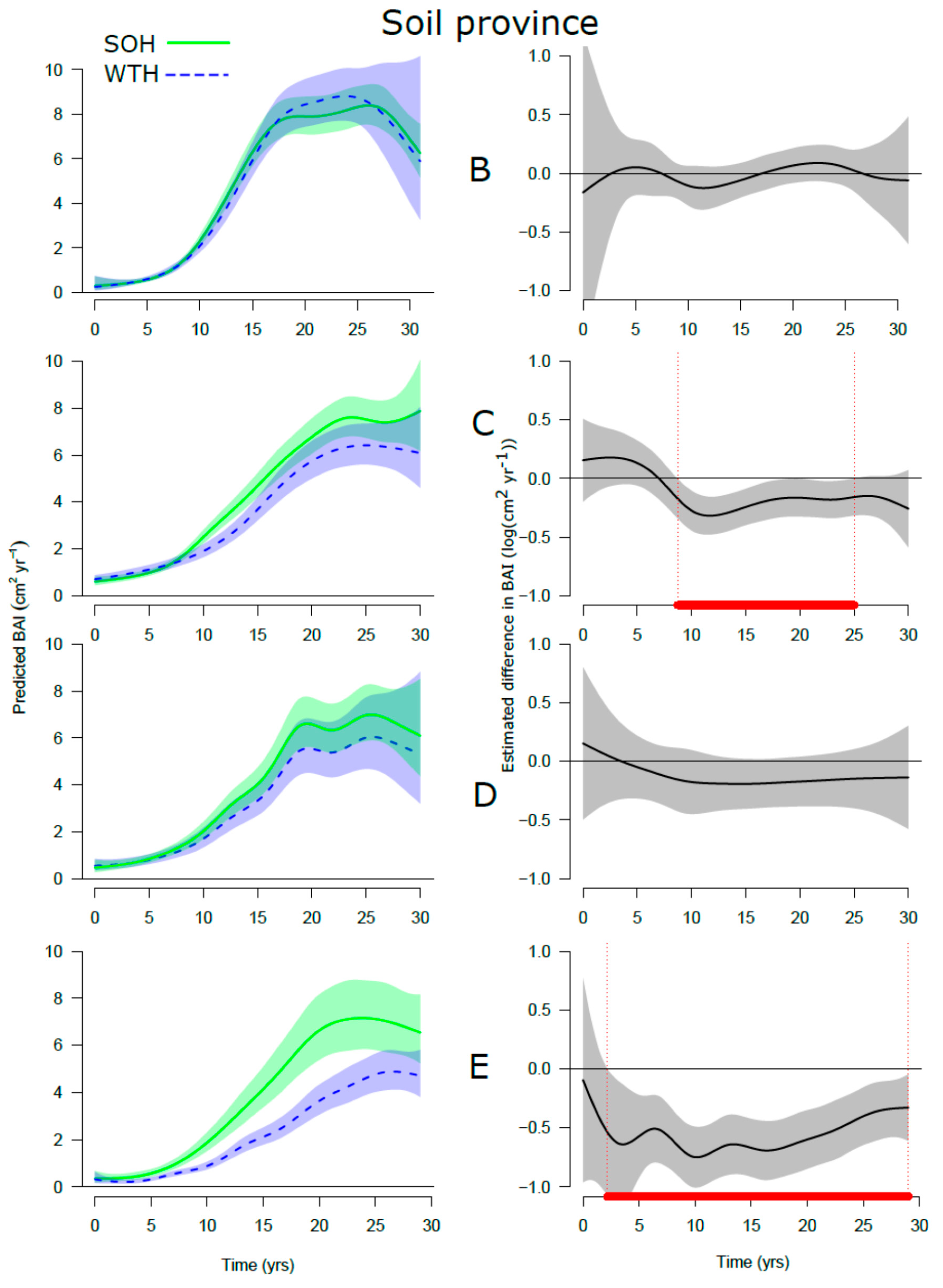
3.4. Site Productivity
4. Discussion
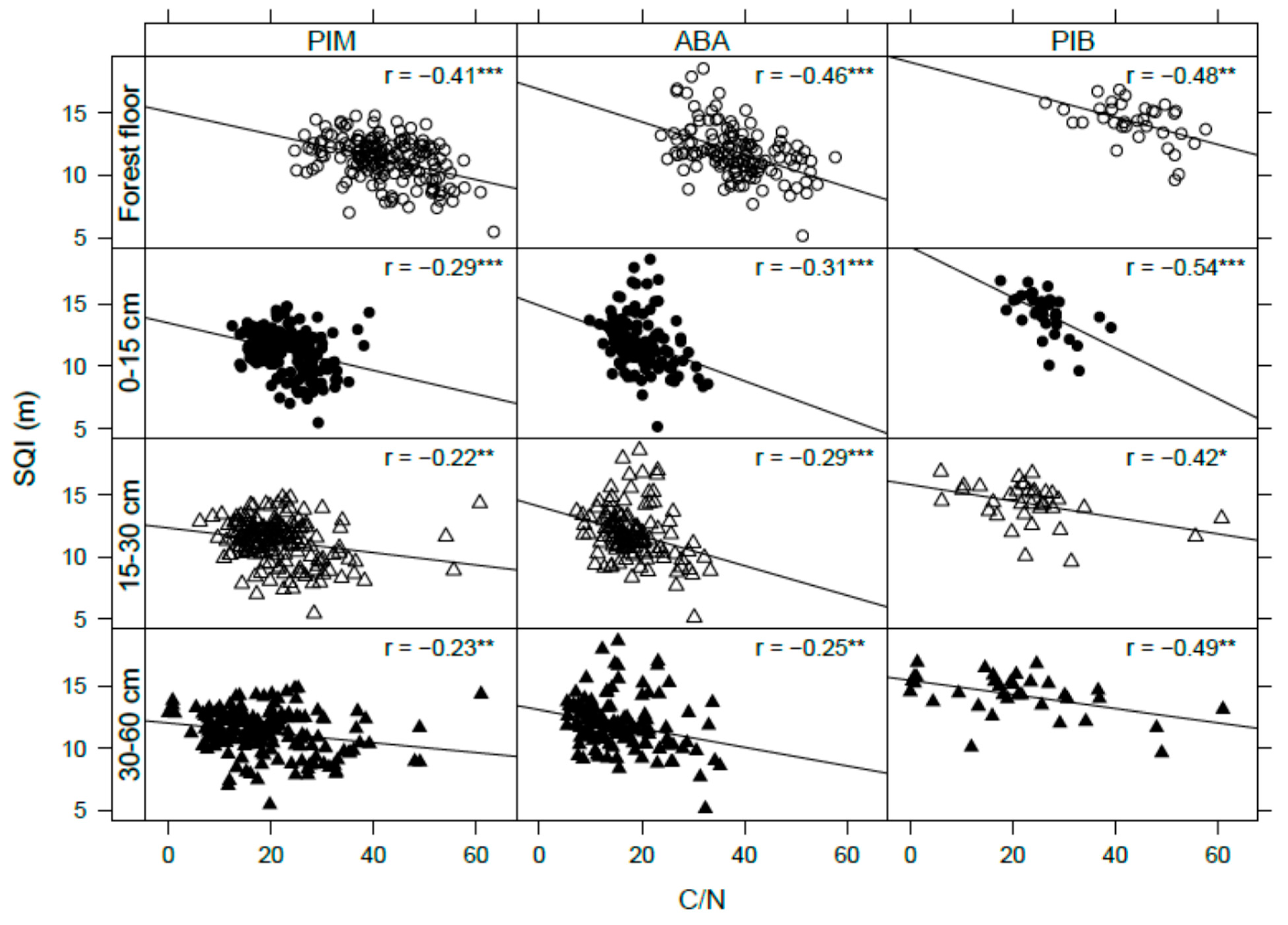
4.1. Soil Organic Carbon
4.2. Soil Nitrogen
4.3. Soil Acidification
4.4. Specificity of Stand Response to Increased Biomass Harvesting
4.5. Management and Policy Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rogelj, J.; Shindell, D.; Jiang, K.; Fifita, S.; Forster, P.; Ginzburg, V.; Handa, C.; Kheshgi, H.; Kobayashi, S.; Kriegler, E.; et al. Mitigation pathways compatible with 1.5 °C in the context of sustainable development. In Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C Above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson Delmotte, V., Zhai, P., Pörtner, H.O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Eds.; 2018; pp. 93–174. Available online: https://www.ipcc.ch/sr15/chapter/chapter-2/ (accessed on 7 April 2021).
- Delisle, J.F. Ressources et Industries Forestières du Québec. Portrait Statistique 2018; Ministère des Forêts, de la Faune et des Parcs: Québec, QC, Canada, 2019; p. 132.
- Beauregard, R.; Lavoie, P.; Thiffault, É.; Ménard, I.; Moreau, L.; Boucher, J.; François; Robichaud, F. Rapport du Groupe de Travail sur la Forêt et les Changements Climatiques; Ministère des Forêts, de la Faune et des Parcs: Québec, QC, Canada, 2019; p. 53.
- Thiffault, E.; Paré, D.; Brais, S.; Titus, B.D. Intensive biomass removals and site productivity in Canada: A review of relevant issues. For. Chron. 2010, 86, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Thiffault, E.; Hannam, K.D.; Paré, D.; Titus, B.D.; Hazlett, P.W.; Maynard, D.G.; Brais, S. Effects of forest biomass harvesting on soil productivity in boreal and temperate forests—A review. Environ. Rev. 2011, 19, 278–309. [Google Scholar] [CrossRef]
- Marshall, V.G. Impacts of forest harvesting on biological processes in northern forest soils. For. Ecol. Manag. 2000, 133, 43–60. [Google Scholar] [CrossRef]
- Wall, A.; Hytönen, J. The long-term effects of logging residue removal on forest floor nutrient capital, foliar chemistry and growth of a Norway spruce stand. Biomass Bioenergy 2011, 35, 3328–3334. [Google Scholar] [CrossRef]
- Helmisaari, H.-S.; Hanssen, K.H.; Jacobson, S.; Kukkola, M.; Luiro, J.; Saarsalmi, A.; Tamminen, P.; Tveite, B. Logging residue removal after thinning in Nordic boreal forests: Long-term impact on tree growth. For. Ecol. Manag. 2011, 261, 1919–1927. [Google Scholar] [CrossRef]
- Grigal, D.F. Effects of extensive forest management on soil productivity. Forest Ecol. Manag. 2000, 138, 167–185. [Google Scholar] [CrossRef]
- Wall, A. Risk analysis of effects of whole-tree harvesting on site productivity. For. Ecol. Manag. 2012, 282, 175–184. [Google Scholar] [CrossRef]
- Jerabkova, L.; Prescott, C.E.; Titus, B.D.; Hope, G.D.; Walters, M.B. A meta-analysis of the effects of clearcut and variable-retention harvesting on soil nitrogen fluxes in boreal and temperate forests. Can. J. For. Res. 2011, 41, 1852–1870. [Google Scholar] [CrossRef]
- Achat, D.L.; Deleuze, C.; Landmann, G.; Pousse, N.; Ranger, J.; Augusto, L. Quantifying consequences of removing harvesting residues on forest soils and tree growth-A meta-analysis. For. Ecol. Manag. 2015, 348, 124–141. [Google Scholar] [CrossRef]
- Ranius, T.; Hämäläinen, A.; Egnell, G.; Olsson, B.; Eklöf, K.; Stendahl, J.; Rudolphi, J.; Sténs, A.; Felton, A. The effects of logging residue extraction for energy on ecosystem services and biodiversity: A synthesis. J. Environ. Manag. 2018, 209, 409–425. [Google Scholar] [CrossRef]
- Brandtberg, P.-O.; Olsson, B.A. Changes in the effects of whole-tree harvesting on soil chemistry during 10years of stand development. For. Ecol. Manag. 2012, 277, 150–162. [Google Scholar] [CrossRef]
- Zetterberg, T.; Olsson, B.A.; Löfgren, S.; Hyvönen, R.; Brandtberg, P.O. Long-term soil calcium depletion after conventional and whole-tree harvest. For. Ecol. Manag. 2016, 369, 102–115. [Google Scholar] [CrossRef]
- Johnson, C.E.; Johnson, A.H.; Huntington, T.G.; Siccama, T.G. Whole-tree clear-cutting effects on soil horizons and organic-matter pools. Soil Sci. Soc. Am. J. 1991, 55, 497–502. [Google Scholar] [CrossRef]
- Olsson, B.A.; Bengtsson, J.; Lundkvist, H. Effects of different forest harvest intensities on the pools of exchangeable cations in coniferous forest soils. For. Ecol. Manag. 1996, 84, 135–147. [Google Scholar] [CrossRef]
- Morris, D.M.; Hazlett, P.W.; Fleming, R.L.; Kwiaton, M.M.; Hawdon, L.A.; Leblanc, J.D.; Primavera, M.J.; Weldon, T.P. Effects of biomass removal levels on soil carbon and nutrient reserves in conifer-dominated, coarse-textured sites in Northern Ontario: 20-year results. Soil Sci. Soc. Am. J. 2019, 83, S116–S132. [Google Scholar] [CrossRef] [Green Version]
- Tamminen, P.; Saarsalmi, A. Effects of whole-tree harvesting on growth of pine and spruce seedlings in southern Finland. Scand. J. For. Res. 2013, 28, 559–565. [Google Scholar] [CrossRef]
- Abbas, D.; Current, D.; Phillips, M.; Rossman, R.; Hoganson, H.; Brooks, K.N. Guidelines for harvesting forest biomass for energy: A synthesis of environmental considerations. Biomass Bioenergy 2011, 35, 4538–4546. [Google Scholar] [CrossRef]
- Clarke, N.; Gundersen, P.; Jönsson-Belyazid, U.; Kjønaas, O.J.; Persson, T.; Sigurdsson, B.D.; Stupak, I.; Vesterdal, L. Influence of different tree-harvesting intensities on forest soil carbon stocks in boreal and northern temperate forest ecosystems. For. Ecol. Manag. 2015, 351, 9–19. [Google Scholar] [CrossRef]
- MFFP. Guide to the Application of the Regulation Respecting the Sustainable Development of Forest in the Domain of the State; Gouvernement du Québec, Ministère des Forêts, de la Faune et des Parcs: Québec, QC, Canada, 2020; p. 301.
- Lisowska Mieszkowska, E. UNECE convention on long-range transboundary air pollution–40 years of action for cleaner air. Ekonomia Środowisko 2020, 1, 168–179. [Google Scholar]
- de Oliveira Garcia, W.; Amann, T.; Hartmann, J. Increasing biomass demand enlarges negative forest nutrient budget areas in wood export regions. Sci. Rep. 2018, 8, 5280. [Google Scholar] [CrossRef]
- Akselsson, C.; Belyazid, S. Critical biomass harvesting–Applying a new concept for Swedish forest soils. For. Ecol. Manag. 2018, 409, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Paré, D.; Thiffault, E. Nutrient budgets in forests under increased biomass harvesting scenarios. Curr. For. Rep. 2016, 2, 81–91. [Google Scholar] [CrossRef] [Green Version]
- James, J.; Harrison, R. The effect of harvest on forest soil carbon: A meta-analysis. Forests 2016, 7, 308. [Google Scholar] [CrossRef]
- Bowd, E.J.; Banks, S.C.; Strong, C.L.; Lindenmayer, D.B. Long-term impacts of wildfire and logging on forest soils. Nat. Geosci. 2019, 12, 113–118. [Google Scholar] [CrossRef]
- Lamontagne, L.; Nolin, M.C. Cadre Pédologique de Référence Pour la Corrélation Des Sols; Équipe pédologique du Québec, Centre de Recherche et de Développement sur les Sols et les Grandes Cultures, Agriculture et Agroalimentaire Canada: Sainte-Foy, QC, Canada, 1997; p. 69. [Google Scholar]
- Soil Classification Working Group. The Canadian System of Soil Classification, 3rd ed.; Agriculture and Agri-Food Canada: Ottawa, ON, Canada, 1998; p. 187.
- New, M.; Lister, D.; Hulme, M.; Makin, I. A high-resolution data set of surface climate over global land areas. Clim. Res. 2002, 21, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Trabucco, A.; Zomer, R. Global Aridity Index and Potential Evapotranspiration (ET0) Climate Database v2; CGIAR-CSI: 2019; Available online: https://cgiarcsi.community/2019/01/24/global-aridity-index-and-potential-evapotranspiration-climate-database-v2 (accessed on 7 April 2021).
- Pothier, D. Évolution de la régénération après la coupe de peuplements récoltés selon différents procédés d’exploitation. For. Chron. 1996, 72, 519–527. [Google Scholar] [CrossRef] [Green Version]
- Saucier, J.P.; Berger, J.P.; D’Avignon, H.; Racine, P. Le Point D’observation Écologique: Normes Techniques; Service des Inventaires Forestiers, Direction de la Gestion des Stocks Forestiers, Ministère des Ressources Naturelles du Québec: Québec, QC, Canada, 1994; p. 116.
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Chicago Press: Chicago, IL, USA, 1968; p. 110. [Google Scholar]
- Yamaguchi, D.K. A simple method for cross-dating increment cores from living trees. Can. J. For. Res. 1991, 21, 414–416. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree Rings and Climate; Blackburn Press: Caldwell, NC, USA, 2001. [Google Scholar]
- Grissino Mayer, H.D. Evaluating crossdating accuracy: A manual and tutorial for the computer program COFECHA. Tree Ring Res. 2001, 57, 205–221. [Google Scholar]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Duchesne, L.; Ouimet, R.; Morneau, C. Assessment of sugar maple health based on basal area growth pattern. Can. J. For. Res. 2003, 33, 2074–2080. [Google Scholar] [CrossRef]
- Paré, D.; Bernier, P.; Lafleur, B.; Titus, B.D.; Thiffault, E.; Maynard Doug, G.; Guo, X. Estimating stand-scale biomass, nutrient contents, and associated uncertainties for tree species of Canadian forests. Can. J. For. Res. 2013, 43, 599–608. [Google Scholar] [CrossRef]
- Federer, C.A.; Turcotte, D.E.; Smith, C.T. The organic fraction-bulk density relationship and the expression of nutrient content in forest soils. Can. J. For. Res. 1993, 23, 1026–1032. [Google Scholar] [CrossRef]
- Beaudette, D.E.; Roudier, P.; O’Geen, A.T. Algorithms for quantitative pedology: A toolkit for soil scientists. Comput. Geosci. 2013, 52, 258–268. [Google Scholar] [CrossRef]
- Pothier, D.; Savard, F. Actualisation des Tables de Production Pour les Principales Espèces Forestière du Québec; Gouvernement du Québec. Ministère des Ressources naturelles: Québec, QC, Canada, 1998; p. 183.
- Prégent, G.; Picher, G.; Auger, I. Tarif de Cubage, Tables de Rendement et Modèles de Croissance Pour les Plantations D’épinette Blanche au Québec; Mémoire de recherche forestière n° 160; Direction de la Recherche Forestière, Ministère des Ressources et de la Faune du Québec: Québec, QC, Canada, 2010; p. 73.
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means; R Package Version 1.4.2; 2019; Available online: https://CRAN.R-project.org/package=emmeans (accessed on 7 April 2021).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models; R Package Version 3.1-150; 2020; Available online: https://CRAN.R-project.org/package=nlme (accessed on 7 April 2021).
- Wood, S.N. Mgcv: Mixed GAM Computation Vehicle with Automatic Smoothness Estimation (Version 1.8-22); 2017; Available online: https://CRAN.R-project.org/package=mgcv (accessed on 7 April 2021).
- Van Rij, J.; Wieling, M.; Baayen, R.H.; Van Rijn, H. Itsadug: Interpreting time series and autocorrelated data using GAMMs. R Package Version 2017, 2. Available online: https://CRAN.R-project.org/package=itsadug (accessed on 7 April 2021).
- Pichler, V.; Gömöryová, E.; Leuschner, C.; Homolák, M.; Abrudan, I.V.; Pichlerová, M.; Střelcová, K.; Di Filippo, A.; Sitko, R. Parent material effect on soil organic carbon concentration under primeval european beech forests at a regional scale. Forests 2021, 12, 405. [Google Scholar] [CrossRef]
- Hassink, J. The capacity of soils to preserve organic C and N by their association with clay and silt particles. Plant Soil 1997, 191, 77–87. [Google Scholar] [CrossRef]
- Spielvogel, S.; Prietzel, J.; Kögel-Knabner, I. Soil organic matter stabilization in acidic forest soils is preferential and soil type-specific. Eur. J. Soil Sci. 2008, 59, 674–692. [Google Scholar] [CrossRef]
- Thiffault, E.; Paré, D.; Bélanger, N.; Munson, A.D.; Marquis, F. Harvesting intensity at clear-felling in the boreal forest: Impact on soil and foliar nutrient status. Soil Sci. Soc. Am. J. 2006, 70, 691–701. [Google Scholar] [CrossRef]
- Hamburg, S.P.; Vadeboncoeur, M.A.; Johnson, C.E.; Sanderman, J. Losses of mineral soil carbon largely offset biomass accumulation 15 years after whole-tree harvest in a northern hardwood forest. Biogeochemistry 2019, 144, 1–14. [Google Scholar] [CrossRef]
- Kaarakka, L.; Tamminen, P.; Saarsalmi, A.; Kukkola, M.; Helmisaari, H.-S.; Burton, A.J. Effects of repeated whole-tree harvesting on soil properties and tree growth in a Norway spruce (Picea abies (L.) Karst.) stand. For. Ecol. Manag. 2014, 313, 180–187. [Google Scholar] [CrossRef]
- Achat, D.L.; Fortin, M.; Landmann, G.; Ringeval, B.; Augusto, L. Forest soil carbon is threatened by intensive biomass harvesting. Sci. Rep. 2015, 5, 15991. [Google Scholar] [CrossRef]
- Fenton, N.; Lecomte, N.; Légaré, S.; Bergeron, Y. Paludification in black spruce (Picea mariana) forests of eastern Canada: Potential factors and management implications. For. Ecol. Manag. 2005, 213, 151–159. [Google Scholar] [CrossRef]
- Prescott, C.; Maynard, D.; Laiho, R. Humus in northern forests: Friend or foe? For. Ecol. Manage 2000, 133, 23–36. [Google Scholar] [CrossRef]
- Hazlett, P.W.; Gordon, A.M.; Voroney, R.P.; Sibley, P.K. Impact of harvesting and logging slash on nitrogen and carbon dynamics in soils from upland spruce forests in northeastern Ontario. Soil Biol. Biochem. 2007, 39, 43–57. [Google Scholar] [CrossRef] [Green Version]
- Kalbitz, K.; Glaser, B.; Bol, R. Clear-cutting of a Norway spruce stand: Implications for controls on the dynamics of dissolved organic matter in the forest floor. Eur. J. Soil Sci. 2004, 55, 401–413. [Google Scholar] [CrossRef]
- Jacobson, S.; Kukkola, M.; Mälkönen, E.; Tveite, B. Impact of whole-tree harvesting and compensatory fertilization on growth of coniferous thinning stands. For. Ecol. Manag. 2000, 129, 41–51. [Google Scholar] [CrossRef]
- Egnell, G. Is the productivity decline in Norway spruce following whole-tree harvesting in the final felling in boreal Sweden permanent or temporary? For. Ecol. Manag. 2011, 261, 148–153. [Google Scholar] [CrossRef]
- Ouimet, R.; Duchesne, L. Évaluation des Types Écologiques Forestiers Sensibles à L’appauvrissement Des sols en Minéraux par la Récolte de Biomasse; Rapport hors Série; Direction de la Recherche Forestière, Ministère des Ressources Naturelles et de la Faune: Québec, QC, Canada, 2009; p. 25.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Soil Province | |||
|---|---|---|---|---|
| B (Appalachians) | C (Laurentians) | D (Abitibi Lowlands) | E (Mistassini Highlands) | |
| Main parent material | Glacial till or regolith made of sedimentary and metamorphic rock | Glacial till made of igneous and metamorphic rocks of the Canadian Shield | Extensive glaciolacustrine clay deposit, glaciofluvial sand, and gravel deposits over clay | Glacial till made of igneous, metamorphic, and volcanic rock |
| Geology | Sandstone, siltstone, shale, slate | Granite, gneiss | Interlayered clays | Granite, gneiss |
| Soils * | Brunisols to Podzols | Podzols | Gleysoils, Gleyic Podzols to Podzols | Podzols |
| Soil texture | Medium (loam) | Coarse (sand to sandy loam) | Fine (clay or silt) | Coarse (sand to sandy loam) |
| Altitude (m) | 180–1300 | 180–1200 | 30–525 | 300–600 |
| Topography | Alternating ridgelines and valleys | Rounded mountains, overgrown valleys | Low-shield terrane | Rounded hills and valleys |
| January temperature † (IQR, °C) | −2.9–−14.1 | −16.0–−18.4 | −17.3–−18.6 | −17.8–−18.5 |
| July temperature † (IQR, °C) | 16.1–17.2 | 15.8–16.8 | 16.3–16.8 | 16.2–16.5 |
| Annual precipitations † (IQR, mm) | 1054–1113 | 964–1067 | 891–919 | 913–952 |
| Degree-days † (IQR) | 1180–1350 | 950–1290 | 1030–1190 | 990–1110 |
| Aridity index ‡ (IQR) | 1.41–1.64 | 1.34–1.52 | 1.19–1.26 | 1.27–1.33 |
| Nb. frost-free days † (d) | 150 | 140 | 140 | 142 |
| Vegetation | Mixed forest to balsam fir–white birch forest | Balsam fir–white birch to black spruce–feather moss forest | Balsam fir–white birch to black spruce–feather moss forest | Black spruce–feather moss forest |
| Soil Province * | Harvesting Treatment | n | Biomass | N | P | K | Ca | Mg |
|---|---|---|---|---|---|---|---|---|
| (Mg·ha−1) | (kg·ha−1) | |||||||
| B | SOH | 39 | 82 ± 5 | 112 ± 10 | 12 ± 1 | 83 ± 6 | 167 ± 15 | 21 ± 2 |
| WTH | 25 | 138 ± 6 | 399 ± 13 | 50 ± 2 | 228 ± 8 | 472 ± 19 | 62 ± 2 | |
| C | SOH | 39 | 70 ± 5 | 94 ± 10 | 11 ± 1 | 63 ± 6 | 182 ± 15 | 20 ± 2 |
| WTH | 30 | 73 ± 5 | 181 ± 12 | 22 ± 1 | 95 ± 7 | 185 ± 17 | 26 ± 2 | |
| D | SOH | 21 | 40 ± 6 | 44 ± 14 | 5 ± 2 | 26 ± 9 | 98 ± 21 | 9 ± 2 |
| WTH | 12 | 71 ± 8 | 173 ± 18 | 23 ± 2 | 98 ± 11 | 206 ± 27 | 27 ± 3 | |
| E | SOH | 11 | 77 ± 6 | 87 ± 19 | 9 ± 2 | 47 ± 12 | 129 ± 28 | 16 ± 3 |
| WTH | 19 | 81 ± 8 | 184 ± 15 | 23 ± 2 | 99 ± 9 | 196 ± 22 | 27 ± 3 | |
| Global mean | SOH | 110 | 68 ± 3 | 84 ± 7 | 9 ± 1 | 55 ± 4 | 144 ± 11 | 17 ± 1 |
| WTH | 86 | 90 ± 3 | 234 ± 7 | 30 ± 1 | 130 ± 4 | 265 ± 10 | 36 ± 1 | |
| Difference | WTH—SOH | 21 ± 4 | 150 ± 10 | 20 ± 1 | 75 ± 6 | 121 ± 15 | 19 ± 2 | |
| Soil Province * | Harvesting Treatment | pH | C/N | Ca | Mg | K | Mn | Al | Fe | Exch. Acidity | CEC | BS |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Exchangeable Pool (kg·ha−1) | (keq·ha−1) | (%) | ||||||||||
| Forest Floor | ||||||||||||
| B | SOH | 4.32 ± 0.06 | 37.5 ± 1.2 | 307 ± 32 | 37 ± 3 | 80 ± 3 | 31 ± 2 | 7 ± 2 | 7 ± 2 | 1.8 ± 0.3 | 25 ± 2 | 88 ± 1 |
| WTH | 4.31 ± 0.06 | 40.3 ± 1.7 | 285 ± 39 | 37 ± 4 | 76 ± 3 | 26 ± 2 | 15 ± 5 | 8 ± 3 | 2.1 ± 0.8 | 26 ± 3 | 90 ± 2 | |
| C | SOH | 4.07 ± 0.06 | 38.7 ± 1.5 | 352 ± 35 | 49 ± 4 | 75 ± 4 | 15 ± 2 | 51 ± 6 | 17 ± 2 | 10.1 ± 1.1 | 38 ± 3 | 70 ± 2 |
| WTH | 4.05 ± 0.06 | 42.9 ± 2.2 | 329 ± 43 | 47 ± 5 | 71 ± 4 | 11 ± 2 | 60 ± 7 | 22 ± 4 | 10.1 ± 1.3 | 32 ± 4 | 59 ± 3 | |
| D | SOH | 4.13 ± 0.04 | 40.5 ± 1.6 | 412 ± 37 | 49 ± 5 | 90 ± 3 | 22 ± 2 | 62 ± 11 | 21 ± 3 | 9.4 ± 1.4 | 40 ± 3 | 72 ± 3 |
| WTH | 4.12 ± 0.06 | 48.5 ± 2.4 | 390 ± 47 | 69 ± 6 | 85 ± 3 | 10 ± 2 | 135 ± 13 | 36 ± 3 | 18.6 ± 1.9 | 51 ± 3 | 58 ± 4 | |
| E | SOH | 4.08 ± 0.06 | 35.4 ± 2.0 | 425 ± 41 | 65 ± 5 | 87 ± 4 | 4 ± 2 | 85 ± 14 | 17 ± 3 | 15.5 ± 1.9 | 46 ± 4 | 60 ± 4 |
| WTH | 4.06 ± 0.06 | 40.8 ± 1.9 | 403 ± 45 | 60 ± 4 | 83 ± 4 | 10 ± 1 | 81 ± 10 | 22 ± 3 | 13.7 ± 1.5 | 47 ± 4 | 66 ± 3 | |
| P(Soil province) | 0.006 | 0.018 | 0.015 | <0.001 | 0.002 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| P(Harvesting treatment) | 0.676 | <0.001 | 0.052 | 0.883 | 0.104 | 0.026 | 0.070 | 0.147 | 0.053 | 0.768 | 0.423 | |
| P(Soil prov. × Harvesting) | 0.989 | 0.131 | 0.360 | 0.040 | 0.068 | <0.001 | 0.003 | 0.050 | 0.002 | 0.045 | <0.001 | |
| Mineral soil 0–15 cm | ||||||||||||
| B | SOH | 4.20 ± 0.07 | 18.7 ± 0.6 | 695 ± 171 | 39 ± 4 | 32 ± 6 | 15.9 ± 2.0 | 656 ± 59 | 65 ± 3 | 75 ± 6 | 93 ± 8 | 25 ± 4 |
| WTH | 4.24 ± 0.08 | 16.9 ± 0.8 | 775 ± 234 | 37 ± 5 | 39 ± 6 | 19.5 ± 2.1 | 752 ± 72 | 57 ± 4 | 87 ± 7 | 90 ± 8 | 15 ± 5 | |
| C | SOH | 4.89 ± 0.06 | 21.6 ± 1.2 | 767 ± 161 | 38 ± 6 | 25 ± 6 | 5.7 ± 0.8 | 433 ± 36 | 67 ± 4 | 51 ± 4 | 53 ± 4 | 25 ± 4 |
| WTH | 4.93 ± 0.07 | 24.4 ± 1.6 | 847 ± 229 | 37 ± 6 | 28 ± 6 | 9.3 ± 0.8 | 252 ± 46 | 59 ± 4 | 31 ± 5 | 50 ± 5 | 32 ± 7 | |
| D | SOH | 4.81 ± 0.09 | 23.2 ± 1.0 | 724 ± 171 | 18 ± 12 | 34 ± 6 | 10.1 ± 2.0 | 488 ± 68 | 70 ± 4 | 57 ± 6 | 67 ± 17 | 28 ± 5 |
| WTH | 4.86 ± 0.10 | 24.7 ± 1.3 | 805 ± 236 | 16 ± 13 | 20 ± 8 | 13.7 ± 2.1 | 599 ± 87 | 63 ± 5 | 69 ± 8 | 63 ± 17 | 22 ± 7 | |
| E | SOH | 4.69 ± 0.07 | 20.6 ± 1.9 | 655 ± 164 | 28 ± 5 | 30 ± 7 | 3.8 ± 0.8 | 417 ± 43 | 56 ± 4 | 49 ± 4 | 49 ± 6 | 21 ± 5 |
| WTH | 4.73 ± 0.08 | 27.5 ± 1.7 | 736 ± 232 | 27 ± 5 | 21 ± 7 | 7.4 ± 0.7 | 372 ± 40 | 49 ± 4 | 43 ± 4 | 46 ± 6 | 23 ± 7 | |
| Mineral soil 15–30 cm | ||||||||||||
| B | SOH | 4.63 ± 0.07 | 15.9 ± 0.6 | 668 ± 171 | 35 ± 4 | 33 ± 8 | 9.8 ± 1.4 | 462 ± 42 | 27 ± 3 | 52 ± 6 | 59 ± 9 | 22 ± 4 |
| WTH | 4.64 ± 0.08 | 15.1 ± 0.9 | 749 ± 234 | 33 ± 5 | 43 ± 8 | 13.4 ± 1.5 | 446 ± 52 | 20 ± 3 | 51 ± 7 | 62 ± 9 | 21 ± 6 | |
| C | SOH | 5.32 ± 0.06 | 20.8 ± 1.4 | 745 ± 161 | 34 ± 5 | 7 ± 8 | 5.4 ± 0.8 | 236 ± 29 | 29 ± 3 | 28 ± 4 | 30 ± 5 | 46 ± 4 |
| WTH | 5.33 ± 0.07 | 23.3 ± 1.8 | 826 ± 229 | 32 ± 6 | 14 ± 8 | 9.0 ± 0.8 | 180 ± 40 | 22 ± 4 | 21 ± 5 | 33 ± 6 | 58 ± 7 | |
| D | SOH | 5.25 ± 0.09 | 21.1 ± 1.1 | 800 ± 171 | 13 ± 12 | 37 ± 9 | 6.7 ± 1.3 | 314 ± 48 | 33 ± 3 | 35 ± 7 | 69 ± 18 | 44 ± 5 |
| WTH | 5.25 ± 0.10 | 21.5 ± 1.5 | 881 ± 236 | 12 ± 13 | 27 ± 10 | 10.3 ± 1.5 | 341 ± 61 | 25 ± 4 | 39 ± 8 | 72 ± 18 | 47 ± 8 | |
| E | SOH | 5.12 ± 0.08 | 19.9 ± 2.3 | 688 ± 165 | 24 ± 4 | 16 ± 9 | 3.9 ± 0.7 | 234 ± 33 | 19 ± 3 | 27 ± 4 | 22 ± 7 | 34 ± 6 |
| WTH | 5.13 ± 0.08 | 26.3 ± 2.0 | 769 ± 231 | 22 ± 5 | 11 ± 9 | 7.5 ± 0.6 | 120 ± 33 | 12 ± 3 | 14 ± 4 | 25 ± 7 | 59 ± 7 | |
| Mineral soil 30–60 cm | ||||||||||||
| B | SOH | 4.85 ± 0.07 | 12.0 ± 0.8 | 890 ± 171 | 43 ± 5 | 96 ± 19 | 22.7 ± 2.2 | 713 ± 53 | 20 ± 3 | 79 ± 6 | 89 ± 29 | 24 ± 4 |
| WTH | 4.87 ± 0.08 | 11.0 ± 1.1 | 970 ± 235 | 41 ± 6 | 130 ± 20 | 26.3 ± 2.2 | 604 ± 64 | 13 ± 3 | 68 ± 7 | 93 ± 30 | 33 ± 7 | |
| C | SOH | 5.55 ± 0.06 | 18.1 ± 1.8 | 841 ± 161 | 42 ± 7 | 7 ± 19 | 6.4 ± 0.9 | 235 ± 33 | 22 ± 3 | 27 ± 4 | 25 ± 14 | 56 ± 5 |
| WTH | 5.56 ± 0.07 | 20.3 ± 2.2 | 921 ± 229 | 40 ± 8 | 24 ± 20 | 10.0 ± 0.9 | 175 ± 43 | 14 ± 4 | 20 ± 5 | 30 ± 17 | 69 ± 8 | |
| D | SOH | 5.47 ± 0.09 | 19.9 ± 1.5 | 1212 ± 172 | 21 ± 13 | 95 ± 24 | 8.8 ± 2.2 | 238 ± 60 | 25 ± 3 | 26 ± 7 | 164 ± 61 | 63 ± 6 |
| WTH | 5.48 ± 0.10 | 18.1 ± 1.9 | 1293 ± 237 | 20 ± 14 | 109 ± 26 | 12.4 ± 2.3 | 209 ± 76 | 18 ± 4 | 23 ± 8 | 169 ± 62 | 66 ± 9 | |
| E | SOH | 5.34 ± 0.08 | 18.2 ± 3.0 | 809 ± 165 | 32 ± 6 | 3 ± 24 | 4.4 ± 0.8 | 270 ± 40 | 12 ± 3 | 31 ± 4 | 9 ± 21 | 42 ± 7 |
| WTH | 5.36 ± 0.08 | 26.5 ± 2.5 | 890 ± 231 | 30 ± 7 | 15 ± 24 | 8.0 ± 0.7 | 111 ± 37 | 4 ± 3 | 12 ± 4 | 14 ± 22 | 74 ± 8 | |
| P(Depth) | <0.001 | <0.001 | 0.039 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| P(Soil province) | <0.001 | <0.001 | 0.045 | 0.001 | <0.001 | <0.001 | <0.001 | 0.034 | <0.001 | <0.001 | 0.005 | |
| P(Harvesting treatment) | 0.036 | 0.438 | 0.789 | 0.683 | 0.237 | 0.014 | 0.018 | 0.012 | 0.025 | 0.306 | 0.015 | |
| P(Depth × Soil prov.) | <0.001 | 0.103 | 0.001 | 0.636 | <0.001 | 0.001 | <0.001 | 0.455 | <0.001 | <0.001 | <0.001 | |
| P(Soil prov. × Harvesting) | 0.763 | 0.035 | 0.885 | 0.818 | 0.437 | 0.531 | 0.360 | 0.892 | 0.307 | 0.626 | 0.249 | |
| P(Depth × Harvesting) | 0.200 | 0.313 | 0.292 | 0.465 | 0.195 | 0.694 | 0.034 | 0.956 | 0.025 | 0.449 | 0.003 | |
| P(Depth × Soil prov. × Harvesting) | 0.141 | 0.009 | 0.352 | 0.626 | 0.563 | 0.430 | <0.001 | 0.665 | <0.001 | 0.138 | 0.102 | |
| Soil Province * | Harvesting Treatment | Thickness | P | Ca | Mg | K | Mn | Al | Fe |
|---|---|---|---|---|---|---|---|---|---|
| (cm) | (Total, kg·ha−1) | ||||||||
| B | SOH | 6.1 ± 0.6 | 88 ± 4 | 352 ± 41 | 57 ± 5 | 140 ± 7 | 54 ± 4 | 166 ± 16 | 110 ± 13 |
| WTH | 5.5 ± 0.7 | 77 ± 6 | 327 ± 48 | 60 ± 6 | 144 ± 9 | 40 ± 5 | 152 ± 20 | 111 ± 17 | |
| C | SOH | 9.9 ± 1.4 | 99 ± 6 | 470 ± 42 | 76 ± 7 | 131 ± 9 | 21 ± 3 | 283 ± 23 | 275 ± 23 |
| WTH | 14.7 ± 1.8 | 85 ± 8 | 341 ± 41 | 80 ± 9 | 139 ± 11 | 20 ± 3 | 225 ± 31 | 247 ± 33 | |
| D | SOH | 17.2 ± 1.9 | 97 ± 5 | 568 ± 81 | 68 ± 7 | 171 ± 9 | 25 ± 3 | 182 ± 14 | 125 ± 10 |
| WTH | 17.1 ± 2.1 | 116 ± 6 | 491 ± 37 | 99 ± 8 | 195 ± 11 | 15 ± 3 | 171 ± 18 | 108 ± 13 | |
| E | SOH | 13.6 ± 1.7 | 109 ± 8 | 461 ± 26 | 74 ± 5 | 190 ± 11 | 12 ± 2 | 298 ± 24 | 175 ± 15 |
| WTH | 11.9 ± 1.3 | 99 ± 7 | 554 ± 59 | 82 ± 5 | 160 ± 10 | 14 ± 2 | 292 ± 23 | 237 ± 16 | |
| P(Soil province) | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| P(Harvesting treatment) | 0.310 | 0.401 | 0.872 | 0.642 | 0.679 | 0.024 | 0.919 | 0.952 | |
| P(Soil prov. × Harvesting) | 0.093 | 0.008 | 0.304 | 0.143 | 0.026 | 0.026 | 0.682 | 0.012 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouimet, R.; Duchesne, L.; Tremblay, S. Long-Term Soil Fertility and Site Productivity in Stem-Only and Whole-Tree Harvested Stands in Boreal Forest of Quebec (Canada). Forests 2021, 12, 583. https://doi.org/10.3390/f12050583
Ouimet R, Duchesne L, Tremblay S. Long-Term Soil Fertility and Site Productivity in Stem-Only and Whole-Tree Harvested Stands in Boreal Forest of Quebec (Canada). Forests. 2021; 12(5):583. https://doi.org/10.3390/f12050583
Chicago/Turabian StyleOuimet, Rock, Louis Duchesne, and Stéphane Tremblay. 2021. "Long-Term Soil Fertility and Site Productivity in Stem-Only and Whole-Tree Harvested Stands in Boreal Forest of Quebec (Canada)" Forests 12, no. 5: 583. https://doi.org/10.3390/f12050583
APA StyleOuimet, R., Duchesne, L., & Tremblay, S. (2021). Long-Term Soil Fertility and Site Productivity in Stem-Only and Whole-Tree Harvested Stands in Boreal Forest of Quebec (Canada). Forests, 12(5), 583. https://doi.org/10.3390/f12050583