
Responses of Visual Forms to Neighboring Competition: A Structural Equation Model for Cotinus coggygria var. cinerea Engl.



Abstract
:1. Introduction
2. Materials and Methodology
2.1. Study Area and Data Collection
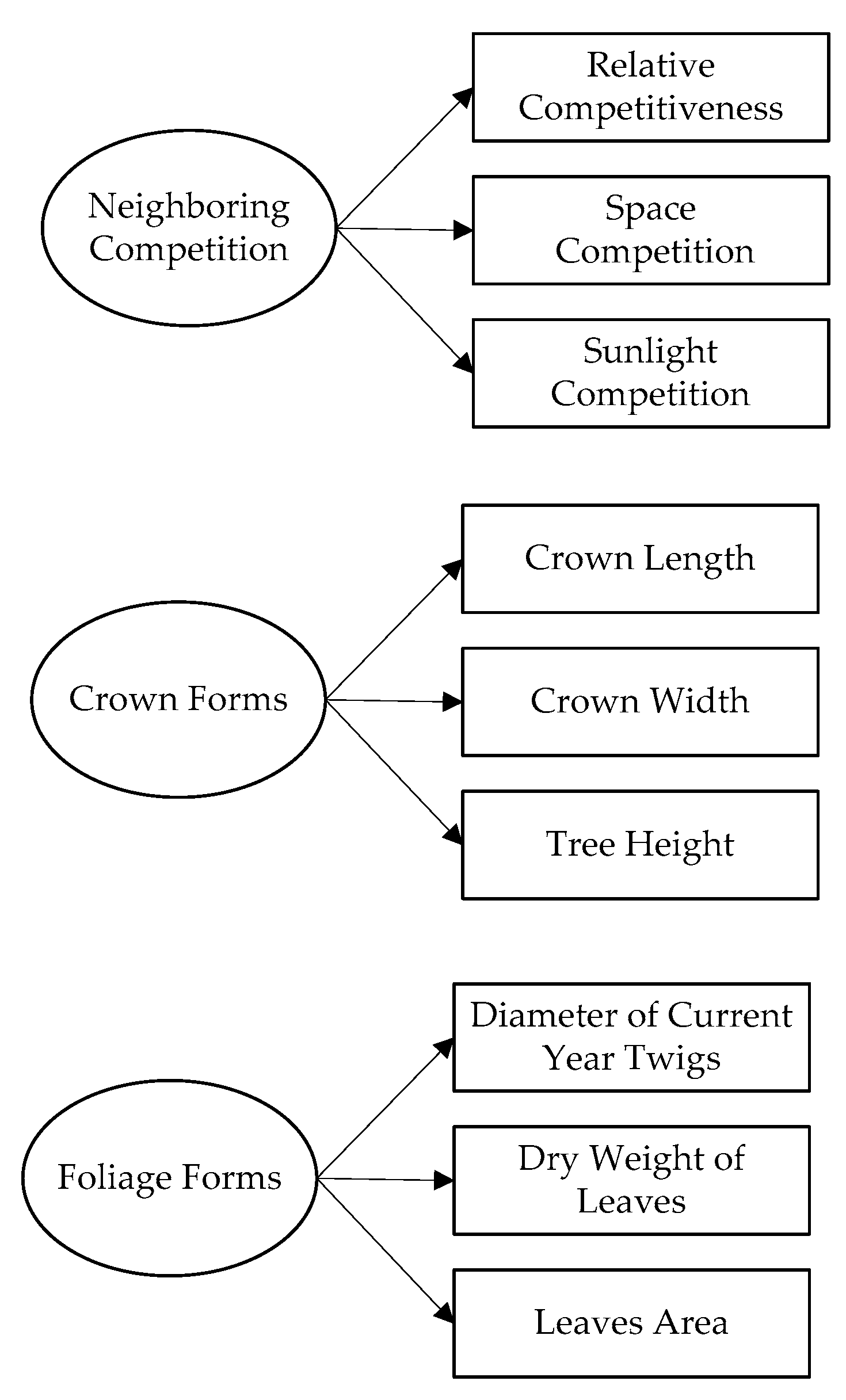
2.2. Overview of Our Approach
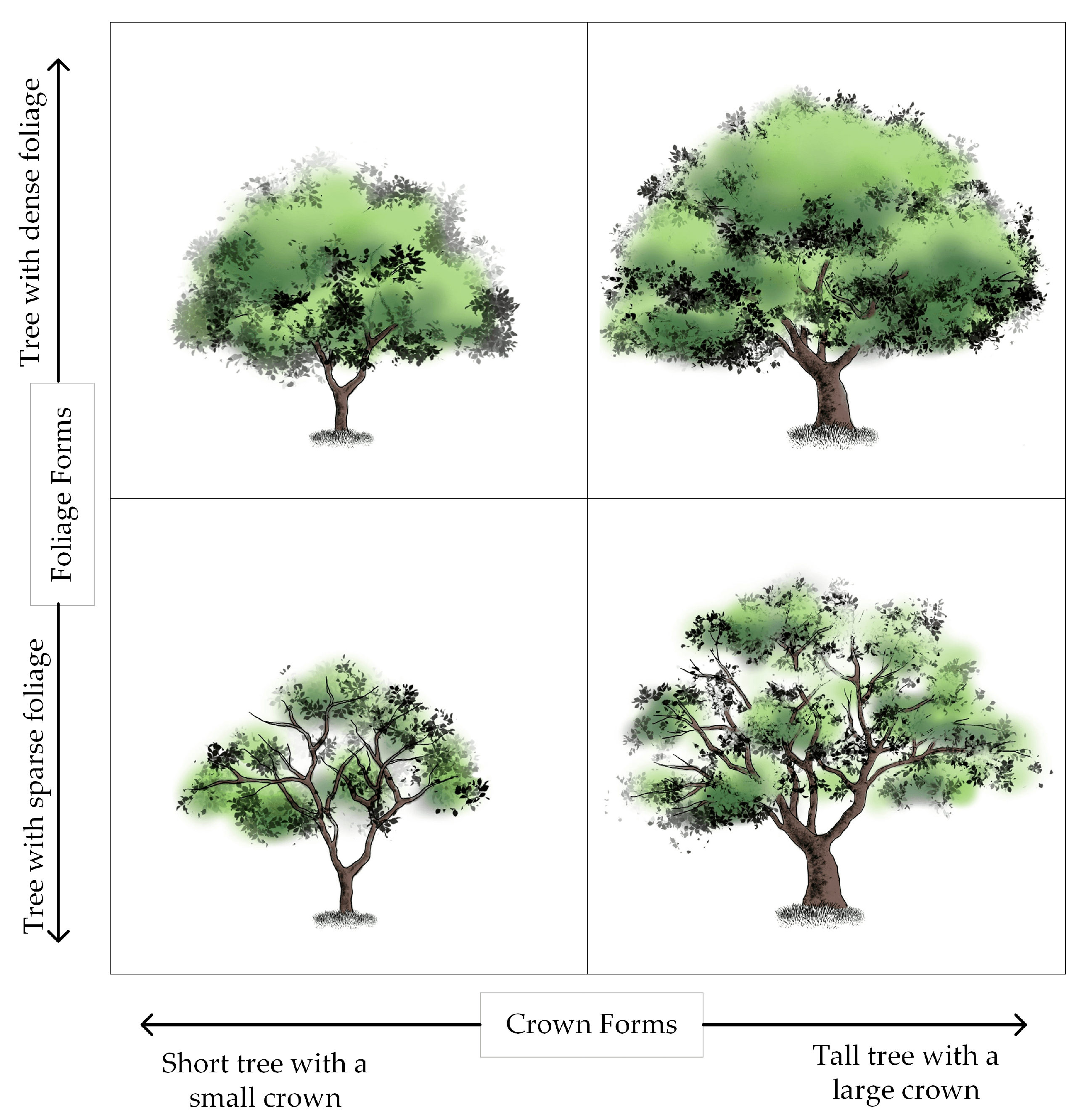
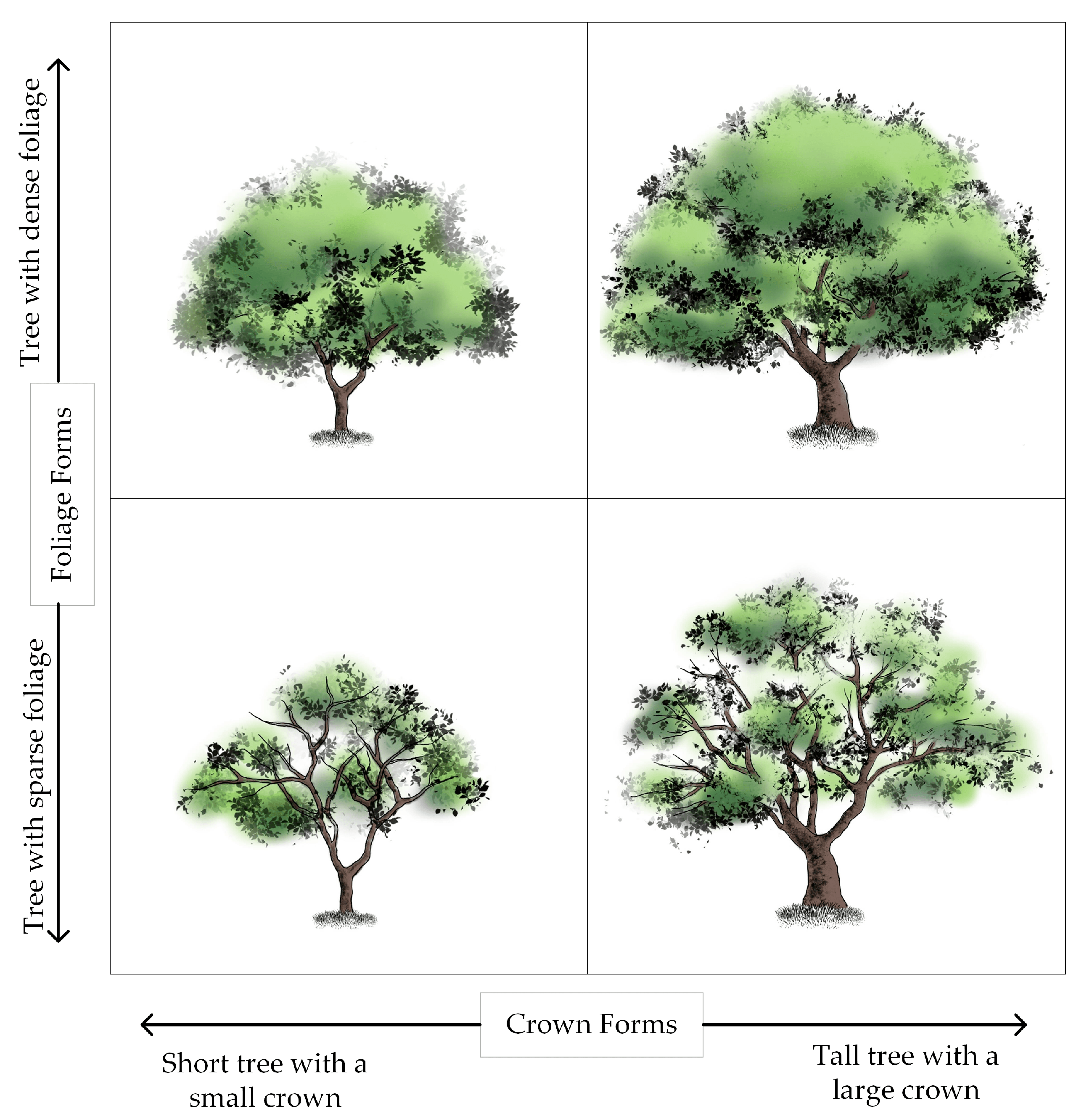
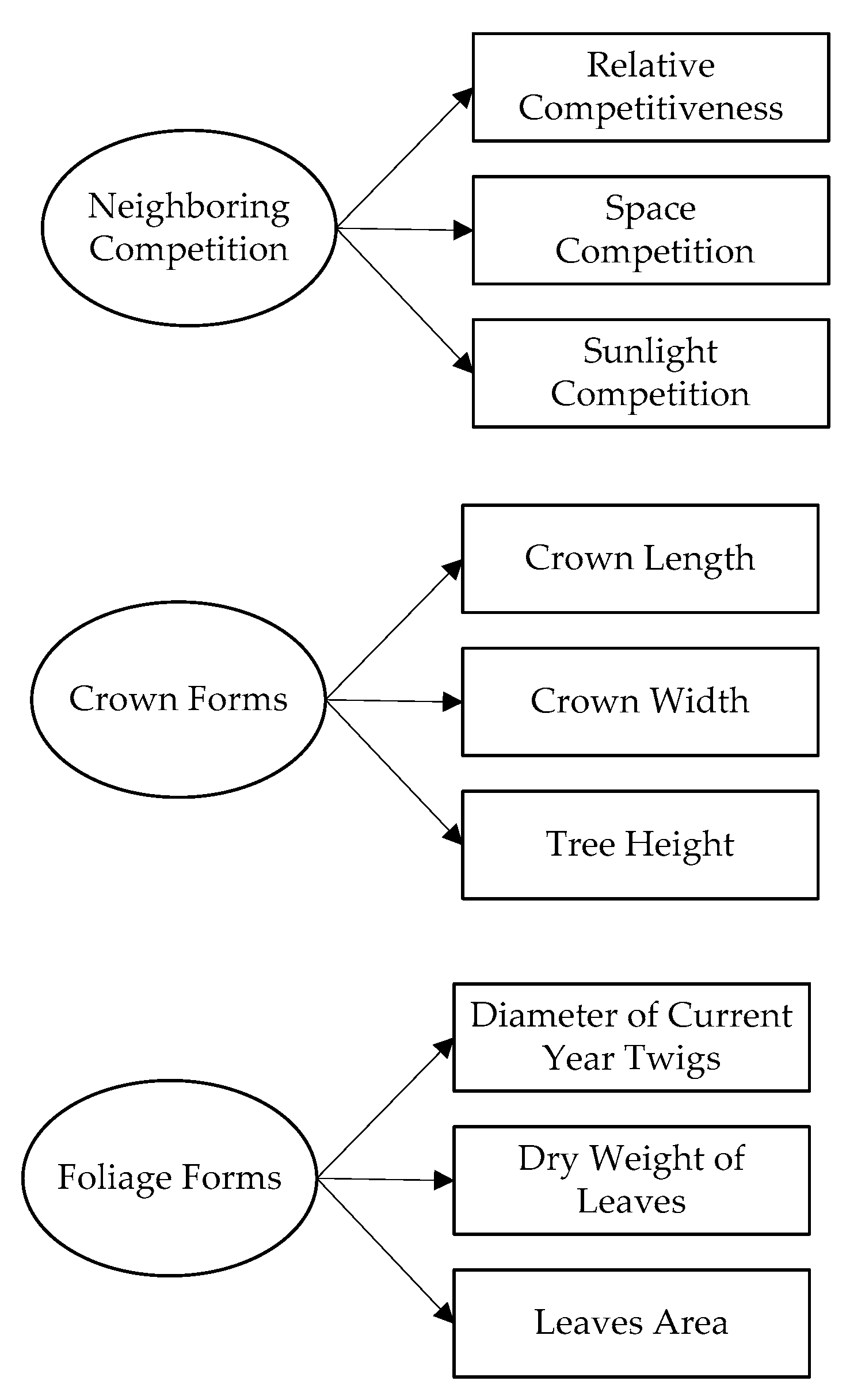
2.3. Visual Forms of the Individual Tree
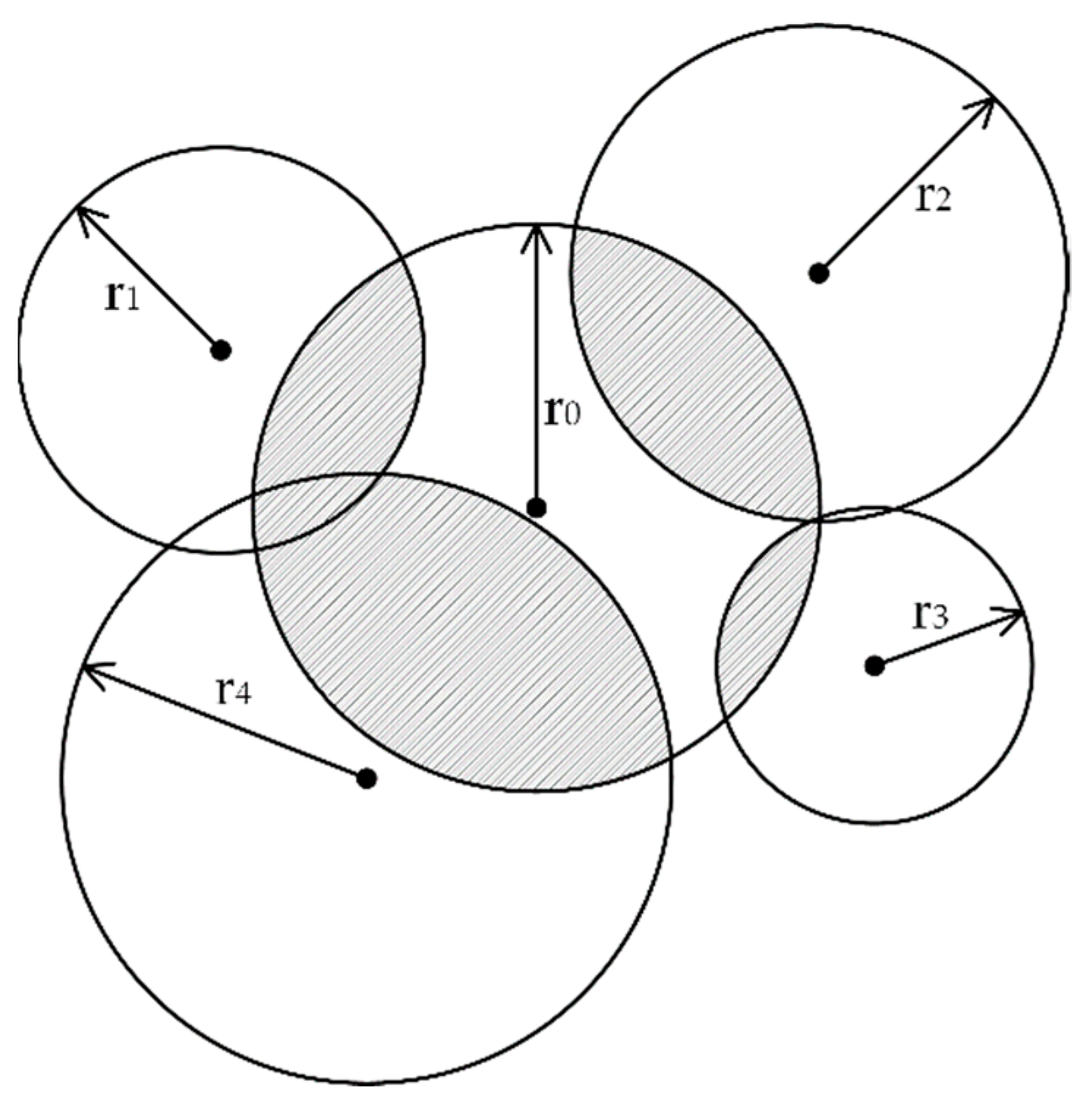
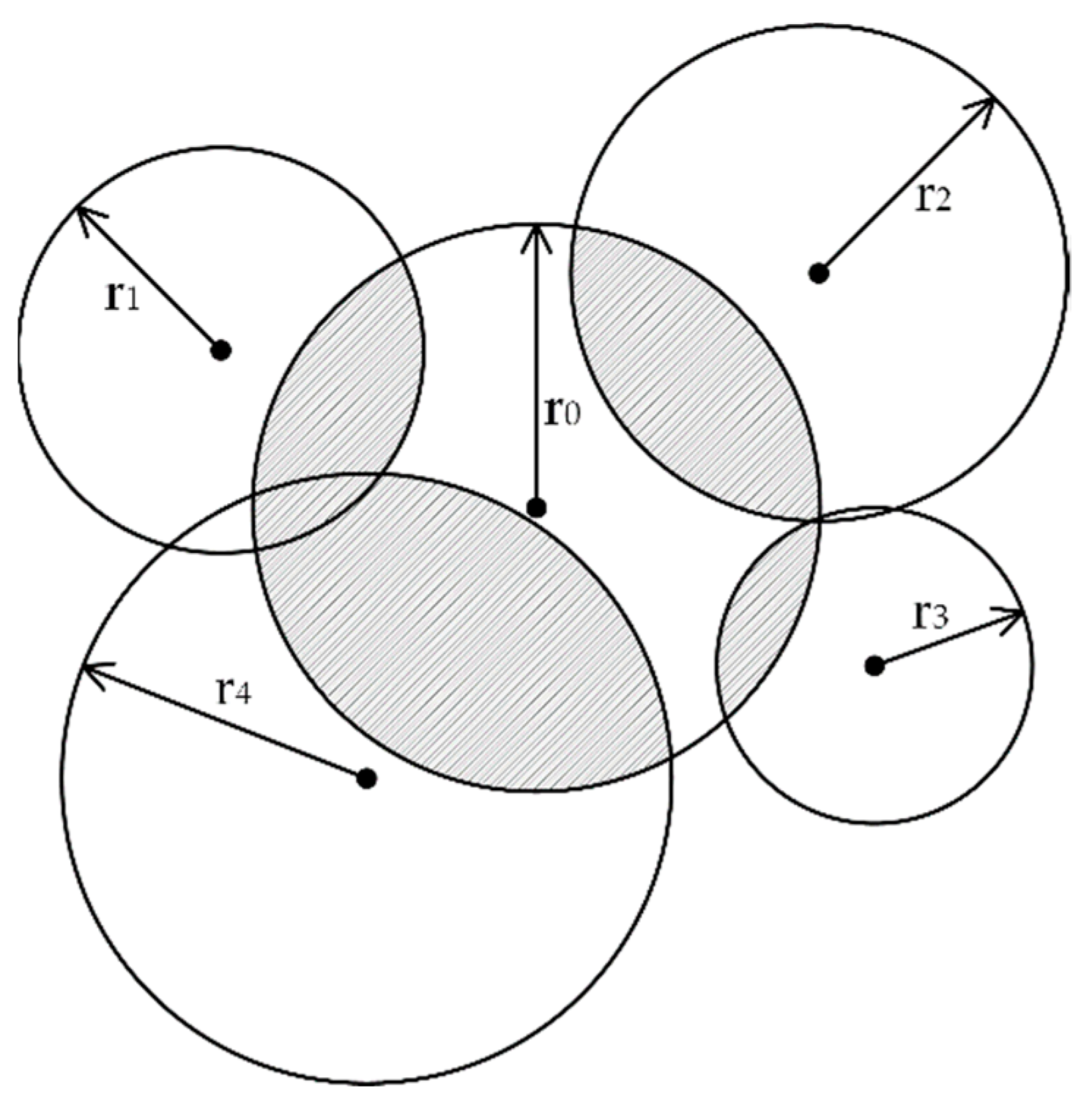
2.4. Neighboring Competition
2.5. Hypotheses
2.6. SEM and Measurement Models
3. Results
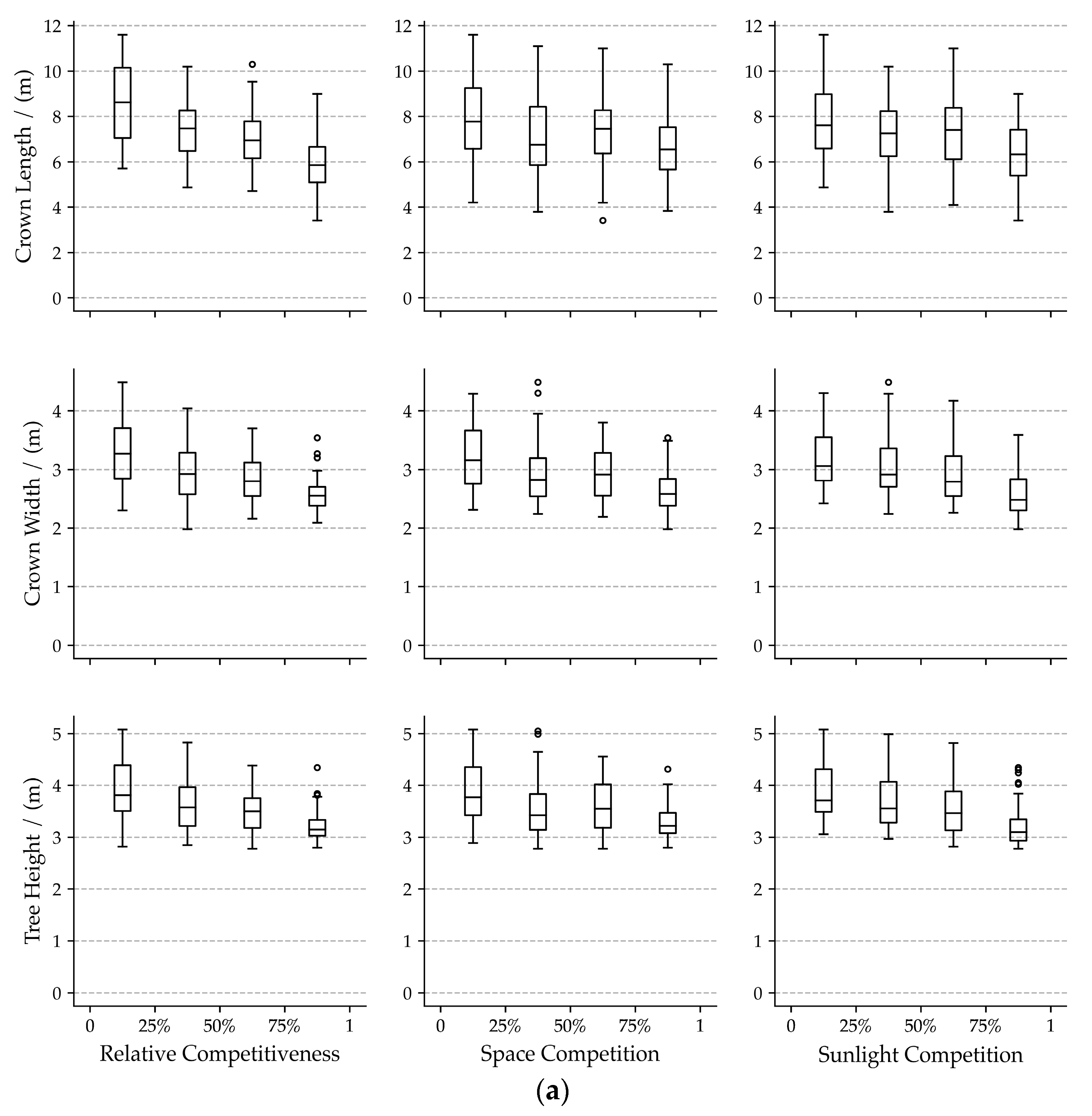
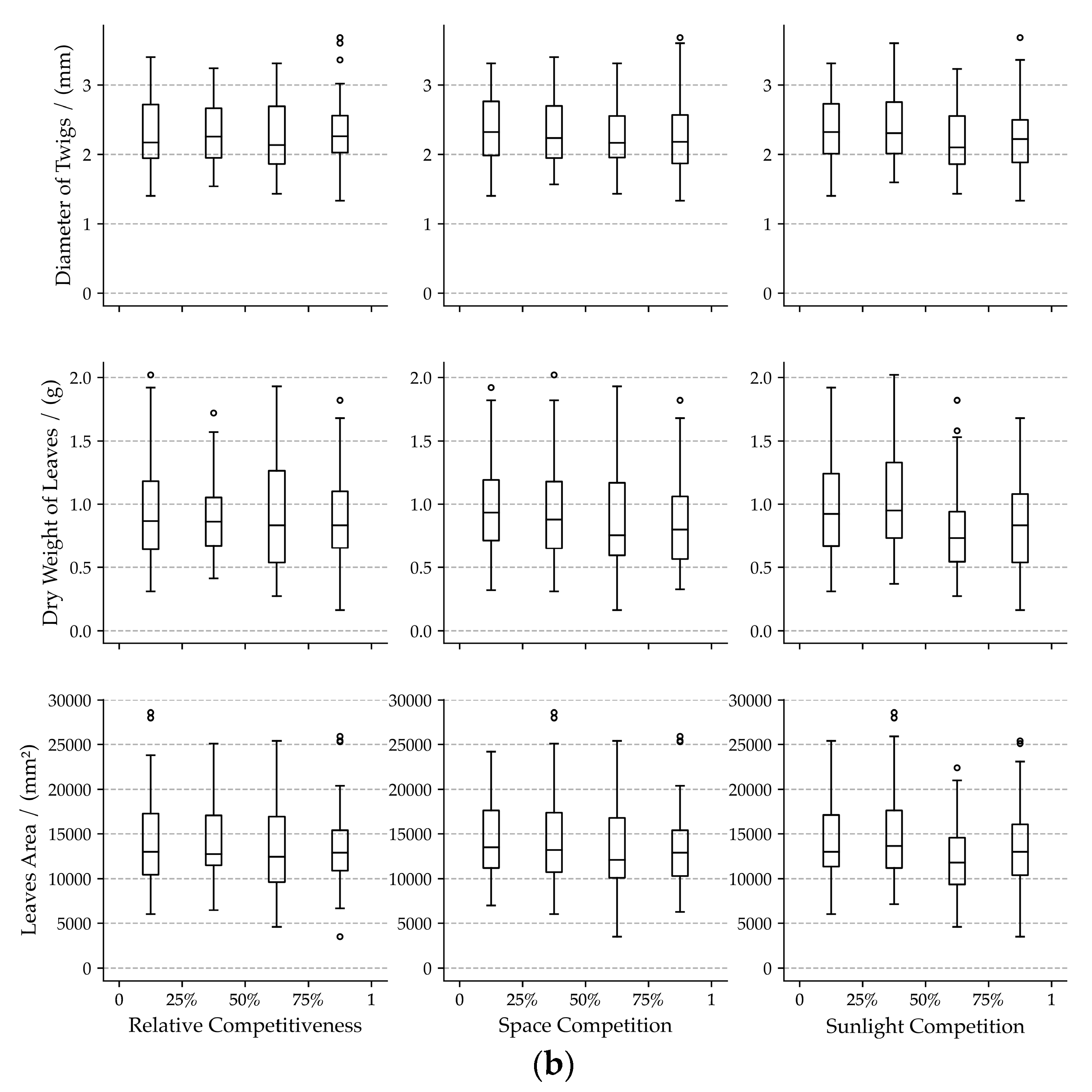
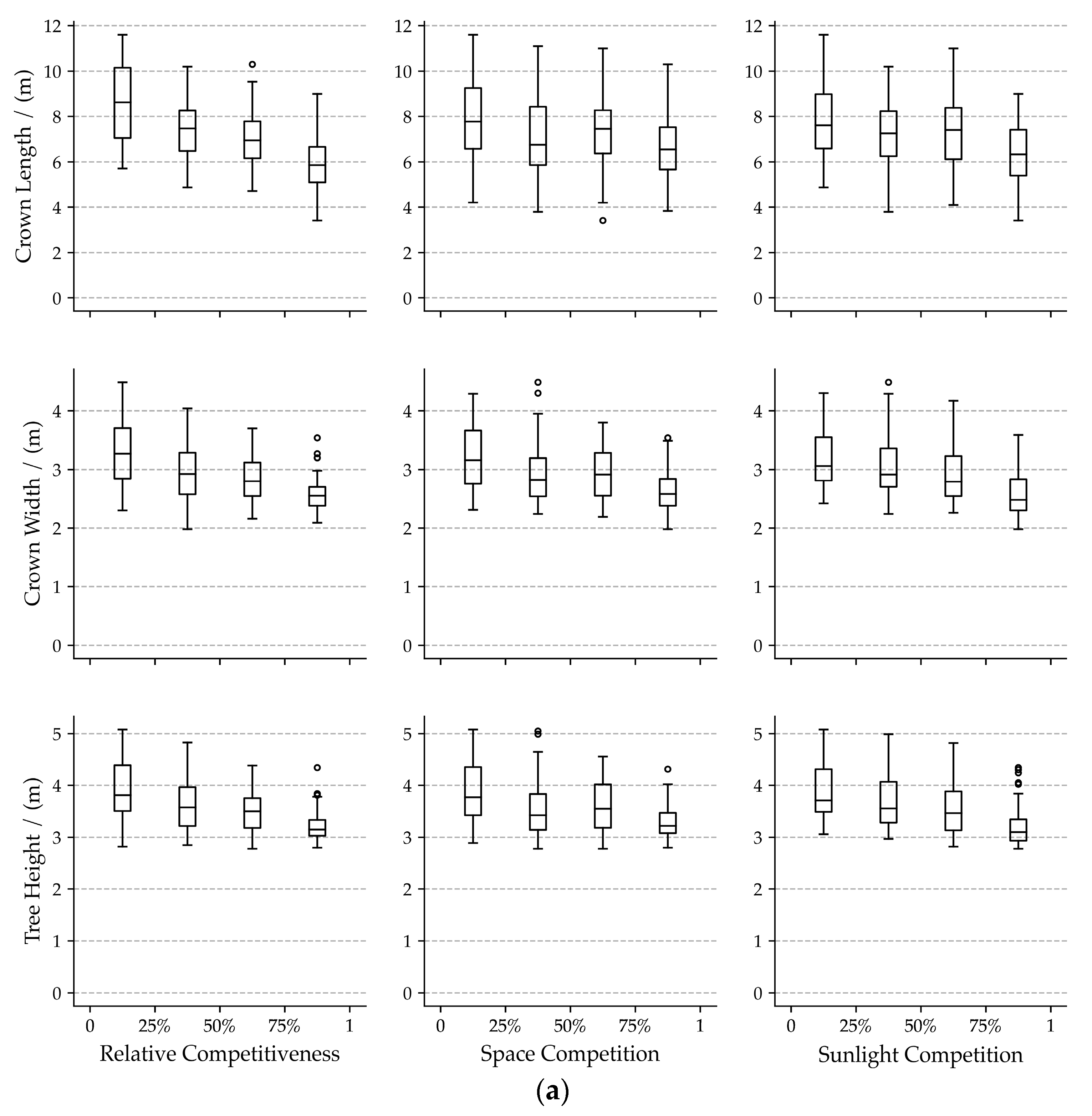
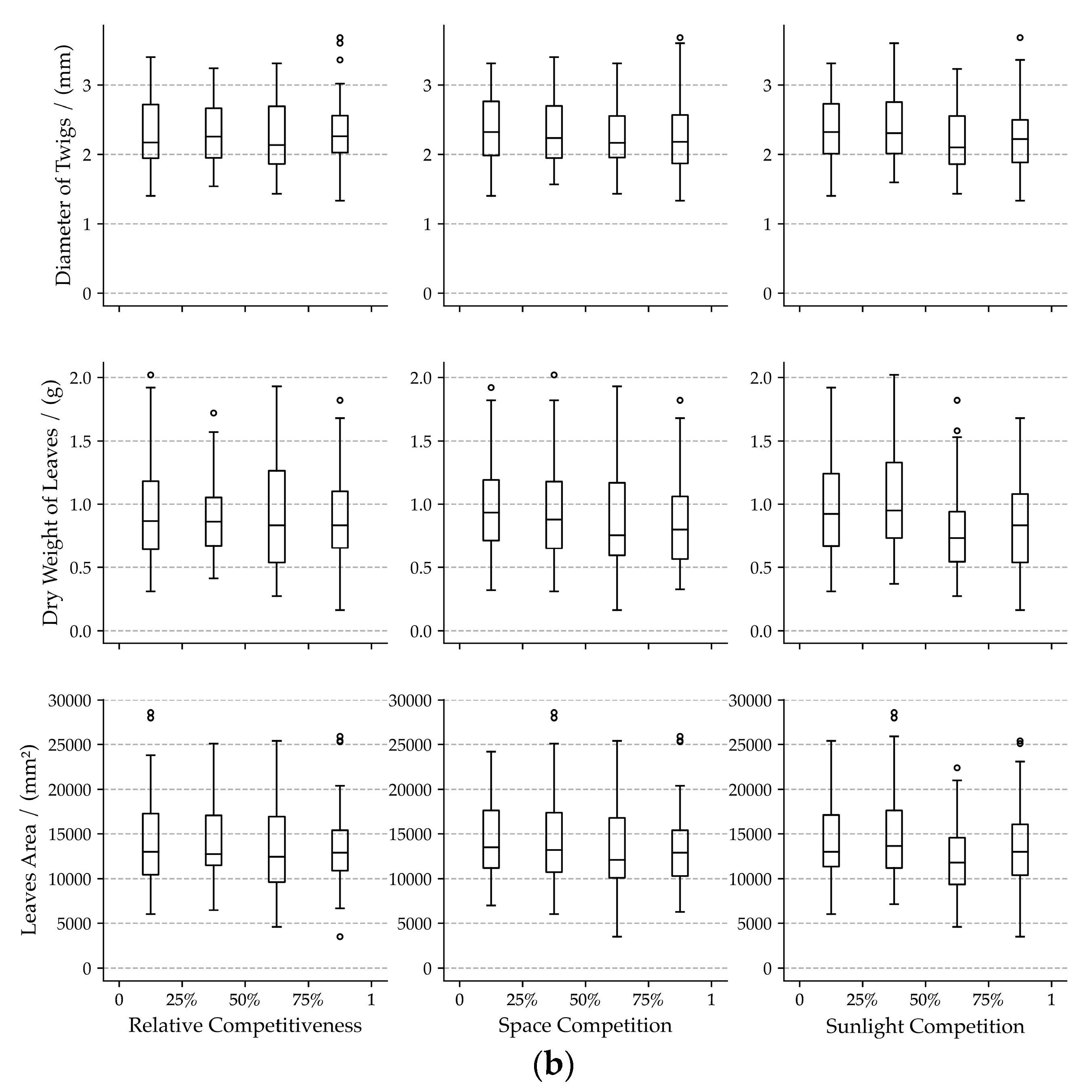
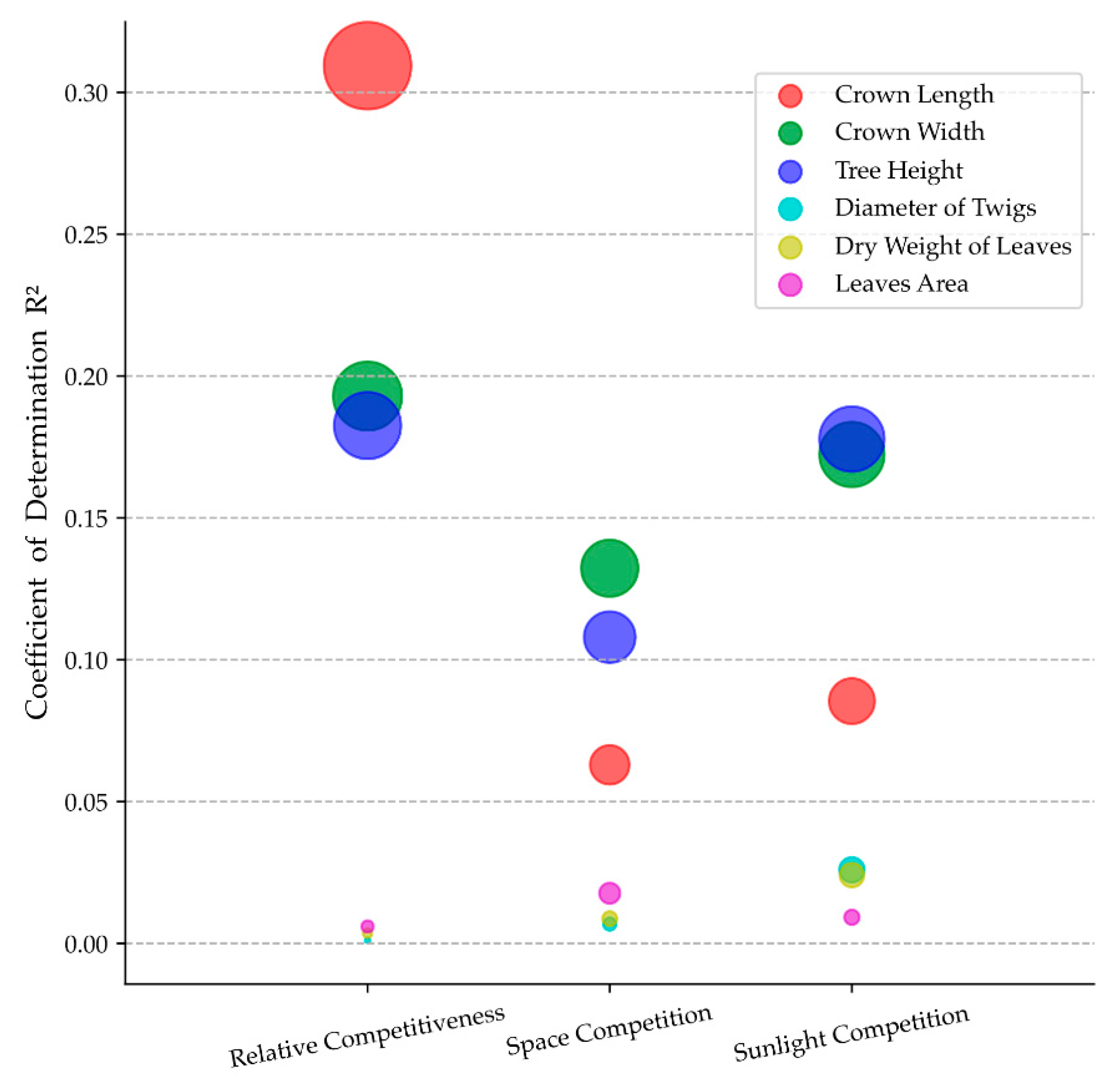
3.1. Bivariate Relationships among Visual Forms Traits and Competition Indicators
3.2. Confirmatory Factor Analysis of SEM
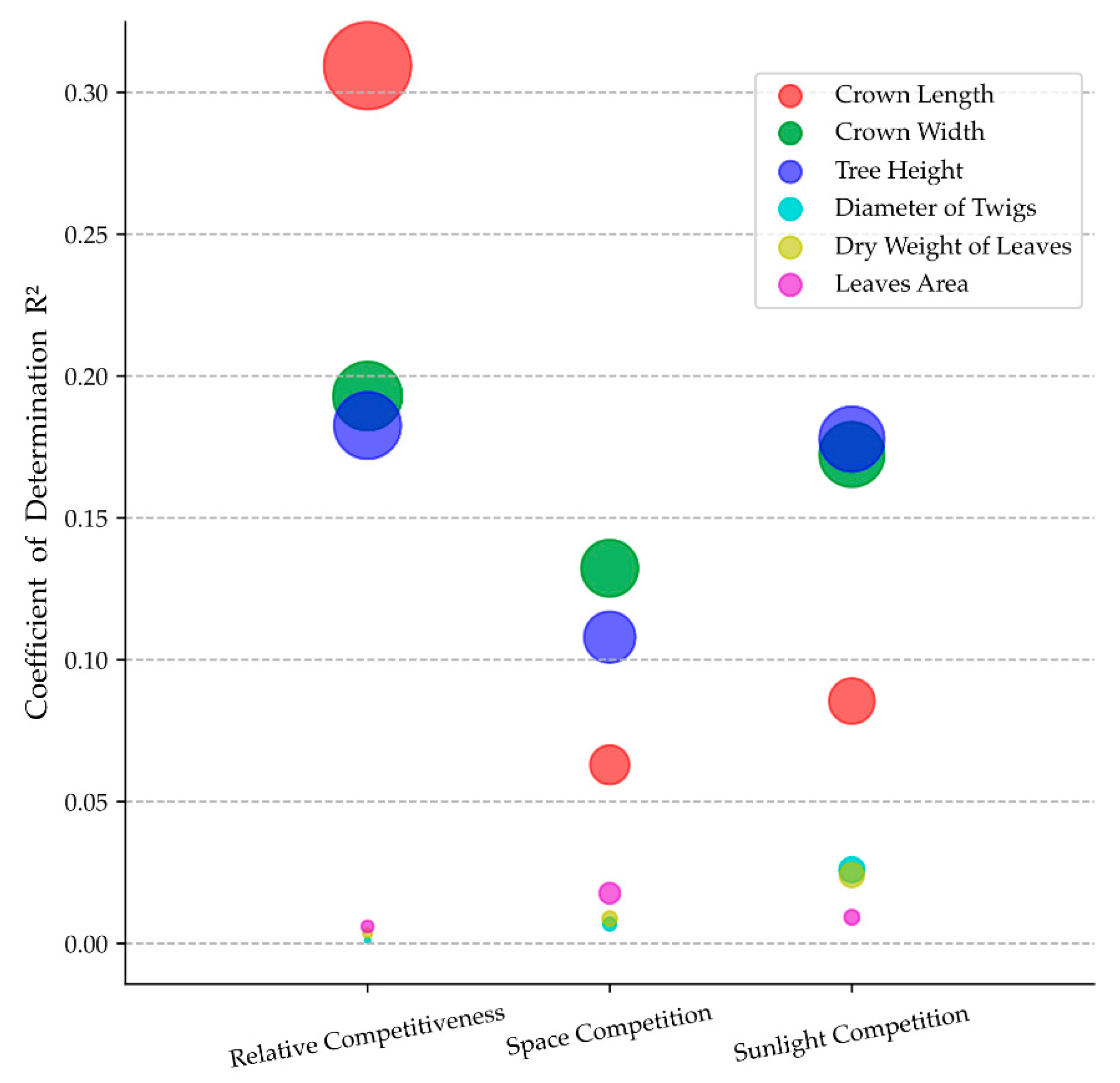
3.3. Effects of the Structural Model in SEM
4. Discussion
4.1. Effects of Neighboring Competition on Visual Forms
4.2. Application of SEM
4.3. Suggestions on Peri-Urban Forest Management
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haase, D.; Larondelle, N.; Andersson, E.; Artmann, M.; Borgström, S.; Breuste, J.; Gomez-Baggethun, E.; Gren, Å.; Hamstead, Z.; Hansen, R.; et al. A quantitative review of urban ecosystem service assessments: Concepts, models, and implementation. Ambio 2014, 43, 413–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirpke, U.; Meisch, C.; Marsoner, T.; Tappeiner, U. Revealing spatial and temporal patterns of outdoor recreation in the European Alps and their surroundings. Ecosyst. Serv. 2018, 31, 336–350. [Google Scholar] [CrossRef]
- Hartig, T.; Evans, G.W.; Jamner, L.D.; Davis, D.S.; Gärling, T. Tracking restoration in natural and urban field settings. J. Environ. Psychol. 2003, 23, 109–123. [Google Scholar] [CrossRef]
- Deng, Z.J.; Cheng, H.Y.; Song, S.Q. Effects of temperature, scarification, dry storage, stratification, phytohormone and light on dormancy-breaking and germination of Cotinus coggygria var. cinerea (Anacardiaceae) seeds. Seed Sci. Technol. 2010, 38, 572–584. [Google Scholar] [CrossRef]
- Zhang, Q. Basic Design the Dynamics of Visual Form from the First Generation Modernism Constructs the Master Reading the Building Constitute Beautiful. Master’s Thesis, Tianjin University, Tianjin, China, 2006. [Google Scholar]
- Dong, W.X. Visual Form Semantics; Shanghai University Press: Shanghai, China, 2007. [Google Scholar]
- Li, W. Study on Skin Renewal Design of the Old Industrial Buildings. Master’s Thesis, Qingdao University of Technological, Qingdao, China, 2012. [Google Scholar]
- Rowe, N.; Speck, T. Plant growth forms: An ecological and evolutionary perspective. New Phytol. 2005, 166, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Umeki, K. A comparison of crown asymmetry between Picea abies and Betula maximowicziana. Can. J. For. Res. 1995, 25, 1876–1880. [Google Scholar] [CrossRef]
- Getzin, S.; Wiegand, K. Asymmetric tree growth at the stand level: Random crown patterns and the response to slope. For. Ecol. Manag. 2007, 242, 165–174. [Google Scholar] [CrossRef]
- Grams, T.E.E.; Andersen, C.P. Competition for resources in trees: Physiological versus morphological plasticity. Prog. Bot. 2007, 68, 356–381. [Google Scholar] [CrossRef]
- Wu, J.; Chen, Y.; Liu, H.X.; Xu, L.J.; Jin, G.X.; Xu, C.Y. Effects of stand density and mingling intensity on tree morphology in natural scenic forest in Changbai Mountain. Sci. Silvae Sin. 2018, 54, 12–21. [Google Scholar] [CrossRef]
- Kang, H.J.; Chen, Z.L.; Liu, P.; Zhang, Z.X.; Zhou, J.H. Intra-specific competition of Emmenopterys henryi and its accompanying species in the Dapanshan national nature reserve of Zhejiang province. Acta Ecol. Sin. 2008, 28, 3456–3463. [Google Scholar] [CrossRef]
- Oheimb, G.V.; Lang, A.C.; Bruelheide, H.; Forrester, D.I.; Wäsche, I.; Yu, M.J.; Härdtle, W. Individual-tree radial growth in a subtropical broad-leaved forest: The role of local neighbourhood competition. For. Ecol. Manag. 2011, 261, 499–507. [Google Scholar] [CrossRef]
- Yin, D.S.; Ge, W.Z.; Zhang, F.H.; Shen, H.L. Competition relationship of populations of natural secondary Acer mono forest. Bull. Bot. Res. 2012, 32, 105–109. [Google Scholar]
- Yu, L.; Song, M.Y.; Lei, Y.B.; Korpelainen, H.; Niinemets, Ü.; Li, C.Y. Effects of competition and phosphorus fertilization on leaf and root traits of late-successional conifers Abies fabri and Picea brachytyla. Environ. Exp. Bot. 2019, 162, 14–24. [Google Scholar] [CrossRef] [Green Version]
- Grace, J.B.; Youngblood, A.; Scheiner, S.M. Structural equation modeling and ecological experiments. In Real World Ecology: Large-Scale and Long-Term Case Studies and Methods; Miao, S.L., Carstenn, S., Nungesser, M., Eds.; Springer: New York, NY, USA, 2009; pp. 19–46. [Google Scholar]
- Shipley, B. Cause and Correlation in Biology: A User’s Guide to Path Analysis, Structural Equations and Causal Inference with R.; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar]
- Grace, J.B. Structural equation modeling for observational studies. J. Wildl. Manag. 2008, 72, 14–22. [Google Scholar] [CrossRef]
- Timilsina, N.; Escobedo, F.J.; Staudhammer, C.L.; Brandeis, T. Analyzing the causal factors of carbon stores in a subtropical urban forest. Ecol. Complex. 2014, 20, 23–32. [Google Scholar] [CrossRef]
- Cottam, G.; Curtis, J.T. The use of distance measures in phytosociological sampling. Ecology 1956, 37, 451–460. [Google Scholar] [CrossRef]
- Zhang, J.E. Experimental Research Methods and Techniques Are Commonly Used in Ecology; Chemical Industry Press: Beijing, China, 2007; pp. 86–87. [Google Scholar]
- Schumacker, R.E.; Lomax, R.G. A Beginner’s Guide to Structural Equation Modeling; Routledge: New York, NY, USA, 2010. [Google Scholar]
- Lam, T.Y.; Maguire, D.A. Structural equation modeling: Theory and applications in forest management. Int. J. Forestry Res. 2012, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kline, R.B. Principles and Practice of Structural Equation Modeling, 3rd ed.; The Guilford Press: New York, NY, USA, 2011. [Google Scholar]
- Roxo, J. Elements of Art: Interpreting Meaning through the Language of Visual Cues. Doctoral Thesis, State University of New York at Stony Brook, New York, NY, USA, 2018. [Google Scholar]
- Iwasa, Y.; Cohen, D.; Leon, J.A. Tree height and crown shape, as results of competitive games. J. Theor. Biol. 1984, 112, 279–297. [Google Scholar] [CrossRef]
- Lindh, B.C.; Gray, A.N.; Spies, T.A. Responses of herbs and shrubs to reduced root competition under canopies and in gaps: A trenching experiment in old-growth Douglas-fir forests. Can. J. Forest Res. 2003, 33, 2052–2057. [Google Scholar] [CrossRef] [Green Version]
- Riegel, G.M.; Miller, R.F.; Krueger, W.C. Competition for resources between understory vegetation and overstory Pinus ponderosa in northeastern Oregon. Ecol. Appl. 1992, 2, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Olivier, M.D.; Robert, S.; Fournier, R.A. Response of sugar maple (Acer saccharum, Marsh.) tree crown structure to competition in pure versus mixed stands. For. Ecol. Manag. 2016, 374, 20–32. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Messier, C.; Stadt, K.J.; Gendron, F.; Comeau, P.G. Predicting and managing light in the understory of boreal forests. Can. J. For. Res. 1999, 29, 796–811. [Google Scholar] [CrossRef]
- Bella, I.E. A new competition model for individual trees. For. Sci. 1971, 17, 364–372. [Google Scholar]
- Opie, J.E. Predictability of individual tree growth using various definitions of competing basal area. For. Sci. 1968, 14, 314–323. [Google Scholar] [CrossRef]
- Smith, W.R.; Farrar, R.M., Jr.; Murphy, P.A.; Yeiser, J.L.; Meldahl, R.S.; Kush, J.S. Crown and basal area relationships of open-grown southern pines for modeling competition and growth. Can. J. For. Res. 1992, 22, 341–347. [Google Scholar] [CrossRef]
- Van der Zee, J.; Lau, A.; Shenkin, A. Understanding crown shyness from a 3-D perspective. Ann. Bot. 2021, 3, 1–11. [Google Scholar] [CrossRef]
- Onoda, Y.; Bando, R. Wider crown shyness between broad-leaved tree species than between coniferous tree species in a mixed forest of Castanopsis cuspidata and Chamaecyparis obtusa. Ecol. Res. 2021, 36, 733–743. [Google Scholar] [CrossRef]
- Smith, S.H.; Bell, J.F. Using competitive stress index to estimate diameter growth for thinned Douglas-fir stands. For. Sci. 1983, 29, 491–499. [Google Scholar] [CrossRef]
- Baskerville, G.L. Dry-matter production in immature balsam fir stands: Roots lesser vegetation, and total stand. For. Sci. 1966, 12, 49–53. [Google Scholar] [CrossRef]
- Weiner, J.; Damgaard, C. Size-asymmetric competition and size-asymmetric growth in a spatially explicit zone-of-influence model of plant competition. Ecol. Res. 2006, 21, 707–712. [Google Scholar] [CrossRef]
- Horn, H.S. The Adaptive Geometry of Trees; Princeton University Press: Princeton, NJ, USA, 1971. [Google Scholar]
- Westoby, M.; Wright, I.J. The leaf size-twig size spectrum and its relationship to other important spectra of variation among species. Oecologia 2003, 135, 621–628. [Google Scholar] [CrossRef]
- Niklas, K.J. Size-dependent allometry of tree height, diameter and trunk-taper. Ann. Bot. 1995, 75, 217–227. [Google Scholar] [CrossRef]
- Tyree, M.T.; Ewers, F.W. The hydraulic architecture of trees and other woody plants. New Phytol. 1991, 119, 345–360. [Google Scholar] [CrossRef]
- Weiner, J.; Berntson, G.M.; Thomas, S.C. Competition and growth form in a woodland annual. J. Ecol. 1990, 78, 459–469. [Google Scholar] [CrossRef]
- Dell’Olio, L. Structural equation models. In Public Transportation Quality of Service; Dell’Olio, L., Ibeas, A., de Oña, J., de Oña, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 141–152. [Google Scholar] [CrossRef] [Green Version]
- Browne, M.W.; Cudeck, R. Alternative ways of assessing model fit. Sociol. Method. Res. 1992, 21, 230–258. [Google Scholar] [CrossRef]
- Gerbing, D.W.; Anderson, J.C. Monte Carlo evaluations of goodness of fit indices for structural equation models. Sociol. Method. Res. 1992, 21, 132–160. [Google Scholar] [CrossRef]
- González de Andrés, E.; Camarero, J.J.; Blanco, J.A.; Imbert, J.B.; Lo, Y.H.; Sangüesa-Barreda, G.; Castillo, F.J. Tree-to-tree competition in mixed European beech-Scots pine forests has different impacts on growth and water-use efficiency depending on site conditions. J. Ecol. 2018, 106, 59–75. [Google Scholar] [CrossRef] [Green Version]
- Kahriman, A.; Şahin, A.; Sönmez, T.; Yavuz, M. A novel approach to selecting a competition index: The effect of competition on individual-tree diameter growth of Calabrian pine. Can. J. For. Res. 2018, 48, 1217–1226. [Google Scholar] [CrossRef]
- Kramer, R.D.; Sillett, S.C.; Van Pelt, R.; Franklin, J.F. Neighborhood competition mediates crown development of Picea sitchensis in Olympic rainforests: Implications for restoration management. For. Ecol. Manag. 2019, 441, 127–143. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- Lee, T.D.; Bazzaz, F.A. Effects of defoliation and competition on growth and reproduction in the annual plant Abutilon theophrasti. J. Ecol. 1980, 68, 813–821. [Google Scholar] [CrossRef]
- Mekonnen, Z.A.; Riley, W.J.; Grant, R.F. Accelerated nutrient cycling and increased light competition will lead to 21st century shrub expansion in North American Arctic tundra. J. Geophys. Res. Biogeo. 2018, 123, 1683–1701. [Google Scholar] [CrossRef]
- Longstreth, D.J.; Nobel, P.S. Nutrient influences on leaf photosynthesis: Effects of nitrogen, phosphorus, and potassium for Gossypium hirsutum L. Plant Physiol. 1980, 65, 541–543. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B.; Ellsworth, D.S.; Walters, M.B. Leaf structure (specific leaf area) modulates photosynthesis-nitrogen relations: Evidence from within and across species and functional groups. Funct. Ecol. 1998, 12, 948–958. [Google Scholar] [CrossRef]
- Long, J.Y.; Zhao, Y.M.; Kong, X.Q.; Chen, Z.Y.; Wang, X.S.; Zhao, K.; Cao, R.; Huang, L.S.; Lü, J.; Cui, Y.; et al. Trade-offs between twig and leaf traits of ornamental shrubs grown in shade. Acta Ecol. Sin. 2018, 38, 1–9. [Google Scholar] [CrossRef]
- Kocher, S.D.; Harris, R. Forest Stewardship Series 5: Tree Growth and Competition; University of California Agriculture and Natural Resources: California, CA, USA, 2007; pp. 1–10. [Google Scholar]
- Coomes, D.A.; Allen, R.B. Testing the metabolic scaling theory of tree growth. J. Ecol. 2009, 97, 1369–1373. [Google Scholar] [CrossRef]
- Lai, Y.B. On the phenomenon of “aesthetic fatigue” in the perspective of consumer society. Mov. Lit. 2011, 21, 17–18. [Google Scholar]
- Feng, X.L. Aesthetics in the System of Human Life; Anhui Education Press: Hefei, China, 2004. [Google Scholar]
- Xue, X. A Study on Aesthetic Fatigue of Forest Landscapes of Tourists Based on EEG. Master’s Thesis, Central South University of Forestry and Technology, Changsha, China, 2015. [Google Scholar]
- Hull, R.B.; Buhyoff, G.J. The scenic beauty temporal distribution method: An attempt to make scenic beauty assessments compatible with forest planning efforts. For. Sci. 1986, 32, 271–286. [Google Scholar]
- Bell, S.; Blom, D.; Rautamäki, M.; Castel-Branco, C.; Simson, A.; Olsen, I.A. Design of urban forests. In Urban Forests and Trees: A Reference Book; Konijnendijk, C.C., Nilsson, K., Randrup, T.B., Schipperijn, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 149–186. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Abbr. | Accuracy | Traits | Abbr. | Accuracy |
|---|---|---|---|---|---|
| Base diameter of current year twig | BD | 0.02 mm | Height to the lowest live branch | HL | 0.01 m |
| Crown width | CW | 0.1 m | Leaf area | LA | 0.1 mm2 |
| Distance | dist | 0.01 m | Stem collar diameter | CD | 0.1 cm |
| Dry weight of leaves | DW | 0.01 g | Tree height | H | 0.1 m |
| Fresh weight of leaves | FW | 0.01 g |
| Construct | Traits | Unit | Min. | Max. | Average | Standard Deviation |
|---|---|---|---|---|---|---|
| Crown Forms | Crown Length | m | 1.08 | 4.49 | 2.94 | 0.87 |
| Crown Width | m | 1.07 | 6.75 | 3.62 | 1.17 | |
| Tree Height | m | 1.68 | 6.41 | 3.58 | 0.91 | |
| Foliage Forms | Diameter of Current Year Twigs | mm | 1.33 | 3.68 | 2.31 | 0.48 |
| Dry Weight of Leaves | g | 0.03 | 0.64 | 0.19 | 0.13 | |
| Leaves Area | mm2 | 3501 | 28,588 | 13,882 | 4810 |
| Neighboring Competition Indicator | Min. | Max. | Average | Standard Deviation |
| Relative Competitiveness | 0.02 | 2.68 | 0.55 | 0.48 |
| Space Competition | 0.00 | 2.46 | 0.46 | 0.47 |
| Sunlight Competition | 1.44 | 35.36 | 7.79 | 5.39 |
| Construct | Item | Unstandardized Factor Loading | Cronbach’s Alpha | Construct Reliability (CR) | Average Variance Extracted (AVE) |
|---|---|---|---|---|---|
| Neighboring Competition | Relative Competitiveness | 1.0 | 0.71 | 0.80 | 0.51 |
| Space Competition | 0.88 | ||||
| Sunlight Competition | 0.66 | ||||
| Crown Forms | Crown Length | 0.65 | 0.82 | 0.86 | 0.68 |
| Crown Width | 0.87 | ||||
| Tree Height | 1.0 | ||||
| Foliage Forms | Diameter of Current Year Twigs | 0.82 | 0.76 | 0.81 | 0.51 |
| Dry Weight of Leaves | 1.0 | ||||
| Leaves Area | 0.83 |
| Index | Value | Recommended Value |
|---|---|---|
| χ2/df | 1.68 | <5.00 |
| Comparative Fit Index (CFI) | 0.99 | >0.90 |
| Goodness of Fit Index (GFI) | 0.96 | >0.90 |
| Root Mean Square Error of Approximation (RMSEA) | 0.06 | <0.08 |
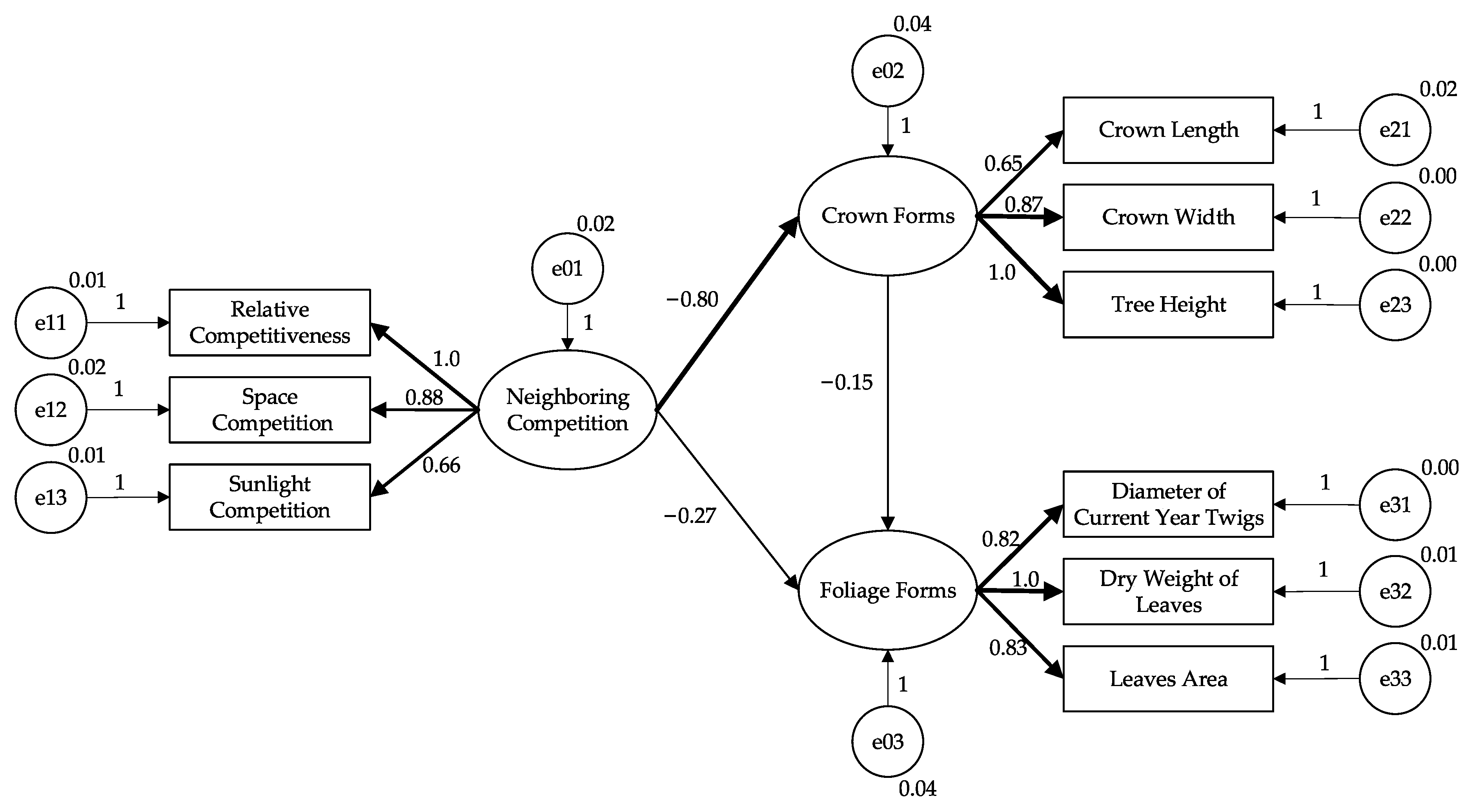
| Paths in SEM | Direct Effect | Indirect Effect | Total Effect 1 | p-Value 2 | |
|---|---|---|---|---|---|
| Neighboring Competition → | Crown Forms | −0.80 | -- 3 | −0.80 | 0.000 |
| Foliage Forms | −0.27 | 0.12 | −0.15 | -- 3 | |
| Crown Forms → | Foliage Forms | −0.15 | -- 3 | −0.15 | 0.006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Zhu, J.; Xu, C.; Hauer, R.J. Responses of Visual Forms to Neighboring Competition: A Structural Equation Model for Cotinus coggygria var. cinerea Engl. Forests 2021, 12, 1073. https://doi.org/10.3390/f12081073
Cao Y, Zhu J, Xu C, Hauer RJ. Responses of Visual Forms to Neighboring Competition: A Structural Equation Model for Cotinus coggygria var. cinerea Engl. Forests. 2021; 12(8):1073. https://doi.org/10.3390/f12081073
Chicago/Turabian StyleCao, Yujuan, Jiyou Zhu, Chengyang Xu, and Richard J. Hauer. 2021. "Responses of Visual Forms to Neighboring Competition: A Structural Equation Model for Cotinus coggygria var. cinerea Engl." Forests 12, no. 8: 1073. https://doi.org/10.3390/f12081073
APA StyleCao, Y., Zhu, J., Xu, C., & Hauer, R. J. (2021). Responses of Visual Forms to Neighboring Competition: A Structural Equation Model for Cotinus coggygria var. cinerea Engl. Forests, 12(8), 1073. https://doi.org/10.3390/f12081073