
Competition Effects on Growth and Crown Dimensions of Shortleaf and Loblolly Pine in Mature, Natural-Origin, Pine–Hardwood Mixtures of the Upper West Gulf Coastal Plain of Arkansas, USA: A Neighborhood Analysis


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Design and Sampling
2.3. Analytical Approach
3. Results
3.1. Focal Tree Characteristics
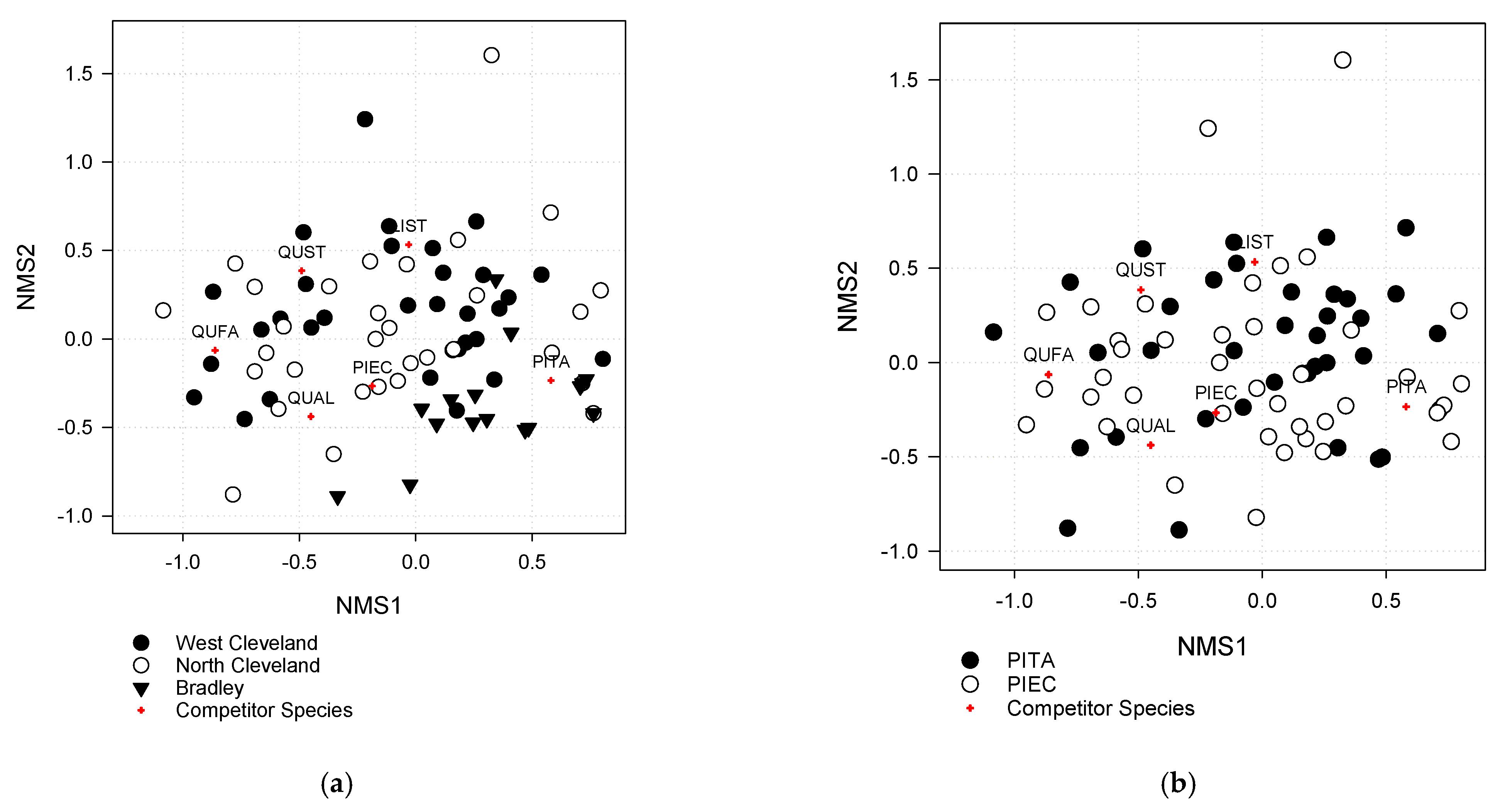
3.2. Neighborhood Composition and Structure
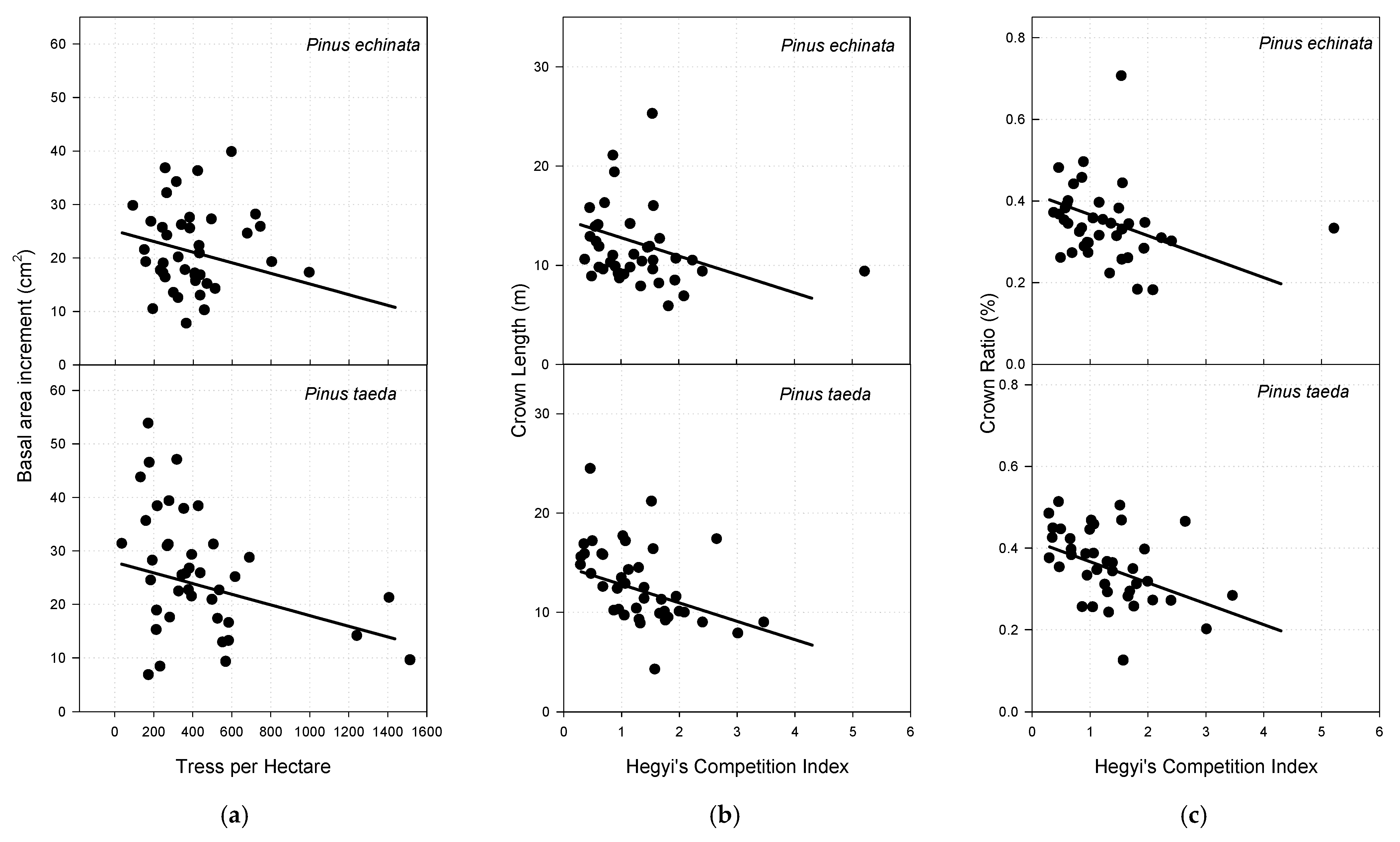
3.3. Basal Area Increment
3.4. Crown Length
3.5. Crown Ratio
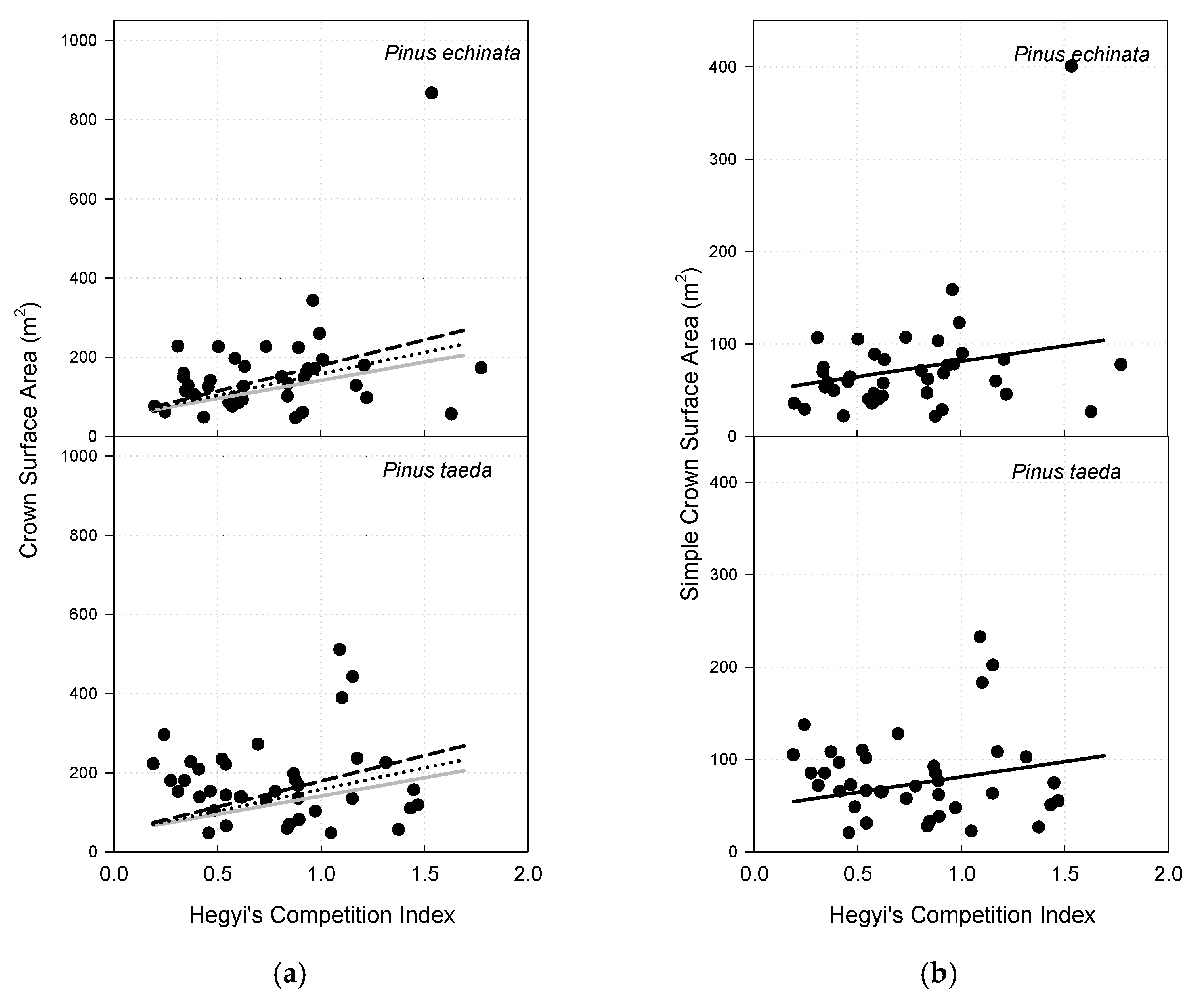
3.6. Crown Surface Area
3.7. Simple Crown Surface Area
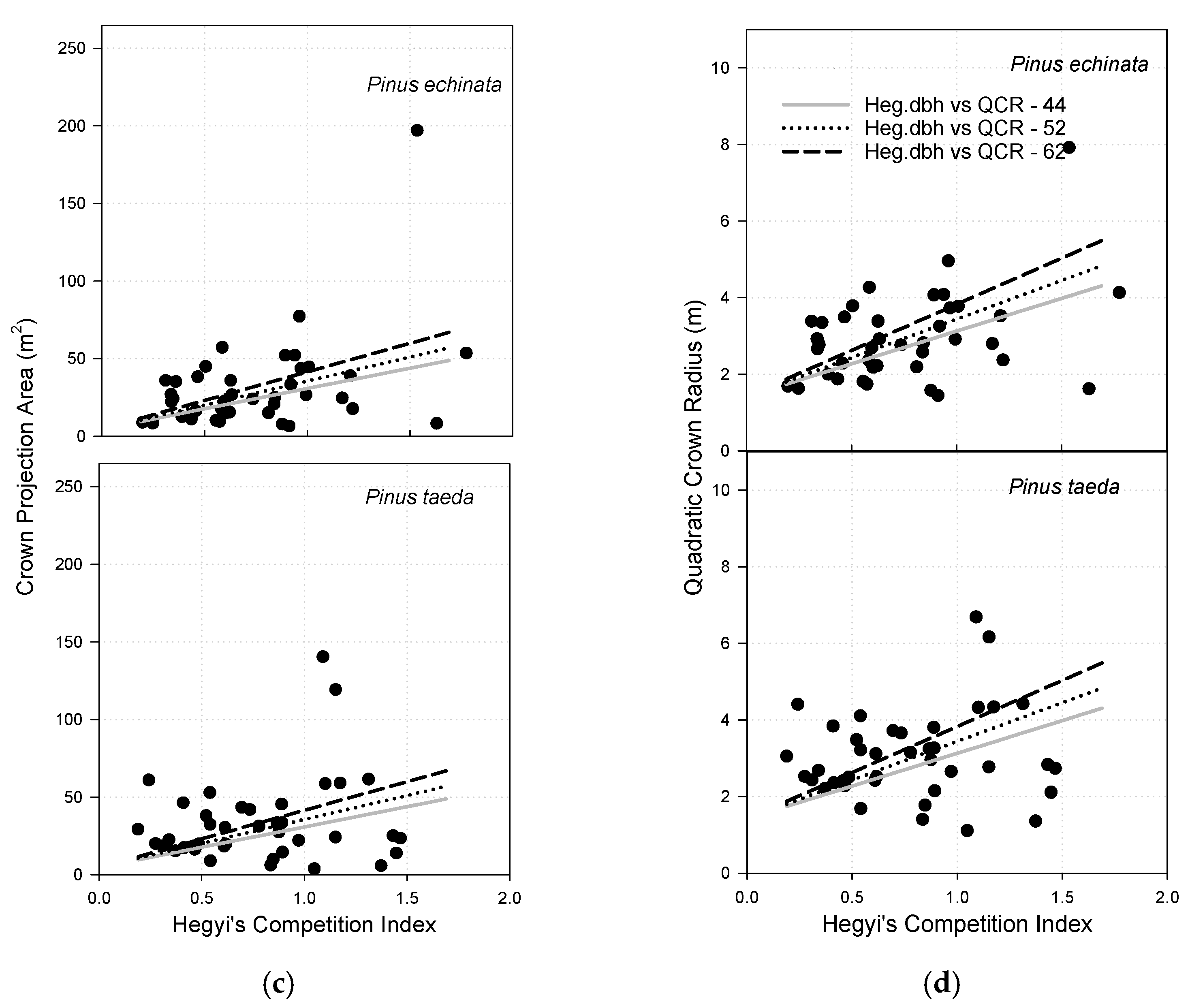
3.8. Crown Projection Area
3.9. Quadratic Crown Radius
4. Discussion
4.1. Focal Trees and Their Neighborhoods
4.2. Shortleaf Versus Loblolly
4.3. Competition Indices
4.4. Conspecific vs. Interspecific Neighborhoods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pacala, S.W.; Canham, C.D.; Saponara, J.; Silander, J.J.A.; Kobe, R.K.; Ribbens, E. Forest models defined by field measurements: Estimation, error analysis and dynamics. Ecol. Monogr. 1996, 66, 1–43. [Google Scholar] [CrossRef]
- Stoll, P.; Weiner, J. A Neighborhood View of interactions among individual plants. In The Geometry of Ecological Interactions: Simplifying Spatial Complexity; Cambridge University Press: Cambridge, UK, 2000; pp. 11–27. [Google Scholar] [CrossRef]
- Canham, C.D.; Uriarte, M. Analysis of neighborhood dynamics of forest ecosystems using likelihood methods and modeling. Ecol. Appl. 2006, 16, 62–73. [Google Scholar] [CrossRef]
- Fraver, S.; D’Amato, A.W.; Bradford, J.B.; Jonsson, B.G.; Jonsson, M.T.; Esseen, P.-A. Tree growth and competition in an old-growth Picea abies forest of boreal Sweden: Influence of tree spatial patterning. J. Veg. Sci. 2014, 25, 374–385. [Google Scholar] [CrossRef]
- Daniels, R.F.; Burkhart, H.E.; Clason, T.R. A comparison of competition measures for predicting growth of loblolly pine trees. Can. J. For. Res. 1986, 16, 1230–1237. [Google Scholar] [CrossRef]
- Miller, J.H.; Zutter, B.R.; Zedaker, S.M.; Edwards, M.B.; Haywood, J.D.; Newbold, R.A. A Regional Study on the Influence of Woody and Herbaceous Competition on Early Loblolly Pine Growth. South. J. Appl. For. 1991, 15, 169–179. [Google Scholar] [CrossRef]
- Cain, M.D. The Influence of Woody and Herbaceous Competition on Early Growth of Naturally Regenerated Loblolly and Shortleaf Pines. South. J. Appl. For. 1991, 15, 179–185. [Google Scholar] [CrossRef]
- Miller, J.H.; Zutter, B.R.; Zedaker, S.M.; Edwards, M.B.; Newbold, R.A. Growth and Yield Relative to Competition for Loblolly Pine Plantations to Midrotation—A Southeastern United States Regional Study. South. J. Appl. For. 2003, 27, 237–252. [Google Scholar] [CrossRef]
- Amateis, R.L.; Burkhart, H.E. Rotation-Age Results from a Loblolly Pine Spacing Trial. South. J. Appl. For. 2012, 36, 11–18. [Google Scholar] [CrossRef]
- Baldwin, V.; Peterson, K.D.; Clark, A.; Ferguson, R.B.; Strub, M.R.; Bower, D.R. The effects of spacing and thinning on stand and tree characteristics of 38-year-old Loblolly Pine. For. Ecol. Manag. 2000, 137, 91–102. [Google Scholar] [CrossRef]
- Lawson, E.R.; Kitchens, R.N. Shortleaf. In Silvicultural Systems for the Major Forest Types of the United States Agricultural Handbook 445; Burns, R.M., Ed.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1983; pp. 157–162. [Google Scholar]
- Wear, D.N.; Gries, J. The Southern Forest Futures Project: Technical Report; U.S. Department of Agriculture, Forest Service, Southern Research Station: Ashville, NC, USA, 2012.
- Guldin, J.M. Restoration and management of shortleaf pine in pure and mixed stands- science, empirical observation, and the wishful application of generalities. In Shortleaf Pine Restoration and Ecology in the Ozarks: Proceedings of a Symposium; Kabrick, J.M., Dey, D.C., Gwaze, D., Eds.; U.S. Department of Agriculture, Forest Service, North-ern Research Station: Springfield, MO, USA, 2007; pp. 47–58. [Google Scholar]
- Guldin, J.M. Ecology of shortleaf pine. In Proceedings of the Symposium on the Shortleaf Pine Ecosystem, Little Rock, AR, USA, 31 March–2 April 1986; Murphy, P.A., Ed.; Arkansas Cooperative Extension Service: Little Rock, AR, USA, 1986; pp. 25–40. [Google Scholar]
- Stewart, J.F.; Tauer, C.G.; Nelson, C.D. Bidirectional introgression between loblolly pine (Pinus taeda L.) and shortleaf pine (P. echinata Mill.) has increased since the 1950s. Tree Genet. Genom. 2011, 8, 725–735. [Google Scholar] [CrossRef]
- Cain, M.D.; Shelton, M.G. Survival and growth of Pinus echinata and Quercus seedlings in response to simulated summer and winter prescribed burns. Can. J. For. Res. 2000, 30, 1830–1836. [Google Scholar] [CrossRef]
- Yeiser, J.L.; Barnett, J.P. Growth and physiological response to four shortleaf pine families to herbicidal control of herbaceous competition. South. J. Appl. For. 1991, 15, 199–204. [Google Scholar] [CrossRef]
- Canham, C.D.; Papaik, M.J.; Uriarte, M.; McWilliams, W.H.; Jenkins, J.C.; Twery, M.J. Neighborhood Analyses Of Canopy Tree Competition Along Environmental Gradients In New England Forests. Ecol. Appl. 2006, 16, 540–554. [Google Scholar] [CrossRef]
- Papaik, M.J.; Canham, C.D. Multi-model analysis of tre competition along environmental gradients in southern New England forests. Ecol. Appl. 2006, 16, 1880–1892. [Google Scholar] [CrossRef]
- Coates, K.D.; Canham, C.D.; Lepage, P.T. Above- versus below-ground competitive effects and responses of a guild of temperate tree species. J. Ecol. 2009, 97, 118–130. [Google Scholar] [CrossRef]
- Hubbell, S.P. The Unified Neutral Theory of Biodiversity and Biogeography (MPB-32). In Monographs in Population Biology; Princeton University Press: Princeton, NJ, USA, 2001; Volume 32. [Google Scholar]
- Uriarte, M.; Condit, R.; Canham, C.; Hubbell, S.P. A spatially explicit model of sapling growth in a tropical forest: Does the identity of neighbours matter? J. Ecol. 2004, 92, 348–360. [Google Scholar] [CrossRef]
- Canham, C.; Lepage, P.T.; Coates, K.D. A neighborhood analysis of canopy tree competition: Effects of shading versus crowding. Can. J. For. Res. 2004, 34, 778–787. [Google Scholar] [CrossRef]
- Uriarte, M.; Canham, C.D.; Thompson, J.; Zimmermann, J.K. A neighborhood analysis of tree growth and survival in a hur-ricane-driven tropical forest. Ecol. Monogr. 2004, 74, 591–614. [Google Scholar] [CrossRef]
- Thorpe, H.; Astrup, R.; Trowbridge, A.; Coates, K. Competition and tree crowns: A neighborhood analysis of three boreal tree species. For. Ecol. Manag. 2010, 259, 1586–1596. [Google Scholar] [CrossRef]
- Waskiewicz, J.; Kenefic, L.; Weiskittel, A.; Seymour, R. Species mixture effects in northern red oak–eastern white pine stands in Maine, USA. For. Ecol. Manag. 2013, 298, 71–81. [Google Scholar] [CrossRef]
- Bragg, D.C. Reference Conditions for Old-Growth Pine Forests in the Upper West Gulf Coastal Plain. J. Torrey Bot. Soc. 2002, 129, 261. [Google Scholar] [CrossRef]
- Chen, J.; Tauer, C.G.; Bai, G.; Huang, Y.; Payton, M.; Holley, A.G. Bidirectional introgression between Pinus taeda and Pinus echinata: Evidence from morphological and molecular data. Can. J. For. Res. 2004, 34, 2508–2516. [Google Scholar] [CrossRef]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; McGraw-Hill, Inc.: New York, NY, USA, 1996; ISBN 0471138339. [Google Scholar]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Grissino-Mayer, H.D. Evaluating crossdating accuracy: A manual and tutorial for the computer program COFECHA. Tree-Ring Res. 2001, 57, 205–221. [Google Scholar]
- Bunn, A. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Miles, P.D.; Smith, B.W. Specific Gravity and Other Properties of Wood And Bark for 156 Tree Species Found in North America; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newton Square, PA, USA, 2009.
- Pinherio, J.; Bates, D. Mixed-Effects Models in S and S-Plus, 2000th ed.; Springer: New York, NY, USA, 2000; ISBN 0-387-98957-9. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Development Core Team: Vienna, Austria, 2013. [Google Scholar]
- Weiskittel, A.; Hann, D.W.; Kershaw, J.; Vanclay, J. Forest Growth and Yield Modeling; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2011. [Google Scholar]
- Hegyi, F. A simulation model for managing jack pine stands. RoyalColl. For. Res. Notes 1974, 30, 74–90. [Google Scholar]
- Brown, G.S. Point Density in Stems Per Acre. N. Z. For. Res. 1965, 38, 11. [Google Scholar]
- McCune, B.; Grace, J.B.; Urban, D.L. Analysis of Ecological Communities; MjM Software Design: Glenden Beach, OR, USA, 2002; ISBN 9780972129008. [Google Scholar]
- Hartsell, A.J.; Conner, R.C. Forest Area and Conditions: A 2010 Update of Chapter 16 of the Southern Forest Resource Assessment; U.S. Department of Agriculture, Forest Service, Southern Research Station: Ashville, NC, USA, 2013; Volume 174.
- Zhang, D.; Polyakov, M. The geographical distribution of plantation forests and land resources potentially available for pine plantations in the U.S. South. Biomass Bioenergy 2010, 34, 1643–1654. [Google Scholar] [CrossRef]
- Bragg, D.C.; Shelton, M.G. Lessons from 72 years of monitoring a once-cut pine-hardwood stand on the Crossett Experimental Forest, Arkansas, U.S.A. For. Ecol. Manag. 2011, 261, 911–922. [Google Scholar] [CrossRef]
- Bragg, D.C. Composition, Biomass, and Overstory Spatial Patterns in a Mature Pine-Hardwood Stand in Southeastern Arkansas. Castanea 2013, 78, 37–55. [Google Scholar] [CrossRef]
- Dipesh, K.; Will, R.; Lynch, T.B.; Heinemann, R.; Holeman, R. Comparison of Loblolly, Shortleaf, and Pitch X Loblolly Pine Plantations Growing in Oklahoma. For. Sci. 2015, 61, 540–547. [Google Scholar] [CrossRef]
- Schubert, M.R.; Rennie, J.C.; Schlarbaum, S.E. Four pine species grown at four spacings on the eastern highland rim, Tennessee, after 30 years. In Proceedings of the 12th Biennial Southern Silvicultural Research Conference, Biloxi, MS, USA, 24–28 February 2003; Connor, K.F., Ed.; U.S. Department of Agriculture, Forest Service, Southern Research Station: Ashville, NC, USA, 2004; pp. 433–436. [Google Scholar]
- Lynch, T.B.; Saud, P.; Dipesh, K.; Will, R. Plantation Site Index Comparisons for Shortleaf Pine and Loblolly Pine in Oklahoma, USA. For. Sci. 2016, 62, 546–552. [Google Scholar] [CrossRef]
- Schultz, R.P. Loblolly Pine: The Ecology and Culture of Loblolly Pine (Pinus taeda L.); U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1997.
- Cl, V.; Blazier, M.; McConnell, E.; Adams, J.P. Determining unthinned shortleaf pine (Pinus echinata Mill.) plantation economic rotation ages in the Western Gulf. For. Res. Eng. Int. J. 2018, 2, 132–138. [Google Scholar] [CrossRef]
- Alemdag, I. Evaluation of Some Competition Indexes for the Prediction of Diameter Increment in Planted White Spruce; Canadian Forestry Service, Department of the Environment: Ottawa, ON, Canada, 1978. [Google Scholar]
- Bella, I. A new competition model for individual trees. For. Sci. 1971, 17, 364–372. [Google Scholar]
- Pukkala, T.; Kolstrom, T. Competition indicies and the prediction of radial growth in Scots Pine. Silva Fenn. 1987, 21, 55–67. [Google Scholar] [CrossRef]
- Tome, M.; Burkhart, H.E. Distance-dependent competition measures for predicting growth of individual trees. For. Sci. 1989, 35, 816–831. [Google Scholar]
- Martin, G.L.; Ek, A.R. A comparison of competition measures and growth models for predicting plantation red pine diameter and height growth. For. Sci. 1984, 30, 731–743. [Google Scholar]
- Biging, G.S.; Dobbertin, M. Evaluation of competition indices in individual tree growth models. For. Sci. 1995, 41, 360–377. [Google Scholar]
- Contreras, M.A.; Affleck, D.; Chung, W. Evaluating tree competition indices as predictors of basal area increment in western Montana forests. For. Ecol. Manag. 2011, 262, 1939–1949. [Google Scholar] [CrossRef]
- Biging, G.S.; Dobbertin, M. A comparison of distance-dependent competition measures for height and basal area growth of individual conifer trees. For. Sci. 1992, 38, 695–720. [Google Scholar]
- Daniels, R.F. Simple competition indices and their correlation with annual loblolly pine tree growth. For. Sci. 1976, 22, 454–456. [Google Scholar]
- Larson, P.R. Stem Form Development of Forest Trees. For. Sci. 1963, 9, a0001-42. [Google Scholar] [CrossRef]
- Farrar, J.L. Longitudinal Variation in The Thickness Of The Annual Ring. For. Chron. 1961, 37, 323–349. [Google Scholar] [CrossRef][Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crown Dimension | Measurements and Calculation |
|---|---|
| Crown length (CL) | , where HT is total height and HLC 1 is the height to the base of the live crown |
| Live crown ratio (LCR) | |
| Quadratic mean crown radius (QCR) | , where r1 − r4 are the crown radii in the four cardinal directions |
| Crown projection area (CPA) | |
| Crown surface area (CSA) | |
| Simple crown surface area (CSAS) | , where QCW is quadratic crown width 2 |
| Stand | Species | n | DBH 1 (cm) | Height (m) | Age (Years) | Crown Ratio (%) | Crown Radius 2 (m) | NeaNeig 3 (m) |
|---|---|---|---|---|---|---|---|---|
| North Cleveland | Shortleaf | 16 | 49.5 (10.3) | 34.5 (3.1) | 107 (5) | 32 (8) | 2.6 (0.9) | 3.3 (1.3) |
| Loblolly | 16 | 56.2 (12.8) | 38.0 (2.1) | 101 (4) | 33 (1) | 2.6 (0.9) | 3.8 (1.2) | |
| West Cleveland | Shortleaf | 16 | 51.2 (7.9) | 33.3 (4.4) | 95 (7) | 36 (12) | 3.5 (1.4) | 2.8 (1.3) |
| Loblolly | 16 | 56.2 (13.5) | 33.4 (4) | 86 (4) | 37 (7) | 3.8 (1.2) | 3.1 (1.4) | |
| Bradley | Shortleaf | 8 | 52.7 (9.8) | 31.4 (3.6) | 131 (9) | 36 (5) | 2.6 (1.1) | 2.9 (0.8) |
| Loblolly | 8 | 56.4 (12.7) | 36.1 (5.7) | 118 (4) | 39 (9) | 2.5 (0.7) | 3.7 (1.8) |
| Stand | Species | n | CC Dens 1 (trees.ha−1) | CC BA 2 (m2.ha−1) | QMD 3 (cm) | Height (m) | Crown Ratio (%) |
|---|---|---|---|---|---|---|---|
| North Cleveland | Shortleaf | 16 | 436 (62) | 65.5 (15.4) | 41.3 (1.9) | 25.5 (2.4) | 46 (3) |
| Loblolly | 16 | 522 (114) | 83.8 (21.8) | 47.0 (3.1) | 28.4 (2.3) | 48 (4) | |
| West Cleveland | Shortleaf | 16 | 349 (39) | 35.7 (6.6) | 33.6 (2.2) | 24.9 (2.1) | 50 (4) |
| Loblolly | 16 | 384 (40) | 41.5 (6.4) | 37.1 (1.8) | 24.3 (2.1) | 49 (4) | |
| Bradley | Shortleaf | 8 | 396 (33) | 29.7 (5.2) | 30.3 (2.8) | 21.3 (2.7) | 49 (0.1) |
| Loblolly | 8 | 332 (57) | 51.1 (11.0) | 42.9 (5.0) | 27.1 (5.1) | 51 (0.1) |
| Predictor | Coefficient | SE | p |
|---|---|---|---|
| Basal area increment 1 (cm2) | |||
| Intercept | 25.081 | 3.390 | <0.001 |
| Loblolly pine (Pinus taeda) 2 | 2.806 | 1.667 | 0.0967 |
| Trees per hectare | −0.010 | 0.003 | <0.001 |
| Crown length (m) | |||
| Intercept | 5.923 | 1.838 | 0.002 |
| DBH (cm) | 0.163 | 0.032 | <0.001 |
| HegyiCL | −1.848 | 0.472 | <0.001 |
| Crown ratio (%) | |||
| Intercept | 0.238 | 0.045 | <0.001 |
| DBH (cm) | 0.003 | 0.0007 | <0.001 |
| HegyiCL | −0.051 | 0.013 | <0.001 |
| Crown surface area (m2) | |||
| Intercept | 109.06 | 13.543 | <0.001 |
| Trees per hectare | −0.145 | 0.022 | <0.001 |
| DBH (cm) * HegyiDBH | 2.092 | 0.380 | <0.001 |
| Simple crown surface area (m2) | |||
| Intercept | −41.87 | 15.424 | 0.008 |
| Trees per hectare | −0.042 | 0.005 | <0.001 |
| DBH (cm) | 2.00 | 0.299 | <0.001 |
| HegyiDBH | 33.17 | 7.364 | <0.001 |
| Crown projection area (m2) | |||
| Intercept | 15.521 | 2.280 | <0.001 |
| Trees per hectare | −0.026 | 0.002 | <0.001 |
| DBH (cm) * HegyiDBH | 0.594 | 0.090 | <0.001 |
| Quadratic crown radius (m) | |||
| Intercept | 2.342 | 0.150 | <0.001 |
| Trees per hectare | −0.002 | <0.001 | <0.001 |
| DBH (cm) * HegyiDBH | 0.039 | 0.004 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bataineh, M.; Childs, E. Competition Effects on Growth and Crown Dimensions of Shortleaf and Loblolly Pine in Mature, Natural-Origin, Pine–Hardwood Mixtures of the Upper West Gulf Coastal Plain of Arkansas, USA: A Neighborhood Analysis. Forests 2021, 12, 935. https://doi.org/10.3390/f12070935
Bataineh M, Childs E. Competition Effects on Growth and Crown Dimensions of Shortleaf and Loblolly Pine in Mature, Natural-Origin, Pine–Hardwood Mixtures of the Upper West Gulf Coastal Plain of Arkansas, USA: A Neighborhood Analysis. Forests. 2021; 12(7):935. https://doi.org/10.3390/f12070935
Chicago/Turabian StyleBataineh, Mohammad, and Ethan Childs. 2021. "Competition Effects on Growth and Crown Dimensions of Shortleaf and Loblolly Pine in Mature, Natural-Origin, Pine–Hardwood Mixtures of the Upper West Gulf Coastal Plain of Arkansas, USA: A Neighborhood Analysis" Forests 12, no. 7: 935. https://doi.org/10.3390/f12070935
APA StyleBataineh, M., & Childs, E. (2021). Competition Effects on Growth and Crown Dimensions of Shortleaf and Loblolly Pine in Mature, Natural-Origin, Pine–Hardwood Mixtures of the Upper West Gulf Coastal Plain of Arkansas, USA: A Neighborhood Analysis. Forests, 12(7), 935. https://doi.org/10.3390/f12070935