Nitrogen Addition Exacerbates the Negative Effect of Throughfall Reduction on Soil Respiration in a Bamboo Forest

 , ,
, ,
Abstract
:
1. Introduction
2. Material and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Soil Respiration Measurements
2.4. Soil Sampling and Analysis
2.5. Statistical Analyses
3. Results
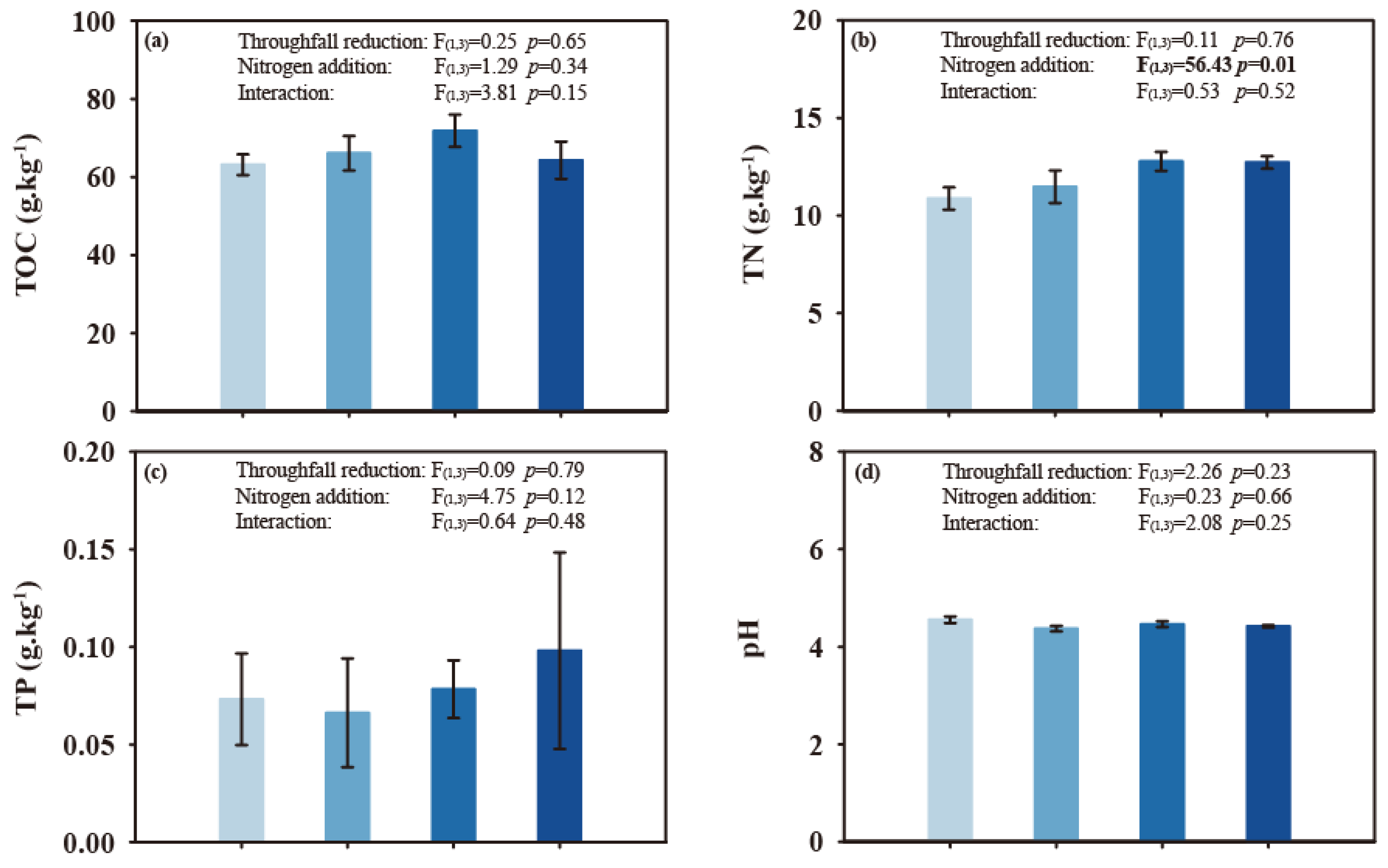
3.1. Throughfall Reduction and N Addition on Soil Environment, Fine Root Biomass and Microbial Properties
3.2. Effects of Throughfall Reduction and N Addition on Soil Respiration and Its Components
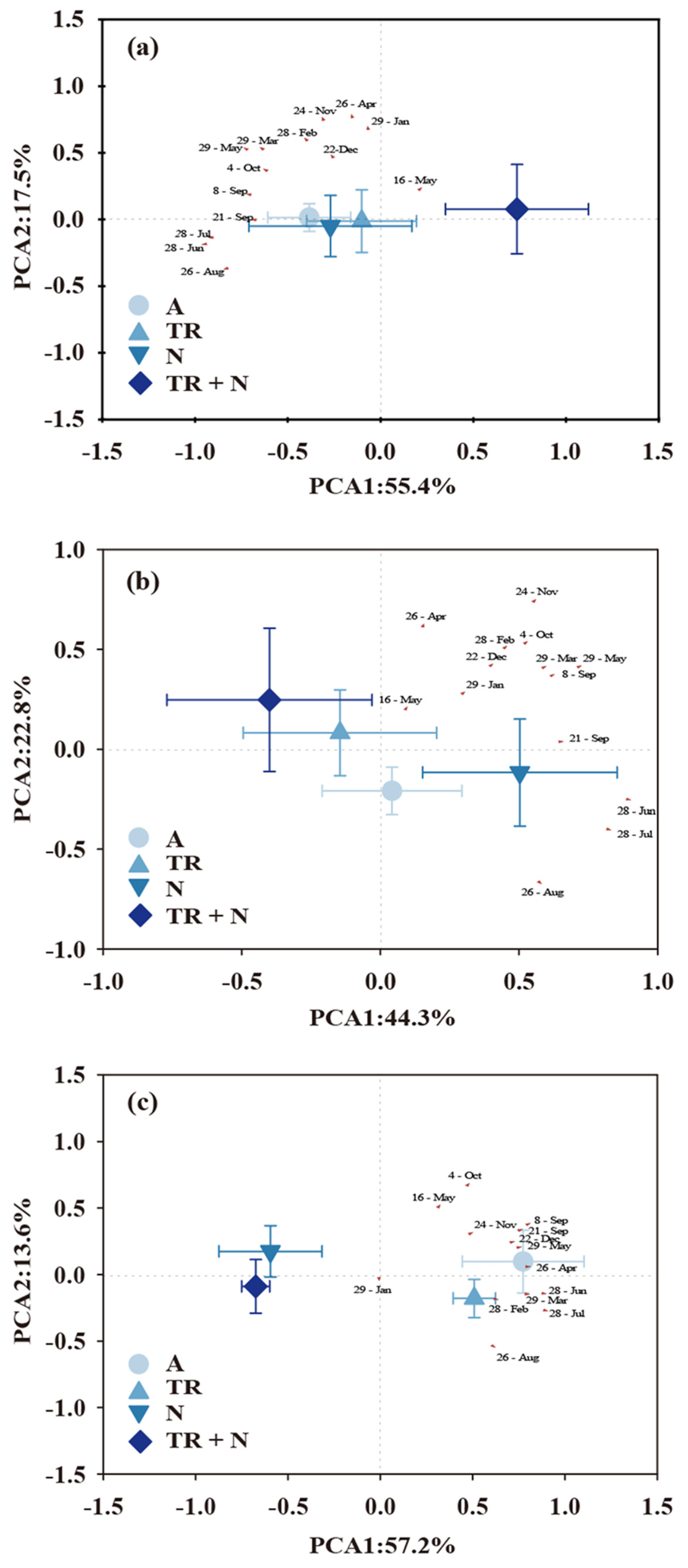
3.3. Controls for Soil Respiration and Its Components
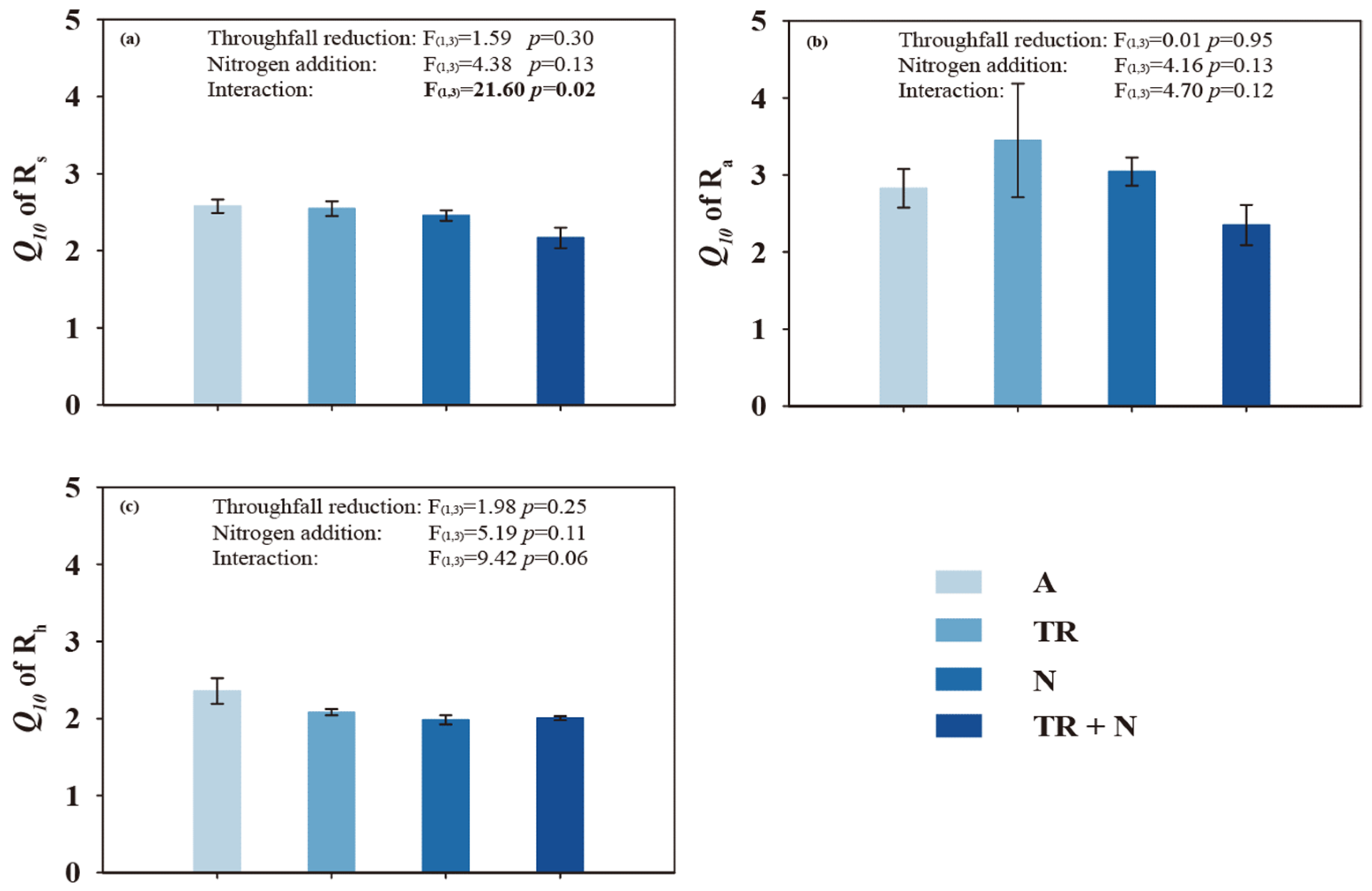
3.4. Throughfall Reduction and N Addition on Temperature Sensitivity of Soil Respiration and Its Components
4. Discussion
4.1. Effects of Throughfall Reduction on Soil Respiration and Its Components
4.2. Effects of N Addition on Soil Respiration and Its Components
4.3. Interactive Effects of N Addition and Throughfall Reduction on Soil Respiration and Its Components
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bond-Lamberty, B.; Thomson, A. Temperature-associated increases in the global soil respiration record. Nature 2010, 464, 579–582. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Zhou, L.; Liu, Y.; Zhang, Y.; Sha, L.; Song, Q.; Zhou, W.; Balasubramanian, D.; Palingamoorthy, G.; Gao, J.; Lin, Y.; et al. Soil respiration after six years of continuous drought stress in the tropical rainforest in Southwest China. Soil Biol. Biochem. 2019, 138, 107564. [Google Scholar] [CrossRef]
- Zhou, X.; Zhou, L.; Nie, Y.; Fu, Y.; Du, Z.; Shao, J.; Zheng, Z.; Wang, X. Similar responses of soil carbon storage to drought and irrigation in terrestrial ecosystems but with contrasting mechanisms: A meta-analysis. Agric. Ecosyst. Environ. 2016, 228, 70–81. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D.G. Human alteration of the global nitrogen cycle sources and consequences. Ecol. Appl. 1997, 7, 737–750. [Google Scholar] [CrossRef] [Green Version]
- Galloway, J.N. The global nitrogen cycle: Past, present and future. Sci. China Ser. C Life Sci. 2005, 48, 669–677. [Google Scholar]
- Liu, X.; Zhang, Y.; Han, W.; Tang, A.; Shen, J.; Cui, Z.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef]
- Song, X.; Peng, C.; Ciais, P.; Li, Q.; Xiang, W.; Xiao, W.; Zhou, G.; Deng, L. Nitrogen addition increased CO2 uptake more than non-CO2 greenhouse gases emissions in a Moso bamboo forest. Sci. Adv. 2020, 6, eaaw5790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, J.; Rillig, M. Distinguishing variability from uncertainty. Nat. Clim. Chang. 2014, 4, 153. [Google Scholar] [CrossRef]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Rillig, M.C.; Ryo, M.; Lehmann, A.; Aguilar-Trigueros, C.A.; Buchert, S.; Wulf, A.; Iwasaki, A.; Roy, J.; Yang, G. The role of multiple global change factors in driving soil functions and microbial biodiversity. Science 2019, 366, 886–890. [Google Scholar] [CrossRef]
- Felzer, B.S.; Cronin, T.W.; Melillo, J.M.; Kicklighter, D.W.; Schlosser, C.A.; Dangal, S.R.S. Nitrogen effect on carbon-water coupling in forests, grasslands, and shrublands in the arid western United States. J. Geophys. Res. 2011, 116. [Google Scholar] [CrossRef] [Green Version]
- Kuzyakov, Y. Sources of CO2 efflux from soil and review of partitioning methods. Soil Biol. Biochem. 2006, 38, 425–448. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, S.; Wang, J.; Chang, S.X.; Luan, J.; Liu, Y.; Lu, H.; Liu, X. Microbe-mediated attenuation of soil respiration in response to soil warming in a temperate oak forest. Sci. Total Environ. 2020, 711, 134563. [Google Scholar] [CrossRef]
- Hinko-Najera, N.; Fest, B.; Livesley, S.J.; Arndt, S.K. Reduced throughfall decreases autotrophic respiration, but not heterotrophic respiration in a dry temperate broadleaved evergreen forest. Agric. For. Meteorol. 2015, 200, 66–77. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, S.; Wan, S.; Wang, J.; Luan, J.; Wang, H. Differential responses of soil respiration to soil warming and experimental throughfall reduction in a transitional oak forest in central China. Agric. For. Meteorol. 2016, 226–227, 186–198. [Google Scholar] [CrossRef] [Green Version]
- Luan, J.; Wu, J.; Liu, S.; Roulet, N.; Wang, M. Soil nitrogen determines greenhouse gas emissions from northern peatlands under concurrent warming and vegetation shifting. Commun. Biol. 2019, 2, 132. [Google Scholar] [CrossRef]
- Chen, F.; Yan, G.; Xing, Y.; Zhang, J.; Wang, Q.; Wang, H.; Huang, B.; Hong, Z.; Dai, G.; Zheng, X.; et al. Effects of N addition and precipitation reduction on soil respiration and its components in a temperate forest. Agric. For. Meteorol. 2019, 271, 336–345. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, S.; Wang, Y.; Tian, P.; Sun, T. Influences of N deposition on soil microbial respiration and its temperature sensitivity depend on N type in a temperate forest. Agric. For. Meteorol. 2018, 260–261, 240–246. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, W.; Sun, T.; Chen, L.; Pang, X.; Wang, Y.; Xiao, F. N and P fertilization reduced soil autotrophic and heterotrophic respiration in a young Cunninghamia lanceolata forest. Agric. For. Meteorol. 2017, 232, 66–73. [Google Scholar] [CrossRef]
- Wang, B.; Zha, T.S.; Jia, X.; Wu, B.; Zhang, Y.Q.; Qin, S.G. Soil moisture modifies the response of soil respiration to temperature in a desert shrub ecosystem. Biogeosciences 2014, 11, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Davidson, E.A.; Verchot, L.V.; Cattânio, J.H.; Ackerman, I.L.; Carvalho, J.E.M. Effects of soil water content on soil respiration in forests and cattle pastures of eastern Amazonia. Biogeochemistry 2000, 48, 53–69. [Google Scholar] [CrossRef]
- Zhou, L.; Zhou, X.; Zhang, B.; Lu, M.; Luo, Y.; Liu, L.; Li, B. Different responses of soil respiration and its components to nitrogen addition among biomes: A meta-analysis. Glob. Chang. Biol. 2014, 20, 2332–2343. [Google Scholar] [CrossRef]
- Yan, L.; Chen, S.; Huang, J.; Lin, G. Differential responses of auto- and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe. Glob. Chang. Biol. 2010, 16, 2345–2357. [Google Scholar] [CrossRef]
- Huang, S.; Ye, G.; Lin, J.; Chen, K.; Xu, X.; Ruan, H.; Tan, F.; Chen, H.Y.H. Autotrophic and heterotrophic soil respiration responds asymmetrically to drought in a subtropical forest in the Southeast China. Soil Biol. Biochem. 2018, 123, 242–249. [Google Scholar] [CrossRef]
- Song, X.; Zhou, G.; Jiang, H.; Yu, S.; Fu, J.; Li, W.; Wang, W.; Ma, Z.; Peng, C. Carbon sequestration by Chinese bamboo forests and their ecological benefits: Assessment of potential, problems, and future challenges. Environ. Rev. 2011, 19, 418–428. [Google Scholar] [CrossRef] [Green Version]
- Mi, N.; Yu, G.; Wen, X.; Sun, X.; Wang, S.; Zhang, L.; Song, X. Use of ecosystem flux data and a simulation model to examine seasonal drought effects on a subtropical coniferous forest. Asia Pac. J. Atmos. Sci. 2009, 45, 207–220. [Google Scholar]
- Dai, A. Drought under global warming: A review. Wiley Interdisciplinary Rev. Clim. Chang. 2011, 2, 45–65. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Jia, Y.; He, N.; Zhu, J.; Chen, Z.; Wang, Q.; Piao, S.; Liu, X.; He, H.; Guo, X.; et al. Stabilization of atmospheric nitrogen deposition in China over the past decade. Nat. Geosci. 2019, 12, 424–429. [Google Scholar] [CrossRef]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- Wu, X.; Liu, S.; Luan, J.; Wang, Y.; Cai, C. Responses of water use in Moso bamboo (Phyllostachys heterocycla) culms of different developmental stages to manipulative drought. For. Ecosyst. 2019, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Bossio, D.A.; Scow, K.M. Impacts of carbon and flooding on soil microbial communities: Phospholipid fatty acid profiles and substrate utilization patterns. Microb. Ecol. 1998, 35, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Phospholipid fatty acid composition, biomass, and activity of microbial communities from two soil types experimentally exposed to different heavy metals. Appl. Environ. Microbiol. 1993, 59, 3605–3617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saiya-Cork, K.R.; Sinsabaugha, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Sistla, S.A.; Schimel, J.P. Seasonal patterns of microbial extracellular enzyme activities in an arctic tundra soil: Identifying direct and indirect effects of long-term summer warming. Soil Biol. Biochem. 2013, 66, 119–129. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Rinkes, Z.L.; Sinsabaugh, R.L.; Weintraub, M.N. Dynamic relationships between microbial biomass, respiration, inorganic nutrients and enzyme activities: Informing enzyme-based decomposition models. Front. Microbiol. 2013, 4, 223. [Google Scholar] [CrossRef] [Green Version]
- Jing, X.; Chen, X.; Fang, J.; Ji, C.; Shen, H.; Zheng, C.; Zhu, B. Soil microbial carbon and nutrient constraints are driven more by climate and soil physicochemical properties than by nutrient addition in forest ecosystems. Soil Biol. Biochem. 2020, 141, 107657. [Google Scholar] [CrossRef]
- Hu, S.; Li, Y.; Chang, S.X.; Li, Y.; Yang, W.; Fu, W.; Liu, J.; Jiang, P.; Lin, Z. Soil autotrophic and heterotrophic respiration respond differently to land-use change and variations in environmental factors. Agric. For. Meteorol. 2018, 250–251, 290–298. [Google Scholar] [CrossRef]
- Zhou, L.; Zhou, X.; Shao, J.; Nie, Y.; He, Y.; Jiang, L.; Wu, Z.; Hosseini Bai, S. Interactive effects of global change factors on soil respiration and its components: A meta-analysis. Glob. Chang. Biol. 2016, 22, 3157–3169. [Google Scholar] [CrossRef]
- Liu, L.; Wang, X.; Lajeunesse, M.J.; Miao, G.; Piao, S.; Wan, S.; Wu, Y.; Wang, Z.; Yang, S.; Li, P.; et al. A cross-biome synthesis of soil respiration and its determinants under simulated precipitation changes. Glob. Chang. Biol. 2016, 22, 1394–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Li, J.; Jin, Y.; Zhang, Y.; Sha, L.; Grace, J.; Song, Q.; Zhou, W.; Chen, A.; Li, P.; et al. The influence of drought strength on soil respiration in a woody savanna ecosystem, southwest China. Plant Soil. 2018, 428, 321–333. [Google Scholar] [CrossRef]
- Sun, S.; Lei, H.; Chang, S.X. Drought differentially affects autotrophic and heterotrophic soil respiration rates and their temperature sensitivity. Biol. Fertil. Soils 2019, 55, 275–283. [Google Scholar] [CrossRef]
- Harper, C.W.; Blair, J.M.; Fay, P.A.; Knapp, A.K.; Carlisle, J.D. Increased rainfall variability and reduced rainfall amount decreases soil CO2 flux in a grassland ecosystem. Glob. Chang. Biol. 2005, 11, 322–334. [Google Scholar] [CrossRef]
- Paradiso, E.; Jevon, F.; Matthes, J. Fine root respiration is more strongly correlated with root traits than tree species identity. Ecosphere. 2019, 10, e02944. [Google Scholar] [CrossRef]
- Wei, S.; Tie, L.; Liao, J.; Liu, X.; Du, M.; Lan, S.; Li, X.; Li, C.; Zhan, H.; Huang, C. Nitrogen and phosphorus co-addition stimulates soil respiration in a subtropical evergreen broad-leaved forest. Plant Soil 2020, 450, 171–182. [Google Scholar] [CrossRef]
- Högberg, M.N.; Högberg, P. Extramatrical ectomycorrhizal mycelium contributes one-third of microbial biomass and produces, together with associated roots, half the dissolved organic carbon in a forest soil. New Phytol. 2002, 154, 791–795. [Google Scholar] [CrossRef]
- Karst, J.; Gaster, J.; Wiley, E.; Landhausser, S.M. Stress differentially causes roots of tree seedlings to exude carbon. Tree Physiol. 2017, 37, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Preece, C.; Farre-Armengol, G.; Llusia, J.; Penuelas, J. Thirsty tree roots exude more carbon. Tree Physiol. 2018, 38, 690–695. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.; de Vries, F.T. Plant root exudation under drought: Implications for ecosystem functioning. New Phytol. 2020, 225, 1899–1905. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Xu, Y.; He, Y.; Zhou, X.; Fan, J.; Yu, H.; Ding, W. Nitrogen fertilization stimulated soil heterotrophic but not autotrophic respiration in cropland soils: A greater role of organic over inorganic fertilizer. Soil Biol. Biochem. 2018, 116, 253–264. [Google Scholar] [CrossRef]
- Zeng, W.; Wang, W. Combination of nitrogen and phosphorus fertilization enhance ecosystem carbon sequestration in a nitrogen-limited temperate plantation of Northern China. For. Ecol. Manag. 2015, 341, 59–66. [Google Scholar] [CrossRef]
- Gao, X.; Liu, S.; Wang, Y.; Luan, J.; Cai, C.; Ren, L. Effects of throughfall recduction and nitrogen addition on stoichiometry of leaf and fine root in Phyllostachys edulis forests. Acta Ecol. Sin. 2020. [Google Scholar] [CrossRef]
- Chapin, F.S., III. The mineral nutrition of wild plants. Annu. Rev. Ecol. Syst. 1980, 11, 230–236. [Google Scholar] [CrossRef]
- Chen, Z.; Xu, Y.; Fan, J.; Yu, H.; Ding, W. Soil autotrophic and heterotrophic respiration in response to different N fertilization and environmental conditions from a cropland in Northeast China. Soil Biol. Biochem. 2017, 110, 103–115. [Google Scholar] [CrossRef]
- Bowden, R.D.; Davidson, E.; Savage, K.; Arabia, C.; Steudler, P. Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest. For. Ecol. Manag. 2004, 196, 43–56. [Google Scholar] [CrossRef]
- Cusack, D.F.; Silver, W.L.; Torn, M.S.; McDowell, W.H. Effects of nitrogen additions on above- and belowground carbon dynamics in two tropical forests. Biogeochemistry 2011, 104, 203–225. [Google Scholar] [CrossRef] [Green Version]
- Hyvönen, R.; Ågren, G.I.; Linder, S.; Persson, T.; Cotrufo, M.F.; Ekblad, A.; Freeman, M.; Grelle, A.; Janssens, I.A.; Jarvis, P.G.; et al. The likely impact of elevated [CO2], nitrogen deposition, increased temperature and management on carbon sequestration in temperate and boreal forest ecosystems: A literature review. New Phytol. 2007, 173, 463–480. [Google Scholar] [CrossRef]
- Zhang, R.; Wu, J.; Li, Q.; Hänninen, H.; Peng, C.; Yao, H.; Song, X.; Ying, Y. Nitrogen deposition enhances photosynthesis in Moso bamboo but increases susceptibility to other stress factors. Front. Plant Sci. 2017, 8, 1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Chen, S.; Jiang, H.; Lin, Y.; Zhang, J.; Song, X.; Zhou, G. Water-Use Characteristics and Physiological Response of Moso Bamboo to Flash Droughts. Int. J. Environ. Res. Public Health 2019, 16, 2174. [Google Scholar] [CrossRef] [Green Version]
- Raich, J.W.; Schlesinger, W.H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B Chem. Phys. Meteorol. 2017, 44, 81–99. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Yuan, H.; Kimberley, M.O.; Jiang, H.; Zhou, G.; Wang, H. Soil CO2 flux dynamics in the two main plantation forest types in subtropical China. Sci. Total Environ. 2013, 444, 363–368. [Google Scholar] [CrossRef]
- Mo, J.; Zhang, W.E.I.; Zhu, W.; Gundersen, P.E.R.; Fang, Y.; Li, D.; Wang, H.U.I. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Glob. Chang. Biol. 2008, 14, 403–412. [Google Scholar] [CrossRef]
- Jia, X.; Shao, M.a.; Wei, X. Soil CO2 efflux in response to the addition of water and fertilizer in temperate semiarid grassland in northern China. Plant Soil. 2013, 373, 125–141. [Google Scholar] [CrossRef]
- Zhang, C.; Niu, D.; Hall, S.J.; Wen, H.; Li, X.; Fu, H.; Wan, C.; Elser, J.J. Effects of simulated nitrogen deposition on soil respiration components and their temperature sensitivities in a semiarid grassland. Soil Biol. Biochem. 2014, 75, 113–123. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A.; Luo, Y. On the variability of respiration in terrestrial ecosystems: Moving beyond Q10. Glob. Chang. Biol. 2006, 12, 154–164. [Google Scholar] [CrossRef]
- Li, Q.; Song, X.; Chang, S.X.; Peng, C.; Xiao, W.; Zhang, J.; Xiang, W.; Li, Y.; Wang, W. Nitrogen depositions increase soil respiration and decrease temperature sensitivity in a Moso bamboo forest. Agric. For. Meteorol. 2019, 268, 48–54. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rs | Ra | Rh | |
|---|---|---|---|
| Time | F(13,168) = 73.1 p < 0.001 | F(13,168) = 29.7 p < 0.001 | F(13,168) = 78.4 p < 0.001 |
| Throughfall reduction | F(1,168) = 14.2 p < 0.001 | F(1,168) = 9.9 p = 0.002 | F(1,168) = 3.5 p = 0.063 |
| N addition | F(1,168) = 5.4 p = 0.021 | F(1,168) = 1.9 p = 0.174 | F(1,168) = 97.2 p < 0.001 |
| Throughfall reduction × N addition | F(1,168) = 4.5 p = 0.034 | F(1,168) = 5.5 p = 0.020 | F(1,168) = 0.2 p = 0.633 |
| Throughfall reduction × Time | F(13,168) = 1.6 p = 0.090 | F(13,168) = 2.1 p = 0.019 | F(13,168) = 0.9 p = 0.522 |
| N addition × Time | F(13,168) = 0.6 p = 0.858 | F(13,168) = 0.2 p = 0.999 | F(13,168) = 4.1 p < 0.001 |
| Throughfall reduction × N addition × Time | F(13,168) = 0.6 p = 0.867 | F(13,168) = 0.6 p = 0.845 | F(13,168) = 0.6 p = 0.881 |
| a | b | c | R2 | p | |
|---|---|---|---|---|---|
| Rs = a.ebT.SWCc | |||||
| A | 0.55 ± 0.08 | 0.09 ± 0.00 | 0.47 ± 0.23 | 0.89 | 0.000 |
| TR | 0.76 ± 0.18 | 0.09 ± 0.00 | 0.52 ± 0.23 | 0.87 | 0.000 |
| N | 0.58 ± 0.09 | 0.09 ± 0.00 | 0.43 ± 0.23 | 0.81 | 0.000 |
| TR + N | 0.73 ± 0.12 | 0.08 ± 0.00 | 0.39 ± 0.05 | 0.76 | 0.000 |
| Ra = a.ebT.SWCc | |||||
| A | 0.16 ± 0.03 | 0.10 ± 0.01 | 0.03 ± 0.32 | 0.78 | 0.000 |
| TR | 0.31 ± 0.14 | 0.12 ± 0.02 | 0.54 ± 0.38 | 0.69 | 0.000 |
| N | 0.23 ± 0.09 | 0.11 ± 0.01 | −0.08 ±0.76 | 0.67 | 0.000 |
| TR + N | 0.35 ± 0.17 | 0.08 ± 0.01 | 0.07 ± 0.33 | 0.51 | 0.000 |
| Rh = a.ebT.SWCc | |||||
| A | 0.45 ± 0.03 | 0.09 ± 0.01 | 1.13 ± 0.13 | 0.86 | 0.000 |
| TR | 0.55 ± 0.08 | 0.07 ± 0.00 | 0.78 ± 0.19 | 0.80 | 0.000 |
| N | 0.51 ± 0.15 | 0.07 ± 0.00 | 1.14 ± 0.44 | 0.78 | 0.000 |
| TR + N | 0.51 ± 0.14 | 0.07 ± 0.00 | 0.85 ± 0.35 | 0.75 | 0.000 |
| TOC | TN | TP | pH | NO3-N | NH4-N | Total PLFAs | FRB | Vector Degree | Vector Length | |
|---|---|---|---|---|---|---|---|---|---|---|
| Rs | −0.141 | 0.009 | −0.402 | 0.391 | −0.186 | 0.210 | 0.312 | 0.680 ** | 0.377 | −0.477 |
| Ra | −0.080 | 0.175 | −0.323 | 0.371 | −0.125 | 0.197 | 0.165 | 0.534 * | 0.262 | −0.236 |
| Rh | −0.188 | −0.392 | −0.314 | 0.166 | −0.203 | 0.098 | 0.443 | 0.554 * | 0.386 | −0.716 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Liu, S.; Luan, J.; Chen, C.; Cai, C.; Zhou, F.; Di, Y.; Gao, X. Nitrogen Addition Exacerbates the Negative Effect of Throughfall Reduction on Soil Respiration in a Bamboo Forest. Forests 2021, 12, 724. https://doi.org/10.3390/f12060724
Wang Y, Liu S, Luan J, Chen C, Cai C, Zhou F, Di Y, Gao X. Nitrogen Addition Exacerbates the Negative Effect of Throughfall Reduction on Soil Respiration in a Bamboo Forest. Forests. 2021; 12(6):724. https://doi.org/10.3390/f12060724
Chicago/Turabian StyleWang, Yi, Shirong Liu, Junwei Luan, Chen Chen, Chunju Cai, Fan Zhou, Yaping Di, and Xiaomin Gao. 2021. "Nitrogen Addition Exacerbates the Negative Effect of Throughfall Reduction on Soil Respiration in a Bamboo Forest" Forests 12, no. 6: 724. https://doi.org/10.3390/f12060724