
Composition and Activity of N2-Fixing Microorganisms in Mangrove Forest Soils

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
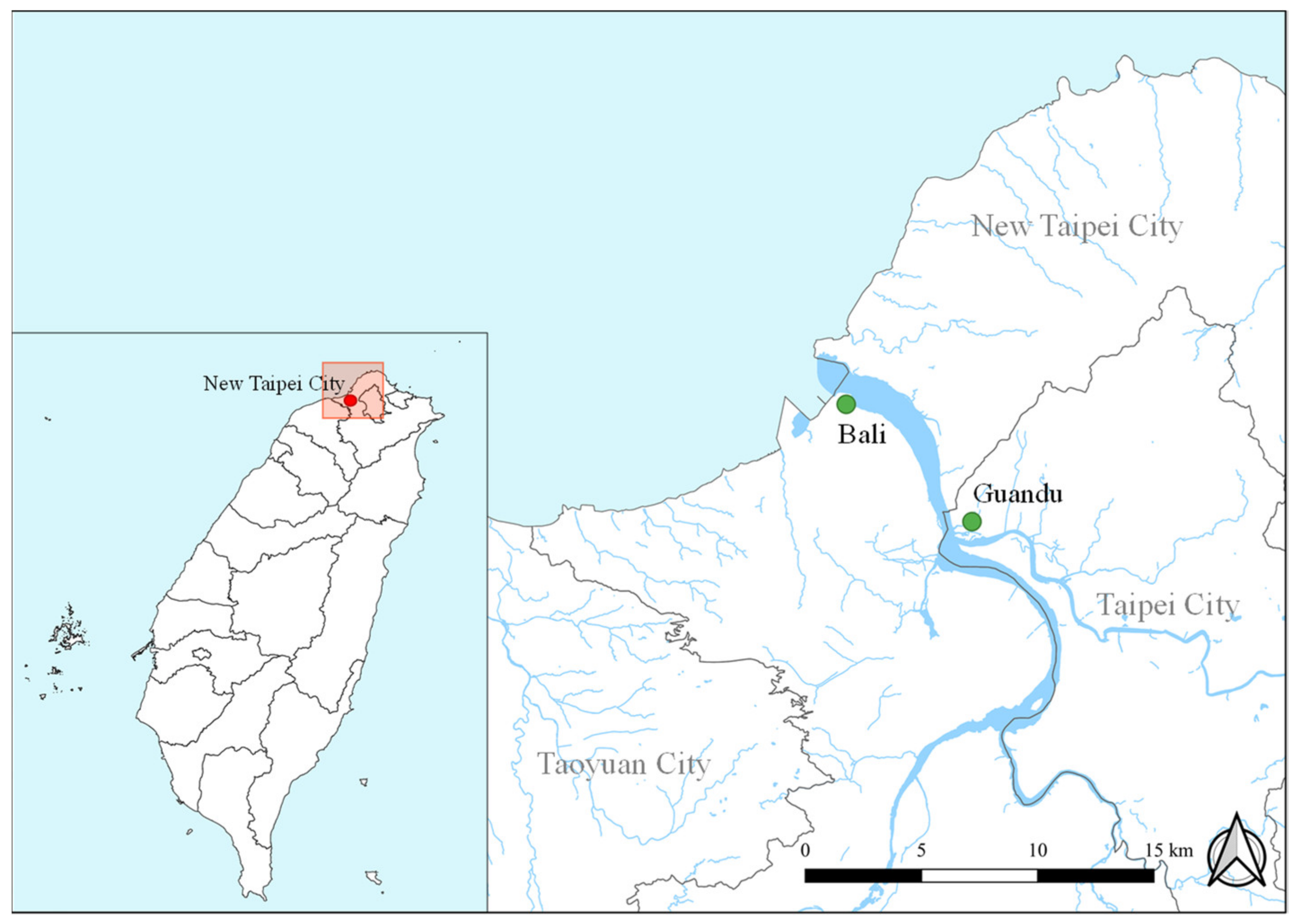
2.1. Sampling Locations
2.2. Determining Diazotroph Abundance and Compositions
2.3. Soil Incubation and Nitrogenase Activity Determination
2.4. Determining Soil Physiochemical Properties
2.5. Statistical Analysis
3. Results
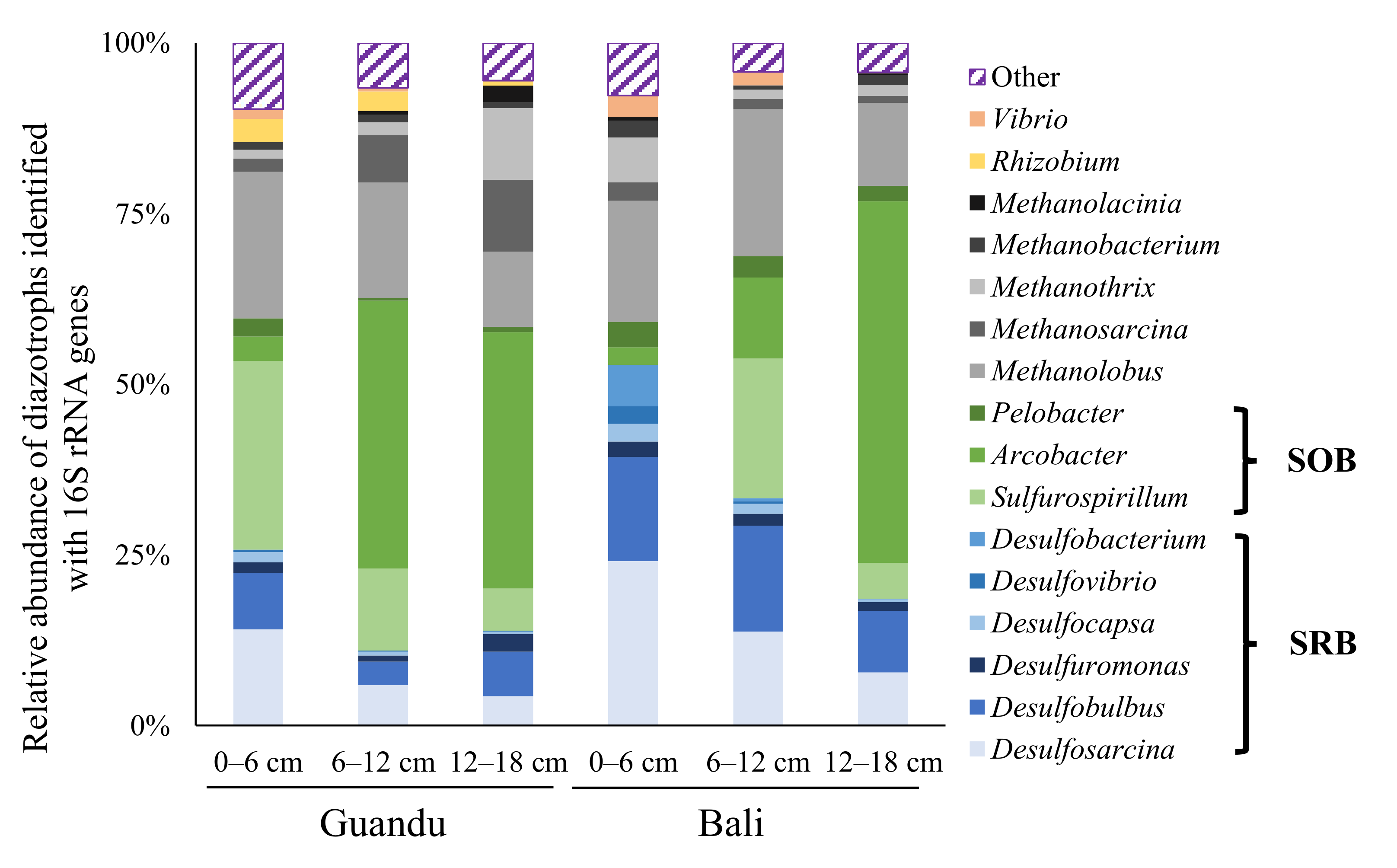
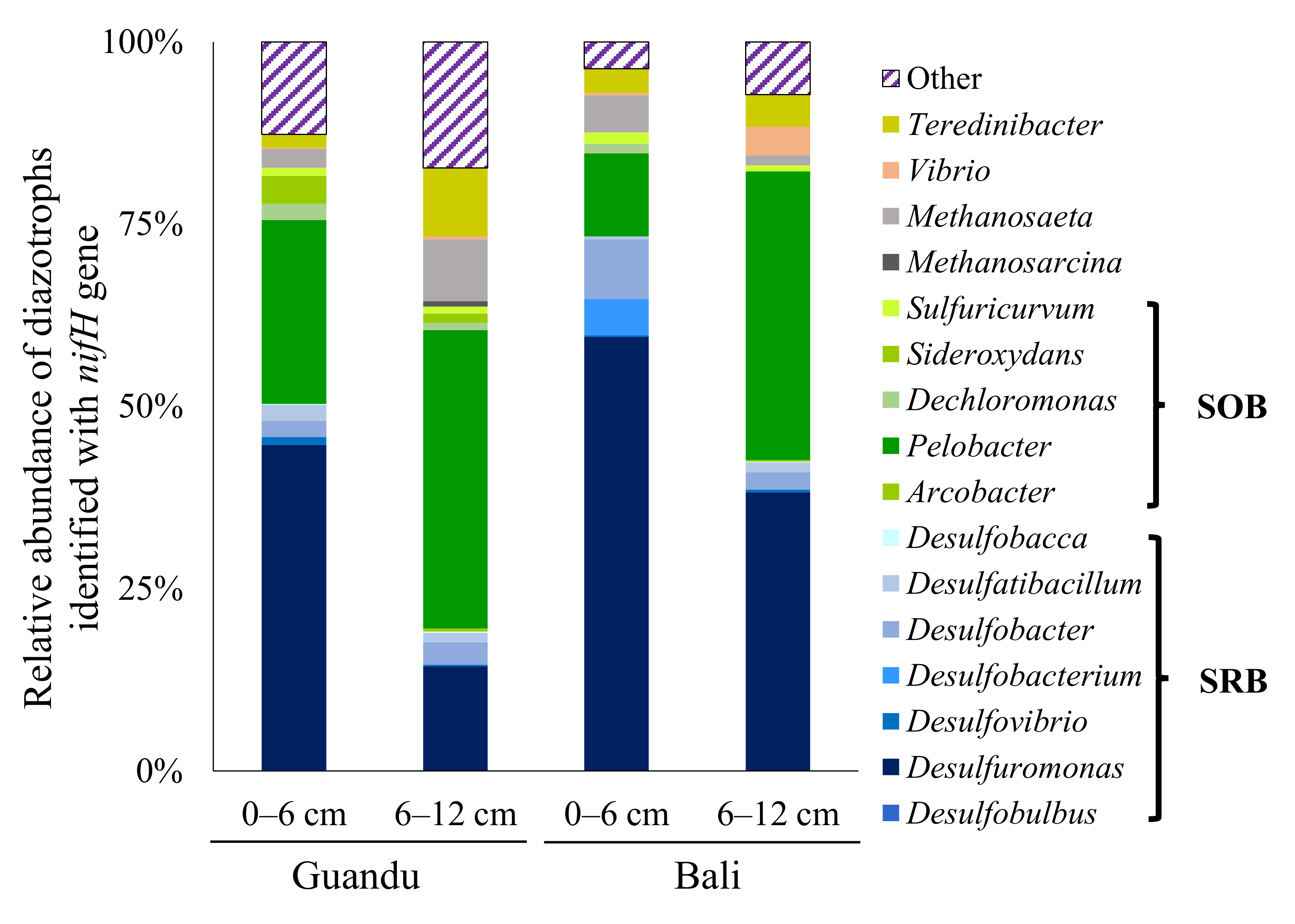
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feller, I.C. Effects of Nutrient Enrichment on Growth and Herbivory of Dwarf Red Mangrove (Rhizophora Mangle). Ecol. Monogr. 1995, 65, 477–505. [Google Scholar] [CrossRef]
- Feller, I.C.; McKee, K.L.; Whigham, D.F.; O’Neill, J.P. Nitrogen vs. phosphorus limitation across an ecotonal gradient in a mangrove forest. Biogeochemistry 2003, 62, 145–175. [Google Scholar] [CrossRef]
- Bertics, V.J.; Sohm, J.A.; Treude, T.; Chow, C.-E.T.; Capone, D.G.; Fuhrman, J.A.; Ziebis, W. Burrowing deeper into benthic nitrogen cycling: The impact of bioturbation on nitrogen fixation coupled to sulfate reduction. Mar. Ecol. Prog. Ser. 2010, 409, 1–15. [Google Scholar] [CrossRef]
- Hamilton, S.E.; Casey, D. Creation of a high spatio-temporal resolution global database of continuous mangrove forest cover for the 21st century (CGMFC-21). Glob. Ecol. Biogeogr. 2016, 25, 729–738. [Google Scholar] [CrossRef]
- Costanza, R.; de Groot, R.; Sutton, P.; van der Ploeg, S.; Anderson, S.J.; Kubiszewski, I.; Farber, S.; Turner, R.K. Changes in the global value of ecosystem services. Glob. Environ. Chang. 2014, 26, 152–158. [Google Scholar] [CrossRef]
- Holguin, G.; Vazquez, P.; Bashan, Y. The role of sediment microorganisms in the productivity, conservation, and rehabilitation of mangrove ecosystems: An overview. Biol. Fertil. Soils 2001, 33, 265–278. [Google Scholar] [CrossRef]
- Alongi, D.M.; Sasekumar, A.; Chong, V.C.; Pfitzner, J.; Trott, L.A.; Tirendi, F.; Dixon, P.; Brunskill, G.J. Sediment accumulation and organic material flux in a managed mangrove ecosystem: Estimates of land–ocean–atmosphere exchange in peninsular Malaysia. Mar. Geol. 2004, 208, 383–402. [Google Scholar] [CrossRef]
- Romero, I.C.; Jacobson, M.; Fuhrman, J.A.; Fogel, M.; Capone, D.G. Long-term nitrogen and phosphorus fertilization effects on N-2 fixation rates and nifH gene community patterns in mangrove sediments. Mar. Ecol. Evol. Perspect. 2012, 33, 117–127. [Google Scholar] [CrossRef]
- Shiau, Y.J.; Lin, M.F.; Tan, C.C.; Tian, G.; Chiu, C.Y. Assessing N2 fixation in estuarine mangrove soils. Estuar. Coast. Shelf Sci. 2017, 189, 84–89. [Google Scholar] [CrossRef]
- Tyler, A.C.; Mastronicola, T.A.; McGlathery, K.J. Nitrogen fixation and nitrogen limitation of primary production along a natural marsh chronosequence. Oecologia 2003, 136, 431–438. [Google Scholar] [CrossRef]
- Coyne, M. Soil Microbiology: An Exploratory Approach; Delmar Publishers: New York, NY, USA, 1999. [Google Scholar]
- Welsh, D.T.; Bourgues, S.; de Wit, R.; Herbert, R.A. Seasonal variations in nitrogen-fixation (acetylene reduction) and sulphate-reduction rates in the rhizosphere of Zostera noltii: Nitrogen fixation by sulphate reducing bacteria. Mar. Biol. 1996, 125, 619–628. [Google Scholar] [CrossRef]
- Diáková, K.; Biasi, C.; Čapek, P.; Martikainen, P.J.; Marushchak, M.E.; Patova, E.N.; Šantrůčková, H. Variation in N2 Fixation in Subarctic Tundra in Relation to Landscape Position and Nitrogen Pools and Fluxes. Arct. Antarct. Alp. Res. 2016, 48, 111–125. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Xiong, X.; Xu, Z.; Lu, C.; Cheng, H.; Lyu, X.; Zhang, J.; He, W.; Deng, W.; Lyu, Y.; et al. Bacterial Communities in the Rhizospheres of Three Mangrove Tree Species from Beilun Estuary, China. PLoS ONE 2016, 11, e0164082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yang, Q.; Ling, J.; Van Nostrand, J.D.; Shi, Z.; Zhou, J.; Dong, J. Diversity and Structure of Diazotrophic Communities in Mangrove Rhizosphere, Revealed by High-Throughput Sequencing. Front. Microbiol. 2017, 8, 2032. [Google Scholar] [CrossRef] [PubMed]
- Gomes, N.C.M.; Cleary, D.F.R.; Pinto, F.N.; Egas, C.; Almeida, A.; Cunha, A.; Mendonca-Hagler, L.C.S.; Smalla, K. Taking root: Enduring effect of rhizosphere bacterial colonization in mangroves. PLoS ONE 2010, 5, e14065. [Google Scholar] [CrossRef] [PubMed]
- Jing, H.; Xia, X.; Liu, H.; Zhou, Z.; Wu, C.; Nagarajan, S. Anthropogenic impact on diazotrophic diversity in the mangrove rhizosphere revealed by nifH pyrosequencing. Front. Microbiol. 2015, 6, 1172. [Google Scholar] [CrossRef] [Green Version]
- Flores-Mireles, A.; Winans, S.; Holguin, G. Molecular characterization of diazotrophic and denitrifying bacteria associated with mangrove roots. Appl. Environ. Microbiol. 2007, 73, 7308–7321. [Google Scholar] [CrossRef] [Green Version]
- Dias, A.C.F.; Pereira e Silva, M.d.C.; Cotta, S.R.; Dini-Andreote, F.; Soares, F.L.; Salles, J.F.; Azevedo, J.L.; van Elsas, J.D.; Andreote, F.D. Abundance and Genetic Diversity of nifH Gene Sequences in Anthropogenically Affected Brazilian Mangrove Sediments. Appl. Environ. Microbiol. 2012, 78, 7960. [Google Scholar] [CrossRef] [Green Version]
- Pelegrí, S.P.; Twilley, R.R. Heterotrophic nitrogen fixation (acetylene reduction) during leaf-litter decomposition of two mangrove species from South Florida, USA. Mar. Biol. 1998, 131, 53–61. [Google Scholar] [CrossRef]
- Kathiresan, K.; Bingham, B.L. Biology of mangroves and mangrove Ecosystems. In Advances in Marine Biology; Academic Press: Cambridge, MA, USA, 2001; Volume 40, pp. 81–251. [Google Scholar]
- Vepraskas, M.J.; Craft, C.B. Wetland Soils: Genesis, Hydrology, Landscapes, and Classification, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 1–506. [Google Scholar]
- Larsen, L.; Moseman, S.; Santoro, A.E.; Hopfensperger, K.; Burgin, A. A complex-systems approach to predicting effects of sea level rise and nitrogen loading on nitrogen cycling in coastal wetland ecosystems. In Ecological Dissertations in the Aquatic Sciences Symposium Proceeding VIII; The American Society of Limnology and Oceanography, Inc.: Waco, TX, USA, 2010; Chapter 5; pp. 67–92. [Google Scholar]
- Romero, I.; Jacobson-Meyers, M.; Fuhrman, J.; Capone, D. Phylogenetic Diversity of Diazotrophs along an Experimental Nutrient Gradient in Mangrove Sediments. J. Mar. Sci. Eng. 2015, 3, 699–719. [Google Scholar] [CrossRef]
- Muyzer, G.; Stams, A.J. The ecology and biotechnology of sulphate-reducing bacteria. Nat. Rev. Microbiol. 2008, 6, 441–454. [Google Scholar] [CrossRef] [PubMed]
- Mußmann, M.; Ishii, K.; Rabus, R.; Amann, R. Diversity and vertical distribution of cultured and uncultured Deltaproteobacteria in an intertidal mud flat of the Wadden Sea. Environ. Microbiol. 2005, 7, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Webster, G.; Watt, L.C.; Rinna, J.; Fry, J.C.; Evershed, R.P.; Parkes, R.J.; Weightman, A.J. A comparison of stable-isotope probing of DNA and phospholipid fatty acids to study prokaryotic functional diversity in sulfate-reducing marine sediment enrichment slurries. Environ. Microbiol. 2006, 8, 1575–1589. [Google Scholar] [CrossRef] [PubMed]
- Shiau, Y.-J.; Lin, C.-W.; Cai, Y.; Jia, Z.; Lin, Y.-T.; Chiu, C.-Y. Niche differentiation of active methane-oxidizing bacteria in estuarine mangrove forest soils in Taiwan. Microorganisms 2020, 8, 1248. [Google Scholar] [CrossRef]
- Poly, F.; Monrozier, L.J.; Bally, R. Improvement in the RFLP procedure for studying the diversity of nifH genes in communities of nitrogen fixers in soil. Res. Microbiol. 2001, 152, 95–103. [Google Scholar] [CrossRef]
- Simonet, P.; Grosjean, M.C.; Misra, A.K.; Nazaret, S.; Cournoyer, B.; Normand, P. Frankia genus-specific characterization by polymerase chain reaction. Appl. Environ. Microbiol. 1991, 57, 3278–3286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, R.W.F.; Holsten, R.D.; Jackson, E.K.; Burns, R.C. The acetylene-ethylene assay for N2 fixation: Laboratory and field evaluation. Plant Physiol. 1968, 43, 1185–1207. [Google Scholar] [CrossRef] [Green Version]
- Shiau, Y.J.; Chiu, C.Y. Changes in soil biochemical properties in a cedar plantation invaded by moso bamboo. Forests 2017, 8, 222. [Google Scholar] [CrossRef]
- Shiau, Y.J.; Wang, H.C.; Chen, T.H.; Jien, S.H.; Tian, G.L.; Chiu, C.Y. Improvement in the biochemical and chemical properties of badland soils by thorny bamboo. Sci. Rep. 2017, 7, 40561. [Google Scholar] [CrossRef] [Green Version]
- O’Dell, J.W. Determination of Nitrate-Nitrite Nitrogen by Automated Colorimetry 600/R-93-100; U.S. Environmental Protection Agency: Cincinnati, OH, USA, 1993.
- Huang, C.-Y.; Jien, S.-H.; Chen, T.-H.; Tian, G.; Chiu, C.-Y. Soluble organic C and N and their relationships with soil organic C and N and microbial characteristics in moso bamboo (Phyllostachys edulis) plantations along an elevation gradient in Central Taiwan. J. Soils Sediments 2014, 14, 1061–1070. [Google Scholar] [CrossRef]
- Sollins, P.; Glassman, C.; Paul, E.A.; Swanston, C.; Lajtha, K.; Heil, J.W.; Elliott, E.T. Soil carbon and nitrogen: Pools and fractions. In Standard Soil Methods for Long-Term Ecological Research; Robertson, G.P., Coleman, D.C., Bledsoe, C.S., Sollins, P., Eds.; Oxford University Press: New York, NY, USA, 1999; pp. 89–105. [Google Scholar]
- Dixon, R.; Kahn, D. Genetic regulation of biological nitrogen fixation. Nat. Rev. Microbiol. 2004, 2, 621–631. [Google Scholar] [CrossRef]
- Howarth, R.W.; Marino, R.; Cole, J.J. Nitrogen fixation in freshwater, estuarine, and marine ecosystems. 2. Biogeochemical controls1. Limnol. Oceanogr. 1988, 33, 688–701. [Google Scholar] [CrossRef]
- EPA. Environmental Water Quality Information. Available online: https://ewq.epa.gov.tw/Code/Station.aspx?Water=River*Sen&Area=11400011&Station=&IsCounty=N&Languages=en (accessed on 18 February 2021).
- Oremland, R.S.; Capone, D.G. Use of specific inhibitors in biogeochemistry and microbial ecology. In Advances in Microbial Ecology; Marshall, K.C., Ed.; Plenum Press: New York, NY, USA, 1988; Volume 10, pp. 285–383. [Google Scholar]
- Steppe, T.F.; Paerl, H.W. Potential N-2 fixation by sulfate-reducing bacteria in a marine intertidal microbial mat. Aquat. Microb. Ecol. 2002, 28, 1–12. [Google Scholar] [CrossRef]
- Stoeva, M.K.; Coates, J.D. Specific inhibitors of respiratory sulfate reduction: Towards a mechanistic understanding. Microbiology 2019, 165, 254–269. [Google Scholar] [CrossRef]
- Taylor, B.F.; Oremland, R.S. Depletion of adenosine triphosphate in Desulfovibrio by oxyanions of group VI elements. Curr. Microbiol. 1979, 3, 101–103. [Google Scholar] [CrossRef]
- Bertics, V.J.; Löscher, C.R.; Salonen, I.; Dale, A.W.; Gier, J.; Schmitz, R.A.; Treude, T. Occurrence of benthic microbial nitrogen fixation coupled to sulfate reduction in the seasonally hypoxic Eckernförde Bay, Baltic Sea. Biogeosciences 2013, 10, 1243–1258. [Google Scholar] [CrossRef] [Green Version]
- Fulweiler, R.W.; Nixon, S.W.; Buckley, B.A.; Granger, S.L. Net sediment N2 fluxes in a coastal marine system—Experimental manipulations and a conceptual model. Ecosystems 2008, 11, 1168–1180. [Google Scholar] [CrossRef]
- Nishiyama, M.; Sumikawa, Y.; Guan, G.; Marumoto, T. Relationship between microbial biomass and extractable organic carbon content in volcanic and non-volcanic ash soil. Appl. Soil Ecol. 2001, 17, 183–187. [Google Scholar] [CrossRef]
- Xu, J.M.; Cheng, H.H.; Koskinen, W.C.; Molina, J.A.E. Characterization of potentially bioreactive soil organic carbon and nitrogen by acid hydrolysis. Nutr. Cycl. Agroecosyst. 1997, 49, 267–271. [Google Scholar] [CrossRef]
- Shiau, Y.J.; Burchell, M.R.; Krauss, K.W.; Birgand, F.; Broome, S.W. Greenhouse gas emissions from a created brackish marsh in eastern North Carolina. Wetlands 2016, 36, 1009–1024. [Google Scholar] [CrossRef]
- Kessler, P.S.; Daniel, C.; Leigh, J.A. Ammonia switch-off of nitrogen fixation in the methanogenic archaeon Methanococcus maripaludis: Mechanistic features and requirement for the novel GlnB homologues, NifI(1) and NifI(2). J. Bacteriol. 2001, 183, 882–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
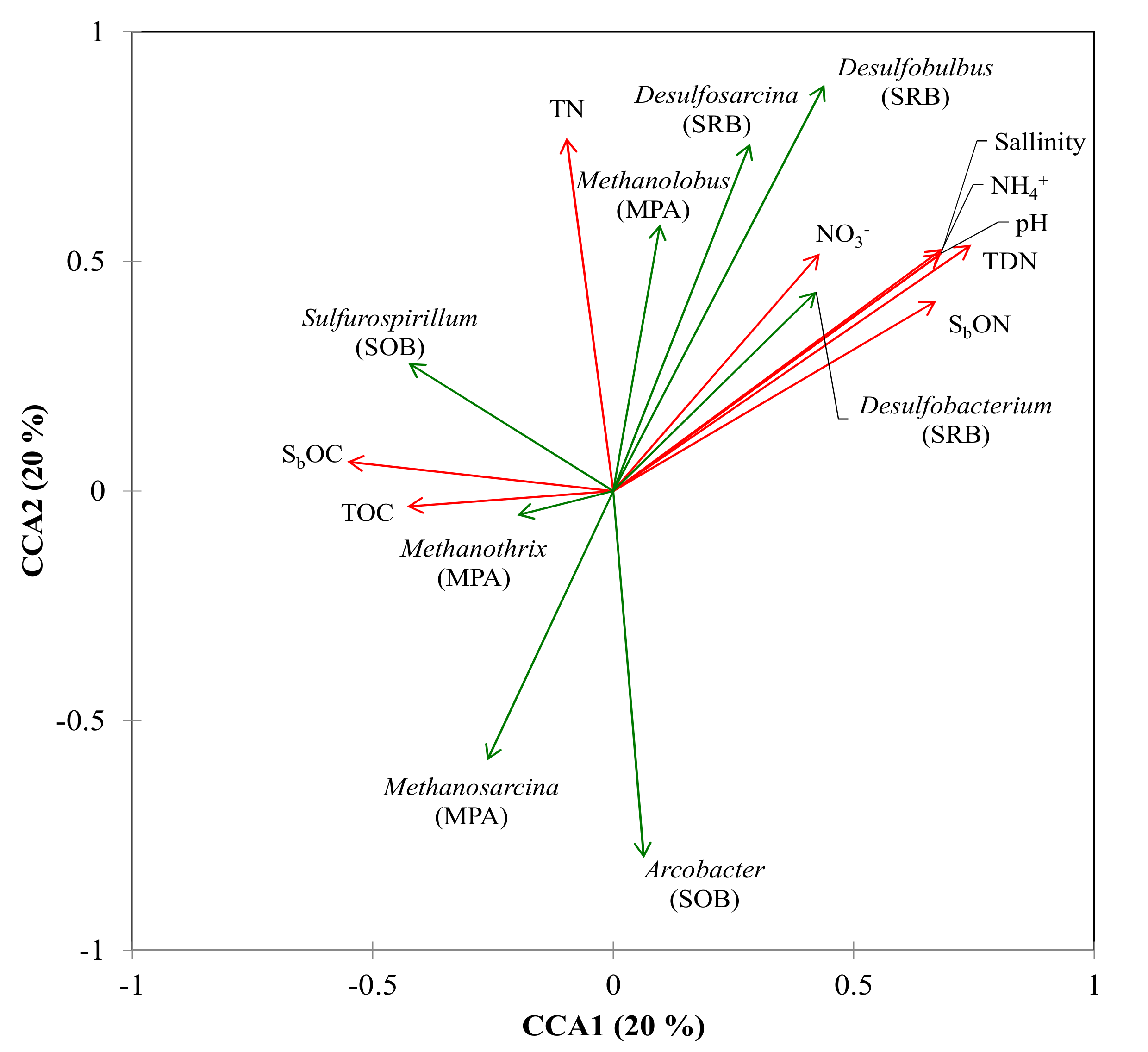{kind=link}
| Factor | Site Effect | Depth Effect | Interaction Effect (Site × Depth) (Soil That Is More Sensitive to Depth Change) |
|---|---|---|---|
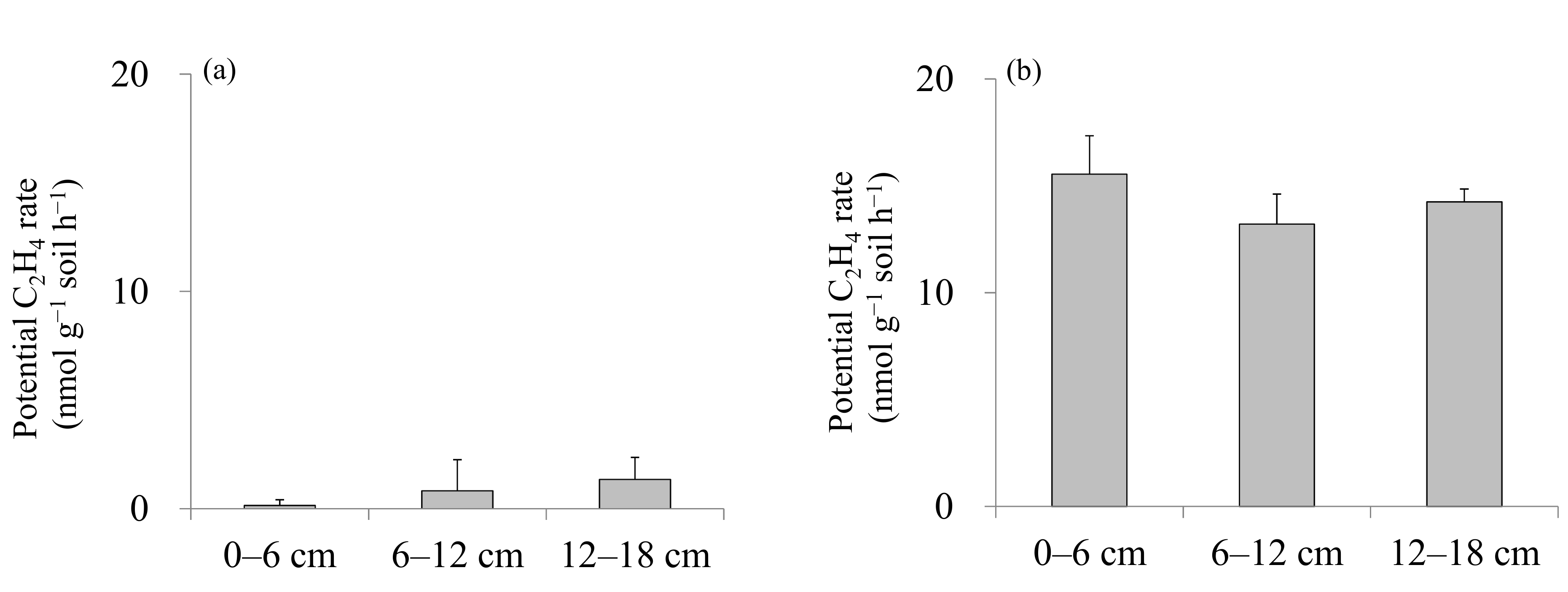
| Potential C2H4 rate | Bali > Guandu | n.d. * | n.d. |
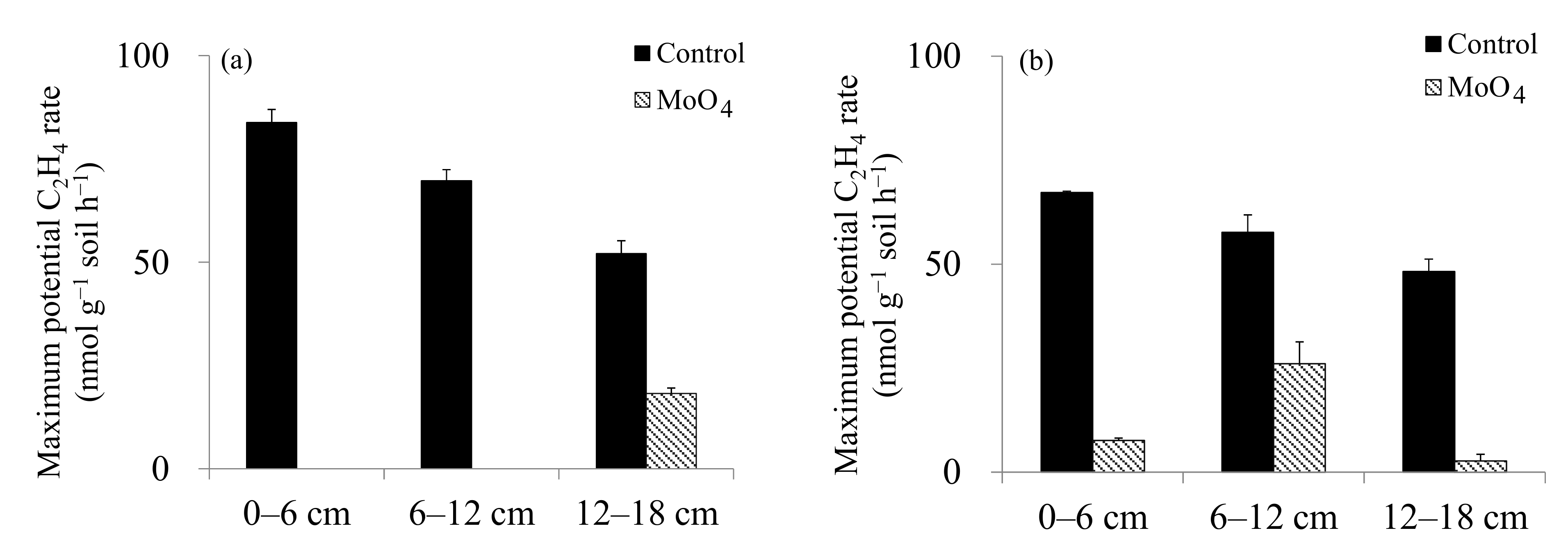
| Maximum potential C2H4 rate (amended with glucose) | Guandu > Bali | 0–6 > 6–12 >12–18 | Guandu |
| Potential C2H4 rate with amended MoO4 and glucose | Bali > Guandu | 6–12 = 12–18 > 0–6 | Guandu |
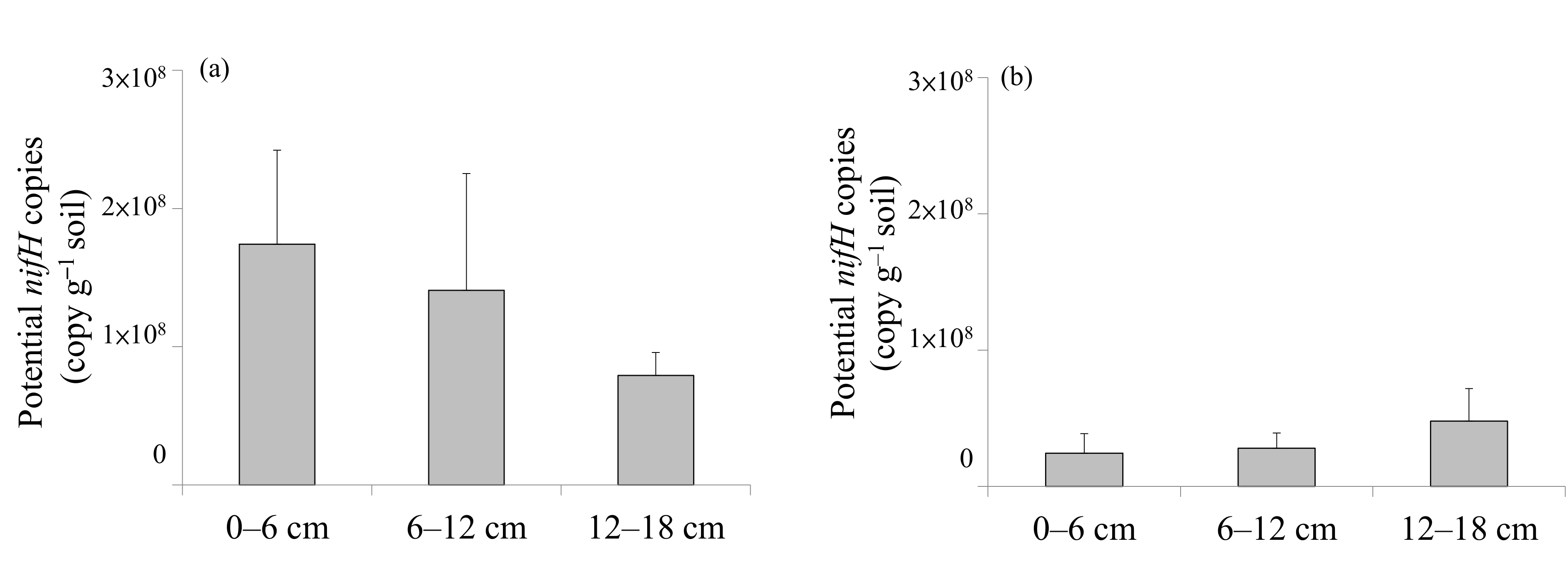
| Potential nifH copies | Guandu > Bali | n.d. | n.d. |
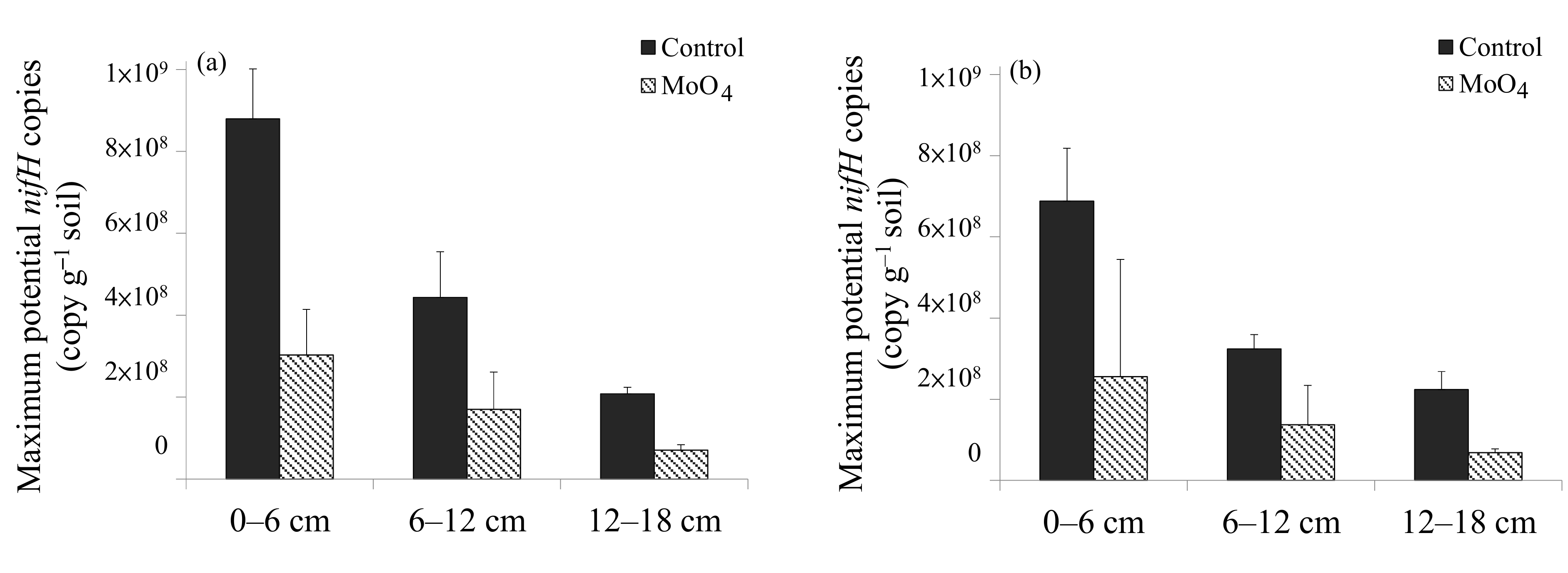
| Maximum potential nifH copies (amended with glucose) | Guandu > Bali | 0–6 > 6–12 >12–18 | Guandu |
| Potential nifH copies with amended MoO4 and glucose | Guandu > Bali | 0–6 > 6–12 = 12–18 | n.d. |
| Site | TOC (%) | TN (%) | SbOC * (mg C kg−1 Soil) | NH4+ | NO3− | SbON * | TDN * |
|---|---|---|---|---|---|---|---|
| (mg N kg−1 Soil) | |||||||
| Guandu | 3.40 ± 0.0 | 0.32 ± 0.0 | 12.7 ± 2.5 | 7.7 ± 0.2 | 0.3 ± 0.0 | 12.8 ± 4.8 | 20.7 ± 4.9 |
| Bali | 2.90 ± 0.0 | 0.28 ± 0.0 | 6.4 ± 3.8 | 54.2 ± 0.7 | 0.3 ± 0.0 | 27.7 ± 15.3 | 82.2 ± 15.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiau, Y.-J.; Lin, Y.-T.; Yam, R.S.W.; Chang, E.-H.; Wu, J.-M.; Hsu, T.-H.; Chiu, C.-Y. Composition and Activity of N2-Fixing Microorganisms in Mangrove Forest Soils. Forests 2021, 12, 822. https://doi.org/10.3390/f12070822
Shiau Y-J, Lin Y-T, Yam RSW, Chang E-H, Wu J-M, Hsu T-H, Chiu C-Y. Composition and Activity of N2-Fixing Microorganisms in Mangrove Forest Soils. Forests. 2021; 12(7):822. https://doi.org/10.3390/f12070822
Chicago/Turabian StyleShiau, Yo-Jin, Yu-Te Lin, Rita S. W. Yam, Ed-Haun Chang, Jhe-Ming Wu, Tsu-Hsuan Hsu, and Chih-Yu Chiu. 2021. "Composition and Activity of N2-Fixing Microorganisms in Mangrove Forest Soils" Forests 12, no. 7: 822. https://doi.org/10.3390/f12070822
APA StyleShiau, Y.-J., Lin, Y.-T., Yam, R. S. W., Chang, E.-H., Wu, J.-M., Hsu, T.-H., & Chiu, C.-Y. (2021). Composition and Activity of N2-Fixing Microorganisms in Mangrove Forest Soils. Forests, 12(7), 822. https://doi.org/10.3390/f12070822