
Phenotypic Variability and Differences in the Drought Response of Norway Spruce Pendula and Pyramidalis Half-Sib Families

 ,
,  and
and
Abstract
1. Introduction
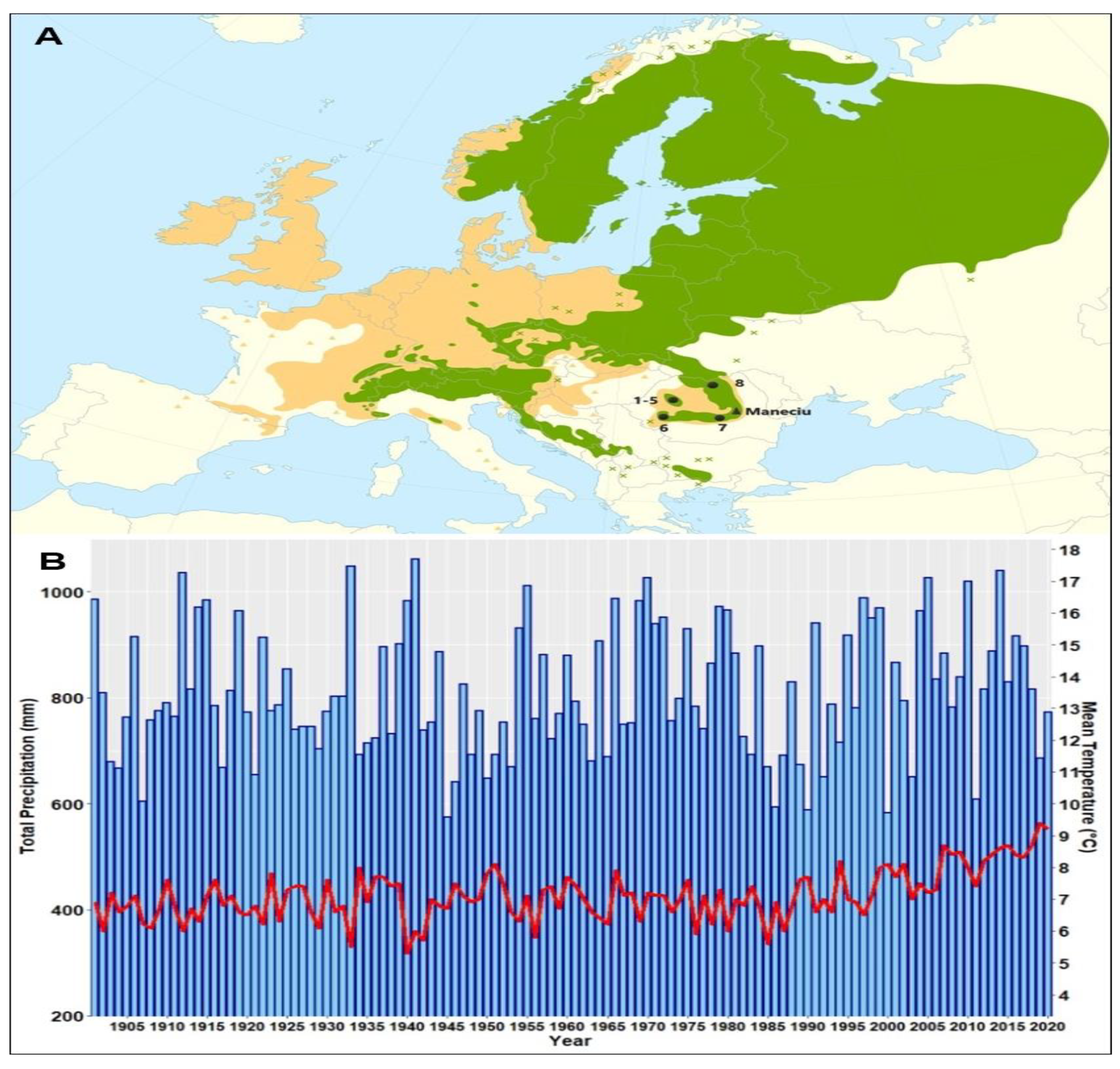
2. Materials and Methods
- -
- De Martonne aridity index: AI = P × (T + 10)−1, where P is the amount of the annual precipitation and T is the average annual temperature; the optimal for spruce is in the range of 40–60 [43].
- -
- -
- Standardized precipitation index (SPI): , for x > 0, where α is a shape parameter, β is a scale parameter, x is the precipitation amount, and Г(α) is the gamma function. The SPI index uses a probability density function of the gamma distribution to determine the wet or drought periods for a certain period of time based on the precipitation records [45,46,47].
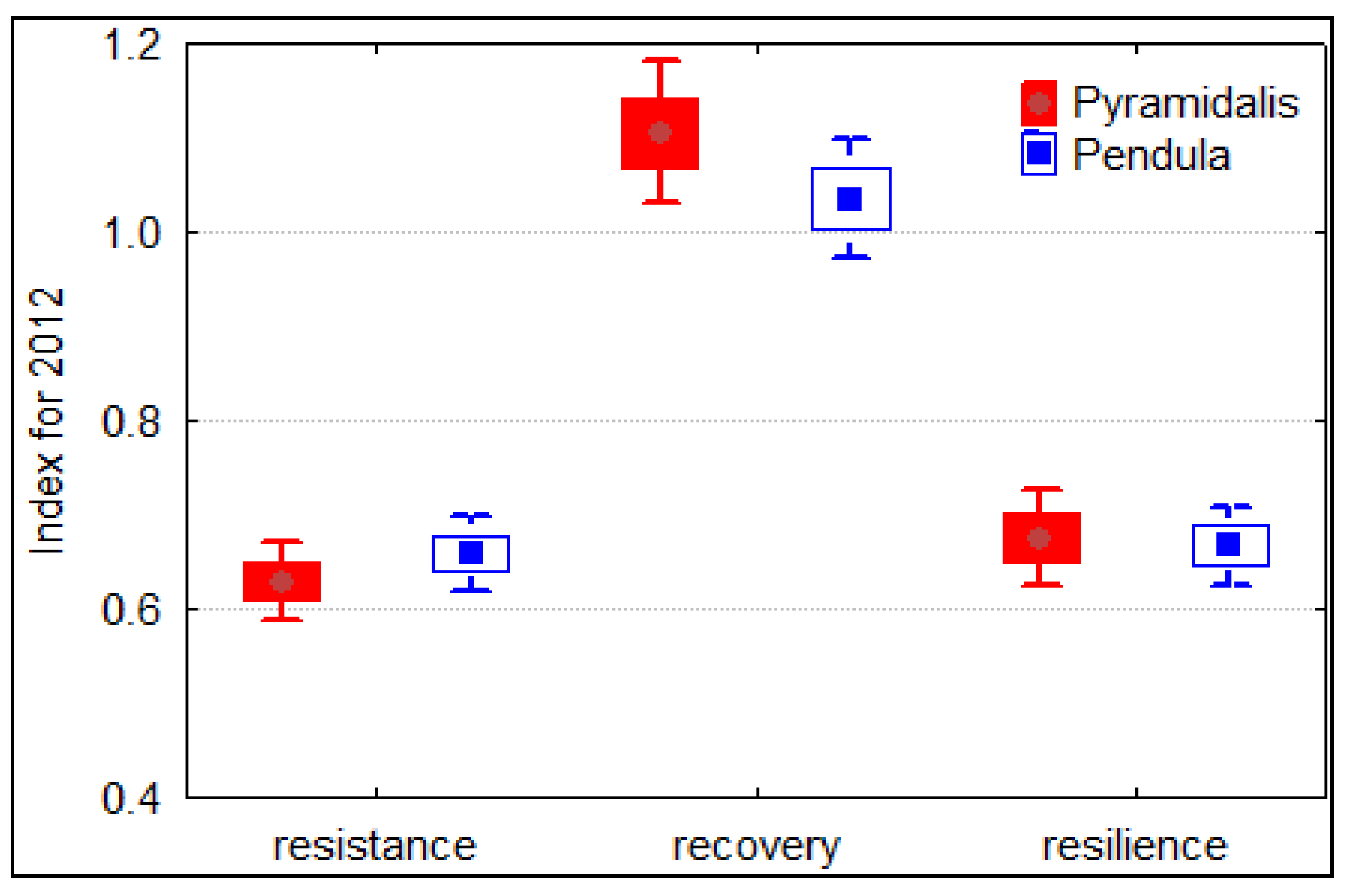
3. Results
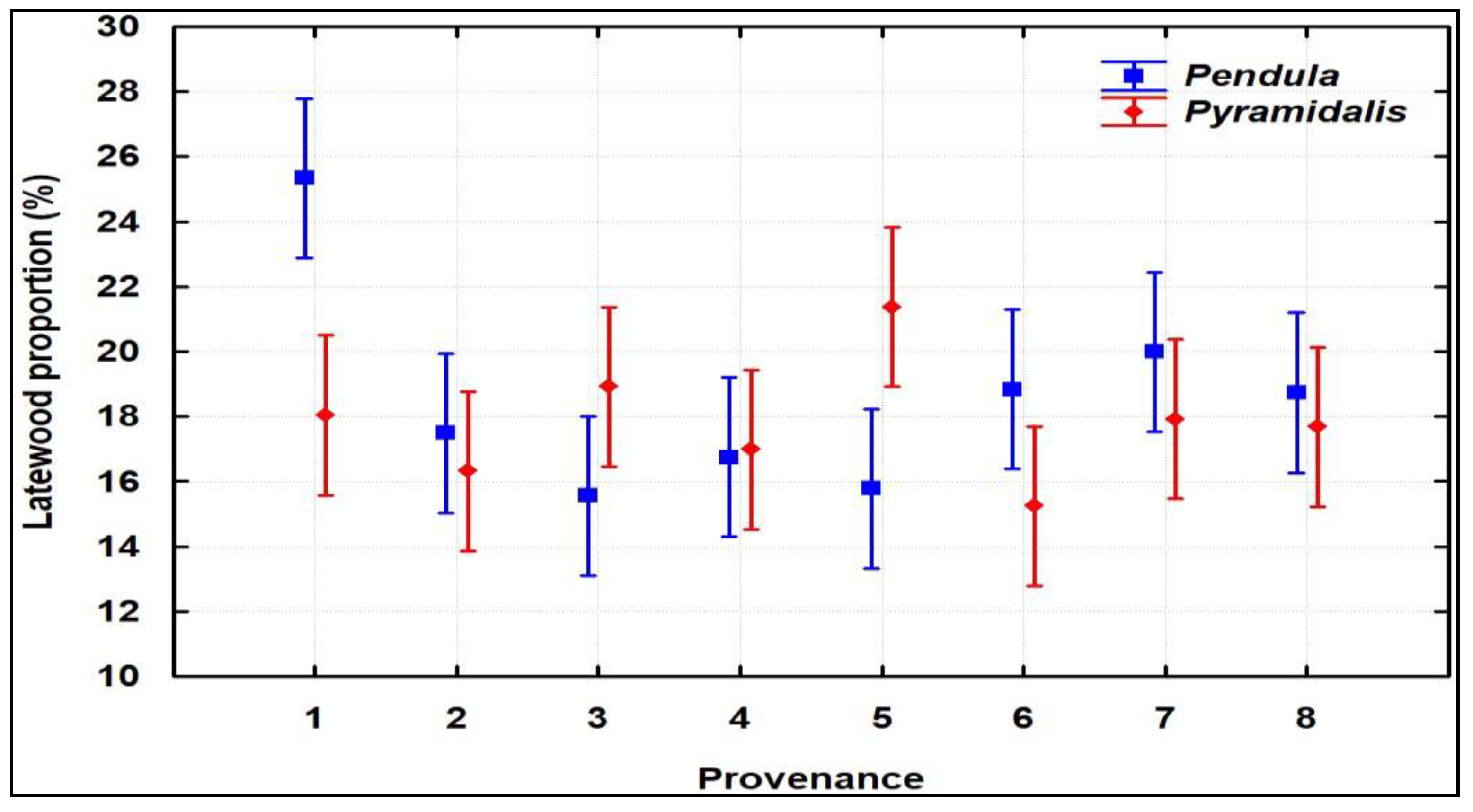
3.1. Phenotypic Variability, Heritability and Genetic Gain
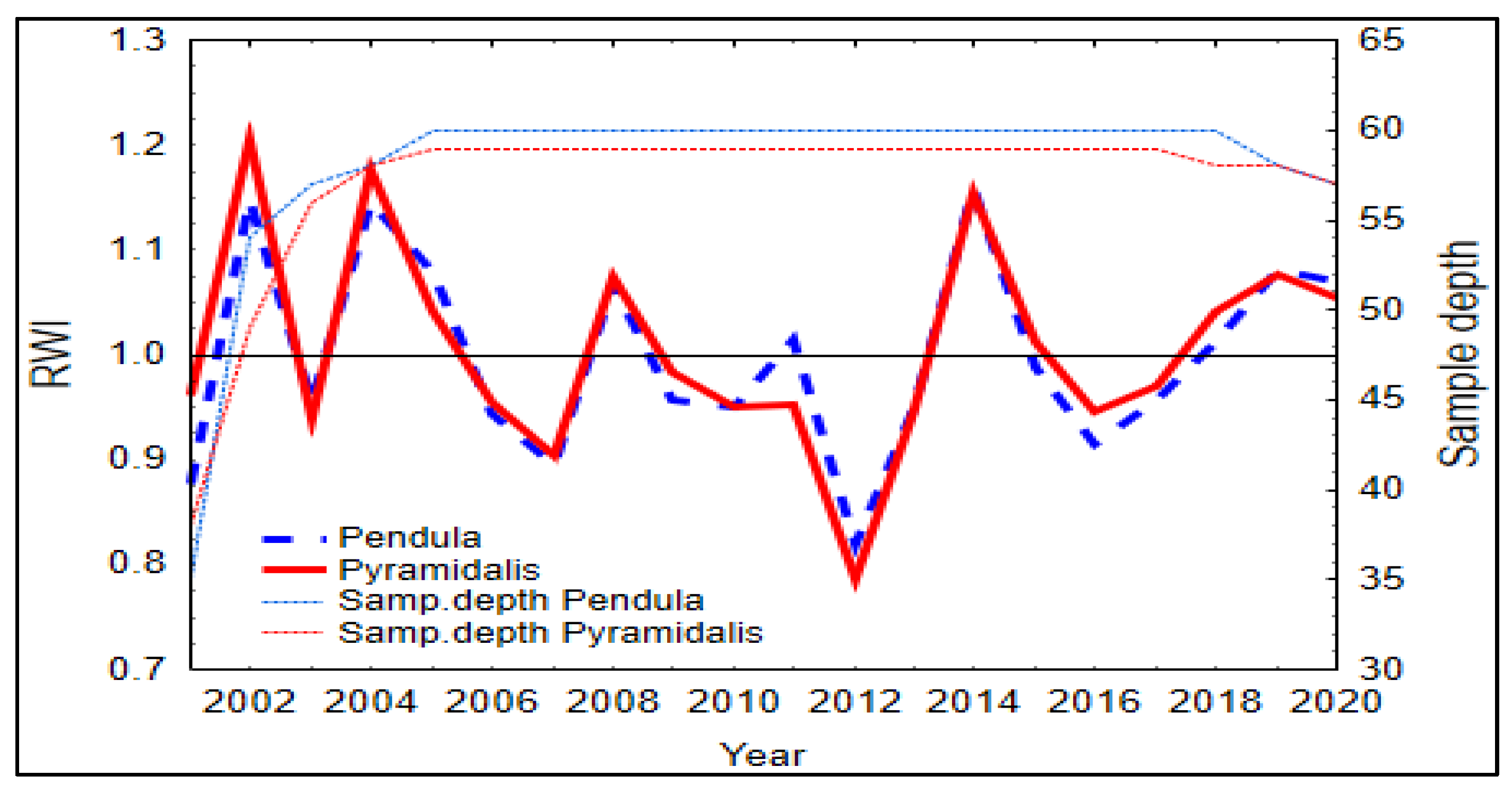
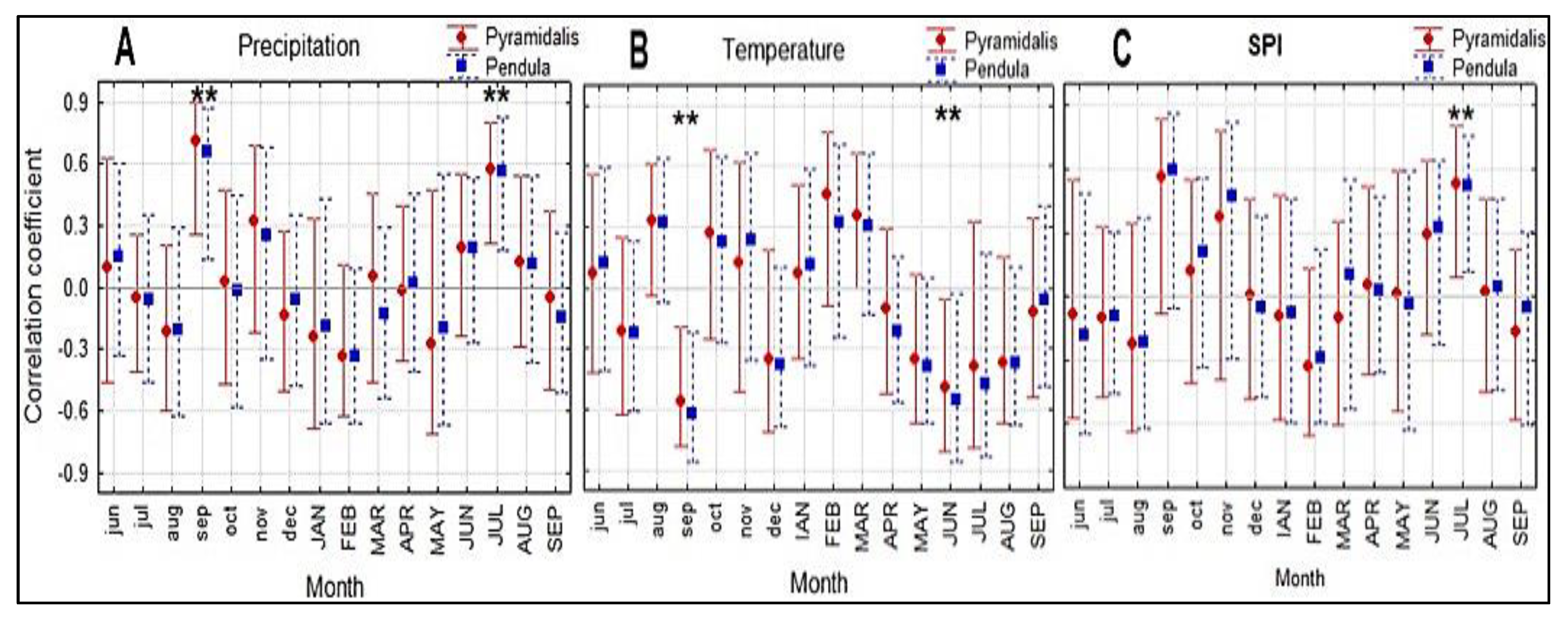
3.2. Climate and Growth Patterns of the Two Spruce Forms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Pretzsch, H.; Biber, P.; Schütze, G.; Uhl, E.; Rötzer, T. Forest stand growth dynamics in Central Europe have accelerated since 1870. Nat. Commun. 2014, 5, 4967. [Google Scholar] [CrossRef] [PubMed]
- Kauppi, P.E.; Posch, M.; Pirinen, P. Large Impacts of Climatic Warming on Growth of Boreal Forests since 1960. PLoS ONE 2014, 9, e111340. [Google Scholar] [CrossRef] [PubMed]
- Henttonen, H.M.; Nöjd, P.; Mäkinen, H. Environment-induced growth changes in the Finnish forests during 1971–2010—An analysis based on National Forest Inventory. For. Ecol. Manag. 2017, 386, 22–36. [Google Scholar] [CrossRef]
- Mensah, A.A.; Holmström, E.; Petersson, H.; Nyström, K.; Mason, E.G.; Nilsson, U. The millennium shift: Investigating the relationship between environment and growth trends of Norway spruce and Scots pine in northern Europe. For. Ecol. Manag. 2021, 481, 118727. [Google Scholar] [CrossRef]
- King, G.M.; Gugerli, F.; Fonti, P.; Frank, D. Tree growth response along an elevational gradient: Climate or genetics? Oecologia 2013, 173, 1587–1600. [Google Scholar] [CrossRef] [PubMed]
- Marcu, N.; Budeanu, M.; Apostol, E.N.; Radu, R.G. Valuation of the Economic Benefits from Using Genetically Improved Forest Reproductive Materials in Afforestation. Forests 2020, 11, 382. [Google Scholar] [CrossRef]
- Laubhann, D.; Sterba, H.; Reinds, G.J.; De Vries, W. The impact of atmospheric deposition and climate on forest growth in European monitoring plots: An individual tree growth model. For. Ecol. Manag. 2009, 258, 1751–1761. [Google Scholar] [CrossRef]
- Rohner, B.; Waldner, P.; Lischke, H.; Ferretti, M.; Thürig, E. Predicting individual-tree growth of central European tree species as a function of site, stand, management, nutrient, and climate effects. Eur. J. For. Res. 2018, 137, 29–44. [Google Scholar] [CrossRef]
- Kharuk, V.; Im, S.T.; Dvinskaya, M.L.; Golukov, A.S.; Ranson, K.J. Climate-induced mortality of spruce stands in Belarus. Environ. Res. Lett. 2015, 10, 125006. [Google Scholar] [CrossRef]
- Cienciala, E.; Tumajer, J.; Zatloukal, V.; Beranová, J.; Holá, Š.; Hůnová, I.; Russ, R. Recent spruce decline with biotic pathogen infestation as a result of interacting climate, deposition and soil variables. Eur. J. For. Res. 2017, 136, 307–317. [Google Scholar] [CrossRef]
- Holuša, J.; Lubojacký, J.; Čurn, V.; Tonka, T.; Lukášová, K.; Horák, J. Combined effects of drought stress and Armillaria infection on tree mortality in Norway spruce plantations. For. Ecol. Manag. 2018, 427, 434–445. [Google Scholar] [CrossRef]
- Sedmáková, D.; Sedmák, R.; Bosela, M.; Ježík, M.; Blazenec, M.; Hlásny, T.; Marušák, R. Growth-climate responses indicate shifts in the competitive ability of European beech and Norway spruce under recent climate warming in East-Central Europe. Dendrochronologia 2019, 54, 37–48. [Google Scholar] [CrossRef]
- Büntgen, U.; Frank, D.; Kaczka, R.J.; Verstege, A.; Zwijacz-Kozica, T.; Esper, J. Growth responses to climate in a multi-species tree-ring network in the Western Carpathian Tatra Mountains, Poland and Slovakia. Tree Physiol. 2007, 27, 689–702. [Google Scholar] [CrossRef] [PubMed]
- Treml, V.; Ponocná, T.; Büntgen, U. Growth trends and temperature responses of treeline Norway spruce in the Czech-Polish Sudetes Mountains. Clim. Res. 2012, 55, 91–103. [Google Scholar] [CrossRef]
- Sidor, C.G.; Popa, I.; Vlad, R.; Cherubini, P. Different tree-ring responses of Norway spruce to air temperature across an altitudinal gradient in the Eastern Carpathians (Romania). Trees 2015, 29, 985–997. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.-J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef]
- Hlásny, T.; Turčáni, M. Persisting bark beetle outbreak indicates the unsustainability of secondary Norway spruce forests: Case study from Central Europe. Ann. For. Sci. 2013, 70, 481–491. [Google Scholar] [CrossRef]
- Budeanu, M.; Apostol, E.N.; Popescu, F.; Postolache, D.; Ioniţă, L. Testing of the narrow crowned Norway spruce ideotype (Picea abies f. pendula) and the hybrids with normal crown form (pyramidalis) in multisite comparative trials. Sci. Total. Environ. 2019, 689, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Bosela, M.; Kulla, L.; Roessiger, J.; Šebeň, V.; Dobor, L.; Büntgen, U.; Lukac, M. Long-term effects of environmental change and species diversity on tree radial growth in a mixed European forest. For. Ecol. Manag. 2019, 446, 293–303. [Google Scholar] [CrossRef]
- Franceschini, T.; Bontemps, J.-D.; Gelhaye, P.; Rittié, D.; Hervé, J.-C.; Gégout, J.-C.; Leban, J.-M. Decreasing trend and fluctuations in the mean ring density of Norway spruce through the twentieth century. Ann. For. Sci. 2010, 67, 816. [Google Scholar] [CrossRef][Green Version]
- Franceschini, T.; Bontemps, J.-D.; Leban, J.-M. Transient historical decrease in earlywood and latewood density and unstable sensitivity to summer temperature for Norway spruce in northeastern France. Can. J. For. Res. 2012, 42, 219–226. [Google Scholar] [CrossRef]
- Semeniuc, A.I.; Popa, I. Comparative analysis of tree ring parameters Variation in four coniferous species: (Picea abies, Abies alba, Pinus sylvestris and Larix decidua). Int. J. Conserv. Sci. 2018, 9, 591–598. [Google Scholar]
- Vlad, R.; Zhiyanski, M.; Dincă, L.; Sidor, C.G.; Constandache, C.; Pei, G.; Ispravnic, A.; Blaga, T. Assessment of the density of wood with stem decay of Norway spruce trees using drill resistance. Proc. Bulg. Acad. Sci. 2018, 71, 1502–1510. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Klein, T.; Bartlett, M.; Sack, L.; Pellegrini, A.; Choat, B.; Jansen, S. Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought-induced tree mortality across the globe. Proc. Natl. Acad. Sci. USA 2016, 113, 5024–5029. [Google Scholar] [CrossRef] [PubMed]
- Jupa, R.; Plavcová, L.; Gloser, V.; Jansen, S. Linking xylem water storage with anatomical parameters in five temperate tree species. Tree Physiol. 2016, 36, 756–769. [Google Scholar] [CrossRef] [PubMed]
- Steffenrem, A.; Solheim, H.; Skrøppa, T. Genetic parameters for wood quality traits and resistance to the pathogens Heterobasidion parviporum and Endoconidiophora polonica in a Norway spruce breeding population. Eur. J. For. Res. 2016, 135, 815–825. [Google Scholar] [CrossRef]
- Zeltiņš, P.; Katrevičs, J.; Gailis, A.; Maaten, T.; Bāders, E.; Jansons, Ā. Effect of Stem Diameter, Genetics, and Wood Properties on Stem Cracking in Norway Spruce. Forests 2018, 9, 546. [Google Scholar] [CrossRef]
- Karki, L. Genetically narrow-crowned trees combine high timber quality and high stem wood production at low cost. In Crop. Physiology of Forest Trees; Landsberg, J.J., Ed.; Tigerstedt Publishing House: Helsinki, Finland, 1985; pp. 245–256. [Google Scholar]
- Gerendiain, A.Z.; Peltola, H.; Pulkkinen, P.; Ikonen, V.-P.; Jaatinen, R. Differences in growth and wood properties between narrow and normal crowned types of Norway spruce grown at narrow spacing in Southern Finland. Silva. Fenn. 2008, 42, 423–437. [Google Scholar] [CrossRef]
- Gerendiain, A.Z.; Peltola, H.; Pulkkinen, P. Growth and wood property traits in narrow crowned Norway spruce (Picea abies f. pendula) clones grown in southern Finland. Silva. Fenn. 2009, 43, 369–382. [Google Scholar] [CrossRef]
- Pârnuţă, G. Variabilitatea Genetică și Ameliorarea Arborilor de Molid cu Coroană Îngustă în România (Genetic Variability and Breeding of Narrow-Crown Spruce Trees in Romania); Silvică Publishing House: Bucharest, Romania, 2008; 181p, (In Romanian with English Abstract). [Google Scholar]
- Apostol, E.N.; Budeanu, M. Adaptability of Narrow-Crowned Norway Spruce Ideotype (Picea abies (L.) Karst. pendula Form) in 25 Years Half-Sib Comparative Trials in the Eastern Carpathians. Forests 2019, 10, 395. [Google Scholar] [CrossRef]
- Anonymous. Management planning of production unit IV Suzana, forest district Măneciu. 2018. [Google Scholar]
- CRU Dataset. Climatic Research Unit, University of East, England. 2021. Available online: www.cru.uea.ac.uk (accessed on 30 April 2021).
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological maps for the main European woody species. Data Brief. 2017, 12, 662–666. [Google Scholar] [CrossRef]
- Cybis Elektronik & Data AB. Cybis Dendrochronology and History. Cybis Electronic and Data, Saltsjöbaden, Sweden. 2020. Available online: www.cybis.se (accessed on 30 April 2021).
- Popa, I. Fundamente Metodologice și Aplicații de Dendrocronologie (Methodological Fundamentals and Applications of Dendrochronology); Ed. Silvică: Câmpulung Moldovenesc, Romania, 2004; p. 200. Available online: http://www.editurasilvica.ro/carti/popa1/integral.pdf (accessed on 2 May 2021).
- Yamaguchi, D.K.; Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology, Applications in the Environmental Sciences. Arct. Alp. Res. 1991, 23, 120. [Google Scholar] [CrossRef]
- Leca, Ş. Creșterea Arborilor și Arboretelor în Sistemul de Monitorizare Forestieră Intensivă (Trees and Stands Growth in Intensive Forest Monitoring System). Ph.D. Thesis, Transylvania University, Brasov, Romania, 2014. [Google Scholar]
- Nechita, C.; Popa, I.; Eggertsson, Ó. Climate response of oak (Quercus spp.), an evidence of a bioclimatic boundary induced by the Carpathians. Sci. Total Environ. 2017, 599–600, 1598–1607. [Google Scholar] [CrossRef] [PubMed]
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; Mérian, P.; Qeadan, F.; Zang, C.; Pucha-Cofrep, D.; Wernicke, J. dplR: Dendrochronology Program Library in R; R Package Version 1.6.9; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.r-project.org/ (accessed on 5 May 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.eea.europa.eu/data-and-maps/indicators/oxygen-consuming-substances-in-rivers/r-development-core-team-2006 (accessed on 5 May 2021).
- Satmari, A. Lucrări Practice de Biogeografie (Practical Applications of Biogeography). Available online: http://www.academia.edu/9909429/05_INDICI_ECOMETRICI (accessed on 2 May 2021).
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas mit den Alpen. In Ökologischer, Dynamischer und Historischer Sicht (Vegetation of Central Europe with the Alps: From an Ecological, Dynamic and Historical Point of View), 6th ed.; Ulmer Verlag: Stuttgart, Germany, 2010; p. 1334. [Google Scholar]
- Edwards, D.C. Characteristics of 20th Century Drought in the United States at Multiple Time Scales. Master’s Thesis, Colorado State University, Fort Collins, CO, USA, 1997; p. 155. [Google Scholar]
- Salehnia, N.; Alizadeh, A.; Sanaeinejad, H.; Bannayan, M.; Zarrin, A.; Hoogenboom, G. Estimation of meteorological drought indices based on AgMERRA precipitation data and station-observed precipitation data. J. Arid. Land 2017, 9, 797–809. [Google Scholar] [CrossRef]
- Trujillo-Moya, C.; George, J.-P.; Fluch, S.; Geburek, T.; Grabner, M.; Karanitsch-Ackerl, S.; Konrad, H.; Mayer, K.; Sehr, E.; Wischnitzki, E.; et al. Drought Sensitivity of Norway Spruce at the Species’ Warmest Fringe: Quantitative and Molecular Analysis Reveals High Genetic Variation Among and Within Provenances. G3 Genes|Genomes|Genetics 2018, 8, 1225–1245. [Google Scholar] [CrossRef] [PubMed]
- Zang, C.; Biondi, F. treeclim: An R package for the numerical calibration of proxy-climate relationships. Ecography 2015, 38, 431–436. [Google Scholar] [CrossRef]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixedversuspure forests: Evidence of stress release by inter-specific facilitation. Plant. Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef]
- Van Der Maaten-Theunissen, M.; Van Der Maaten, E.; Bouriaud, O. pointRes: An R package to analyze pointer years and components of resilience. Dendrochronologia 2015, 35, 34–38. [Google Scholar] [CrossRef]
- Nanson, A. Génétique et Amélioration Des. Arbres Forestières (Genetic and Forest Trees Breeding); Les presses agronomique de Gembloux: Gembloux, Belgium, 2004; p. 712. (In French) [Google Scholar]
- Falconer, D.S. Introduction to Quantitative Genetics, 2nd ed.; Longmans Green: London, UK; New York, NY, USA, 1981. [Google Scholar]
- Abengmeneng, C.S.; Ofori, D.A.; Kumapley, P.; Akromah, R.; Jamnadass, R. Estimation of heritability and genetic gain in height growth in Ceiba pentandra. Afr. J. Biotechnol. 2015, 14, 1880–1885. [Google Scholar] [CrossRef]
- Breed, R. An Open Statistical Package to Analyse Genetic Data (WP6). 2016. Available online: http://famuvie.github.io/breedR/ (accessed on 5 May 2021).
- Hacke, U.G.; Stiller, V.; Sperry, J.S.; Pittermann, J.; McCulloh, K.A. Cavitation Fatigue. Embolism and Refilling Cycles Can Weaken the Cavitation Resistance of Xylem. Plant. Physiol. 2001, 125, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Wolkerstorfer, S.; Rosner, S.; Hietz, P. Assessment of vulnerability to cavitation in small wood samples of Picea abies (L. Karst.). For. Ideas 2010, 16, 83–89. [Google Scholar]
- Şofletea, N.; Curtu, A.L. Dendrologie (Dendrology); Transylvania University Publishing House: Brasov, Romania, 2007; p. 540. [Google Scholar]
- Stănescu, V.; Șofletea, N. Cercetări de genetică ecologică în molidișuri montane (II) (Ecological genetics research in mountain spruces (II)). Revista Pădurilor 1992, 107, 2–5. [Google Scholar]
- Şofletea, N.; Curtu, A.L.; Daia, M.L.; Budeanu, M. The dynamics and variability of radial growth in provenance trials of Norway spruce (Picea abies (L.) Karst.) within and beyond the hot margins of its natural range. Not. Bot. Horti Agrobo. 2015, 43, 265–271. [Google Scholar] [CrossRef]
- Lundqvist, S.-O.; Seifert, S.; Grahn, T.; Olsson, L.; García-Gil, M.R.; Karlsson, B.; Seifert, T. Age and weather effects on between and within ring variations of number, width and coarseness of tracheids and radial growth of young Norway spruce. Eur. J. For. Res. 2018, 137, 719–743. [Google Scholar] [CrossRef]
- Irbe, I.; Sable, I.; Noldt, G.; Grinfelds, U.; Jansons, A.; Treimanis, A.; Koch, G. Wood and Tracheid Properties of Norway Spruce (Picea abies [L] Karst.) Clones Grown on Former Agricultural Land in Latvia. Balt. For. 2015, 21, 114–123. [Google Scholar]
- Chen, Z.-Q.; Hai, H.N.T.; Helmersson, A.; Liziniewicz, M.; Hallingbäck, H.R.; Fries, A.; Berlin, M.; Wu, H.X. Advantage of clonal deployment in Norway spruce (Picea abies (L.) H. Karst). Ann. For. Sci. 2020, 77, 14. [Google Scholar] [CrossRef]
- Șofletea, N. Genetică și Ameliorarea Arborilor (Genetics and Trees Breeding); Pentru viață Publishing House: Brașov, Romania, 2005; p. 455. (In Romanian) [Google Scholar]
- Castagneri, D.; Vacchiano, G.; Hacket-Pain, A.; DeRose, R.J.; Klein, T.; Bottero, A. Meta-analysis Reveals Different Competition Effects on Tree Growth Resistance and Resilience to Drought. Ecosystems 2021, 24, 1–14. [Google Scholar] [CrossRef]
- Gömöry, D.; Longauer, R.; Hlásny, T.; Pacalaj, M.; Strmeň, S.; Krajmerová, D. Adaptation to common optimum in different populations of Norway spruce (Picea abies Karst.). Eur. J. For. Res. 2011, 131, 401–411. [Google Scholar] [CrossRef]
- Kapeller, S.; Lexer, M.J.; Geburek, T.; Hiebl, J.; Schueler, S. Intraspecific variation in climate response of Norway spruce in the eastern Alpine range: Selecting appropriate provenances for future climate. For. Ecol. Manag. 2012, 271, 46–57. [Google Scholar] [CrossRef]
- Klisz, M.; Ukalska, J.; Koprowski, M.; Tereba, A.; Puchałka, R.; Przybylski, P.; Jastrzębowski, S.; Nabais, C. Effect of provenance and climate on intra-annual density fluctuations of Norway spruce Picea abies (L.) Karst. in Poland. Agric. For. Meteorol. 2019, 269, 145–156. [Google Scholar] [CrossRef]
- Burczyk, J.; Giertych, M. Response of Norway spruce (Picea abies [L] Karst) annual increments to drought for various provenances and locations. Silvae Genet. 1991, 40, 146–152. [Google Scholar]
- Budeanu, M.; Petritan, A.M.; Popescu, F.; Vasile, D.; Tudose, N.C. The resistance of European beech (Fagus sylvatica) from the eastern natural limit of species to climate change. Not. Bot. Horti Agrobo. 2016, 44, 625–633. [Google Scholar] [CrossRef]
- Kolář, T.; Čermák, P.; Trnka, M.; Žid, T.; Rybníček, M. Temporal changes in the climate sensitivity of Norway spruce and European beech along an elevation gradient in Central Europe. Agric. For. Meteorol. 2017, 239, 24–33; [Google Scholar] [CrossRef]
- Gyllenstrand, N.; Clapham, D.; Källman, T.; Lagercrantz, U.; Richardt, S.; Lang, D.; Reski, R.; Frank, W.; Rensing, S.A. A Norway spruce FLOWERING LOCUS T homolog is implicated in control of growth rhythm in conifers. Plant. Physiol. 2007, 144, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Hayatgheibi, H.; Haapanen, M.; Lundströmer, J.; Berlin, M.; Kärkkäinen, K.; Helmersson, A. Impact of Drought Stress on Height Growth of Young Norway Spruce Full-Sib and Half-Sib Clonal Trials in Sweden and Finland. Forests 2021, 12, 498. [Google Scholar] [CrossRef]
- Obladen, N.; Dechering, P.; Skiadaresis, G.; Tegel, W.; Keßler, J.; Höllerl, S.; Kaps, S.; Hertel, M.; Dulamsuren, C.; Seifert, T.; et al. Tree mortality of European beech and Norway spruce induced by 2018–2019 hot droughts in central Germany. Agric. For. Meteorol. 2021, 307, 108482. [Google Scholar] [CrossRef]
- Dincă, L.; Murariu, G.; Iticescu, C.; Budeanu, M.; Murariu, A. Norway spruce (Picea abies (L.) Karst.) smart forests from the southern Carpathians. Int. J. Conserv. Sci. 2019, 10, 781–790. [Google Scholar]
- Teodosiu, M.; Mihai, G.; Fussi, B.; Ciocîrlan, E. Genetic diversity and structure of Silver fir (Abies alba Mill.) at the south-eastern limit of its distribution range. Ann. For. Res. 2019, 62, 139–156. [Google Scholar] [CrossRef]
- Zwetsloot, M.J.; Bauerle, T.L. Repetitive seasonal drought causes substantial species-specific shifts in fine-root longevity and spatio-temporal production patterns in mature temperate forest trees. New Phytol. 2021. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Provenance | Coordinates (N latitude/E Longitude/Altitude (m)) | T (°C)/P (mm)/AI |
|---|---|---|
| 1. Stâna de Vale I | 46°41′/22°38′/1200 | 4.0/1200/86 |
| 2. Stâna de Vale II | 46°46′/22°36′/1225 | 4.0/1200/86 |
| 3. Izbuc I | 46°36′/22°46′/1200 | 4.0/1200/86 |
| 4. Izbuc II | 46°38′/22°50′/1275 | 4.0/1200/86 |
| 5. Cetăţile Ponorului | 46°34′/22°42′/1050 | 4.5/1100/76 |
| 6. Bozovici | 44°57′/21°57′/600 | 9.5/770/39 |
| 7. Horoaba | 45°23′/25°25′/1675 | 2.8/1200/94 |
| 8. Cucureasa | 47°23′/25°04′/935 | 4.5/960/66 |
| Crown Form | RW mm | EW mm | LW mm | LWP % | Cores Signal | |||
|---|---|---|---|---|---|---|---|---|
| Ar 1 | IC | EPS | Rbar | |||||
| Pendula | 4.580 ± 0.67 | 3.733 ± 0.60 | 0.848 ± 0.27 | 18.6 ± 5.2 | 0.739 | 0.455 | 0.953 | 0.251 |
| Pyramidalis | 4.495 ± 0.86 | 3.696 ± 0.75 | 0.799 ± 0.25 | 17.8 ± 4.4 | 0.732 | 0.445 | 0.946 | 0.244 |
| Trial | 4.538 ± 0.77 | 3.714 ± 0.68 | 0.823 ± 0.26 | 18.2 ± 4.8 | 0.737 | 0.490 | 0.975 | 0.271 |
| Crown Form | CVRW | CVEW | CVLW | CVLWP | CVRW 2001–2010 | CVRW 2011–2020 | CVLWP 2001–2010 | CVLWP 2011–2020 |
|---|---|---|---|---|---|---|---|---|
| Pendula | 14.5 | 16.2 | 32.1 | 27.8 | 15.1 | 35.8 | 36.8 | 32.6 |
| Pyramidalis | 19.1 | 20.4 | 31.5 | 24.7 | 19.4 | 41.3 | 31.5 | 32.1 |
| Trial | 16.9 | 18.3 | 31.9 | 26.4 | 17.3 | 38.6 | 35.1 | 32.6 |
| Măneciu Trial | RW | EW | LW | LWP |
|---|---|---|---|---|
| All families | ||||
| Heritability | 0.39 | 0.27 | 0.52 | 0.34 |
| Genetic gain | 3.4 | 1.7 | 13.0 | 4.9 |
| Pendula families | ||||
| Heritability | 0.15 | 0.03 | 0.56 | 0.36 |
| Genetic gain | 0.3 | 0.1 | 13.3 | 4.4 |
| Pyramidalis families | ||||
| Heritability | 0.54 | 0.43 | 0.49 | 0.33 |
| Genetic gain | 5.5 | 3.9 | 7.8 | 2.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budeanu, M.; Apostol, E.N.; Besliu, E.; Crișan, V.E.; Petritan, A.M. Phenotypic Variability and Differences in the Drought Response of Norway Spruce Pendula and Pyramidalis Half-Sib Families. Forests 2021, 12, 947. https://doi.org/10.3390/f12070947
Budeanu M, Apostol EN, Besliu E, Crișan VE, Petritan AM. Phenotypic Variability and Differences in the Drought Response of Norway Spruce Pendula and Pyramidalis Half-Sib Families. Forests. 2021; 12(7):947. https://doi.org/10.3390/f12070947
Chicago/Turabian StyleBudeanu, Marius, Ecaterina Nicoleta Apostol, Emanuel Besliu, Vlad Emil Crișan, and Any Mary Petritan. 2021. "Phenotypic Variability and Differences in the Drought Response of Norway Spruce Pendula and Pyramidalis Half-Sib Families" Forests 12, no. 7: 947. https://doi.org/10.3390/f12070947
APA StyleBudeanu, M., Apostol, E. N., Besliu, E., Crișan, V. E., & Petritan, A. M. (2021). Phenotypic Variability and Differences in the Drought Response of Norway Spruce Pendula and Pyramidalis Half-Sib Families. Forests, 12(7), 947. https://doi.org/10.3390/f12070947