
Climate-Driven Differences in Growth Performance of Cohabitant Fir and Birch in a Subalpine Forest in Dhorpatan Nepal

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Tree Species
2.3. Climate Data
2.4. Tree-Ring Data
2.5. Tree Growth-Climate Correlations
3. Results
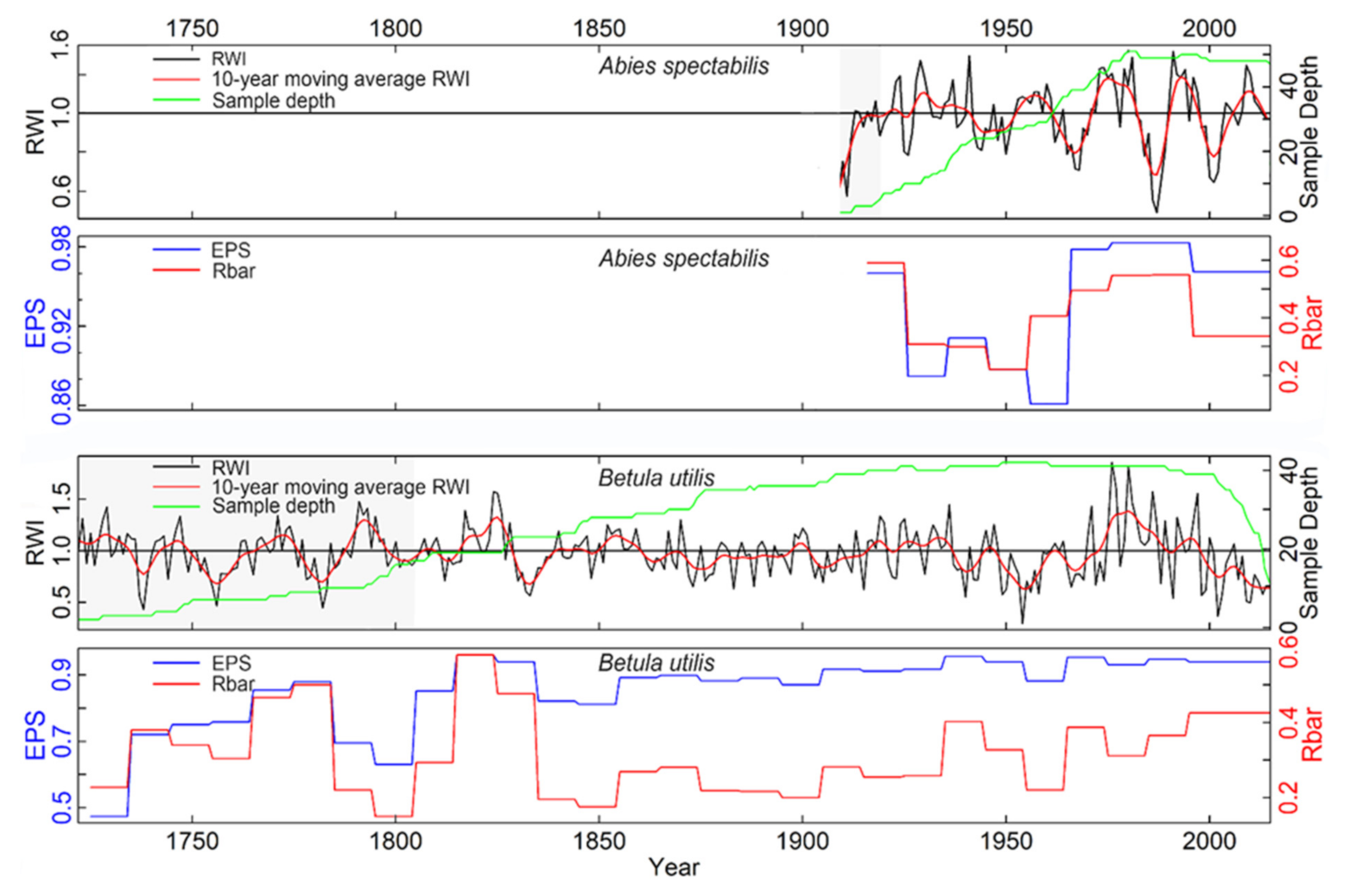
3.1. Tree-Ring Chronology and Radial Growth Trend
3.2. Growth Response to Climate Variation
3.2.1. Static Correlation
3.2.2. Moving Correlation
4. Discussion
4.1. Tree-Ring Chronology and Growth Trend
4.2. Growth-Climate Relationships
4.3. Shifts in Growth-Climate Sensitivity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Climate | Temperature (°C) | Precipitation (mm) |
|---|---|---|
| Annual | 0.52 * | −61.02 |
| January | 0.48 * | −1.97 |
| February | 0.87 * | −1.74 |
| March | 0.72 * | 9.23 |
| April | 0.43 | 0.52 |
| May | 0.02 | 28.50 * |
| June | 0.17 | −27.95 |
| July | 0.19 | −16.39 |
| August | 0.34 | −53.11 * |
| September | 0.36 * | 6.96 |
| October | 0.59 * | −8.95 |
| November | 1.06 * | −0.37 |
| December | 1.02 * | 4.26 |
Appendix B

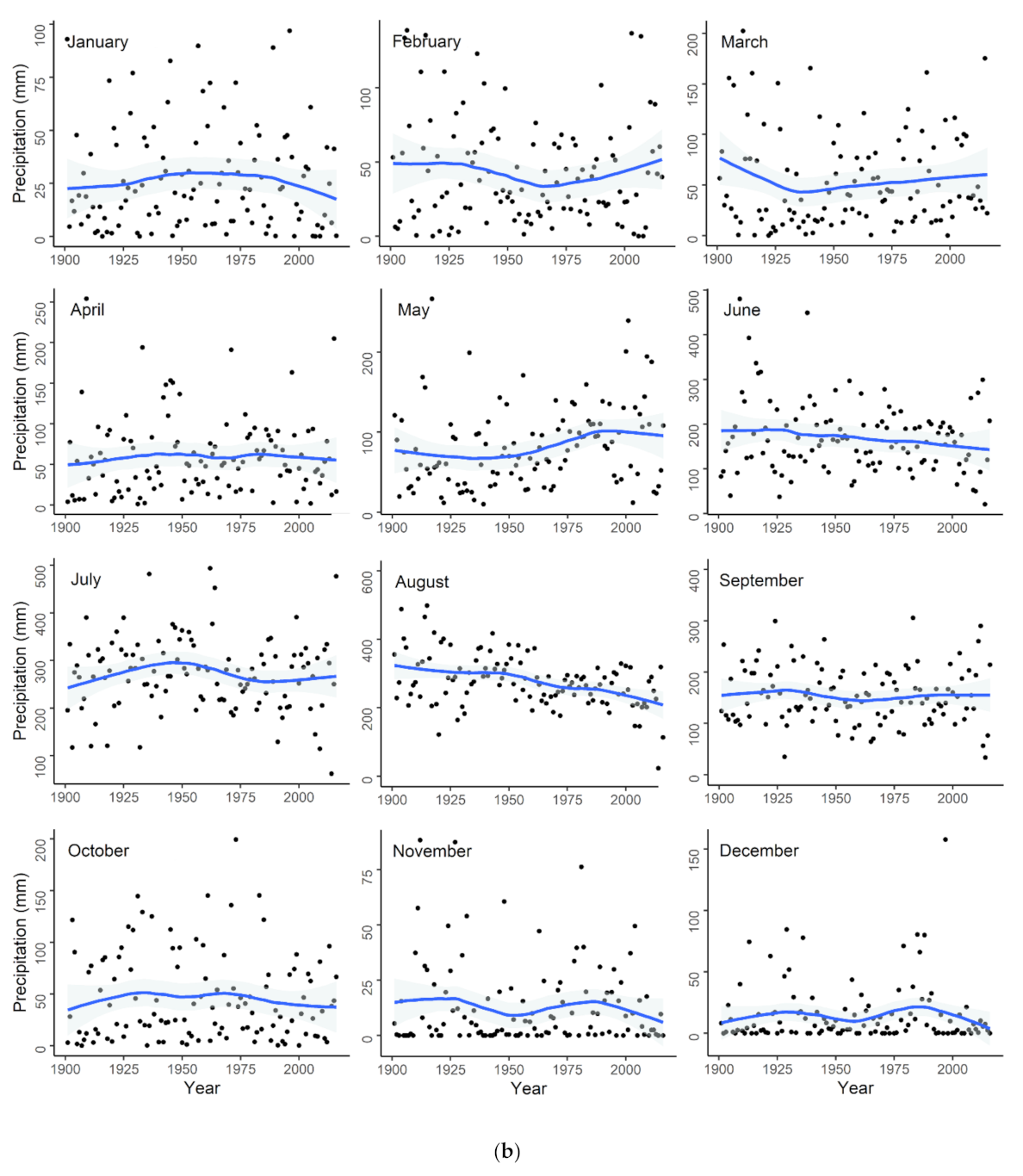
Appendix C
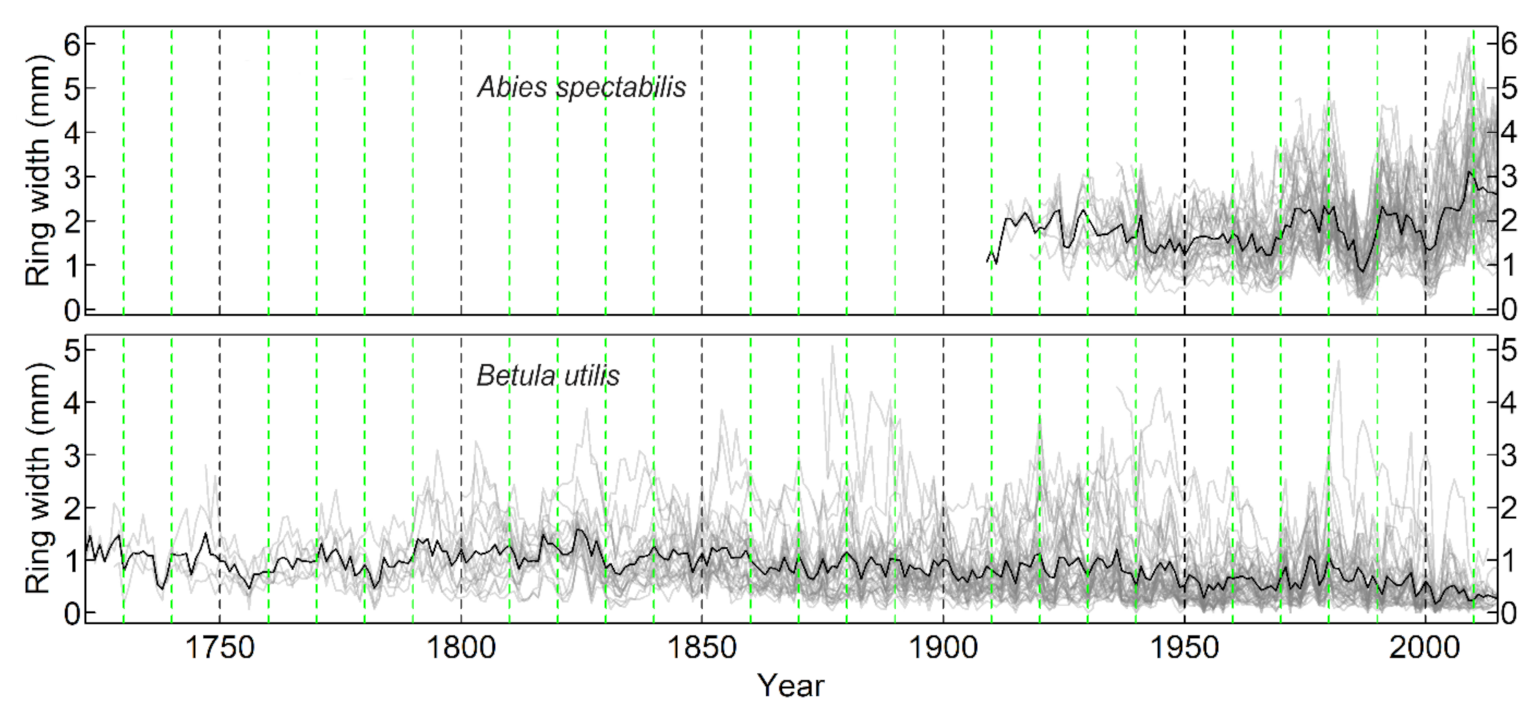
Appendix D
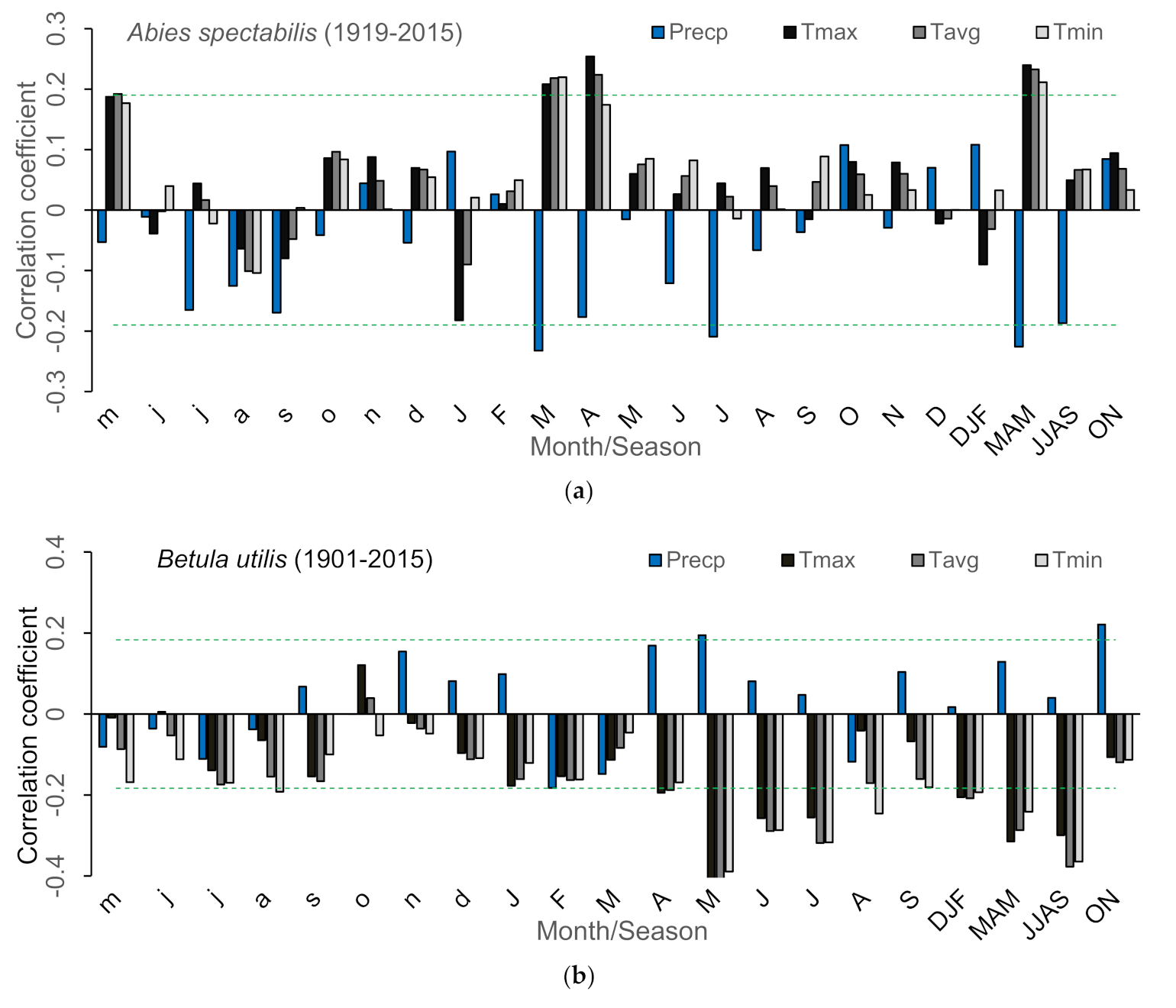
Appendix E

References
- Götzenberger, L.; de Bello, F.; Bråthen, K.A.; Davison, J.; Dubuis, A.; Guisan, A.; Lepš, J.; Lindborg, R.; Moora, M.; Pärtel, M.; et al. Ecological Assembly rules in plant communities—Approaches, patterns and prospects. Biol. Rev. 2012, 87, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Bastias, C.C.; Godoy, O.; Granda, E.; Escudero, A. Species coexistence in a changing world. Front. Plant Sci. 2015, 6, 866. [Google Scholar] [CrossRef]
- Sperry, J.S.; Hacke, U.G.; Pittermann, J. Size and function in conifer tracheids and angiosperm vessels. Am. J. Bot. 2006, 93, 1490–1500. [Google Scholar] [CrossRef] [Green Version]
- Stewart, G.H. Population dynamics of a montane conifer forest, western Cascade Range, Oregon, USA. Ecology 1986, 67, 534–544. [Google Scholar] [CrossRef]
- Bond, W.J. The tortoise and the hare: Ecology of angiosperm dominance and gymnosperm persistence. Biol. J. Linn. Soc. 1989, 36, 227–249. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems, 2nd ed.; Springer: New York, NY, USA, 2003. [Google Scholar]
- Rayback, S.A.; Shrestha, K.B.; Hofgaard, A. growth variable-specific moisture and temperature limitations in co-occurring alpine tree and shrub species, central Himalayas, Nepal. Dendrochronologia 2017, 44, 193–202. [Google Scholar] [CrossRef]
- Schickhoff, U.; Bobrowski, M.; Böhner, J.; Bürzle, B.; Chaudhary, R.P.; Gerlitz, L.; Heyken, H.; Lange, J.; Müller, M.; Scholten, T.; et al. Do Himalayan treelines respond to recent climate change? An evaluation of sensitivity indicators. Earth Syst. Dyn. 2015, 6, 245–265. [Google Scholar] [CrossRef]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef]
- Gaire, N.P.; Koirala, M.; Bhuju, D.R.; Carrer, M. Site- and species-specific treeline responses to climatic variability in eastern Nepal Himalaya. Dendrochronologia 2017, 41, 44–56. [Google Scholar] [CrossRef]
- Pandey, S.; Cherubini, P.; Saurer, M.; Carrer, M.; Petit, G. Effects of climate change on treeline trees in Sagarmatha (Mt. Everest, Central Himalaya). J. Veg. Sci. 2020, 31, 1144–1153. [Google Scholar] [CrossRef]
- Schwab, N.; Kaczka, R.J.; Janecka, K.; Böhner, J.; Chaudhary, R.P.; Scholten, T.; Schickhoff, U. Climate change-induced shift of tree growth sensitivity at a central Himalayan treeline ecotone. Forests 2018, 9, 267. [Google Scholar] [CrossRef] [Green Version]
- Gaire, N.P.; Koirala, M.; Bhuju, D.R.; Borgaonkar, H.P. Treeline dynamics with climate change at the central Nepal Himalaya. Clim. Past 2014, 10, 1277–1290. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, A.; Fan, Z.-X.; Jump, A.S.; Zhou, Z.-K. Warming induced growth decline of Himalayan birch at its lower range edge in a semi-arid region of trans-Himalaya, central Nepal. Plant Ecol. 2017, 218, 621–633. [Google Scholar] [CrossRef]
- Chhetri, P.K.; Cairns, D.M. Contemporary and historic population structure of Abies spectabilis at treeline in Barun Valley, eastern Nepal Himalaya. J. Mt. Sci. 2015, 12, 558–570. [Google Scholar] [CrossRef]
- Chhetri, P.K.; Cairns, D.M. Dendroclimatic response of Abies spectabilis at treeline ecotone of Barun Valley, eastern Nepal Himalaya. J. For. Res. 2016, 27, 1163–1170. [Google Scholar] [CrossRef]
- Lv, L.-X.; Zhang, Q.-B. Asynchronous recruitment history of Abies spectabilis along an altitudinal gradient in the Mt. Everest region. J. Plant Ecol. 2012, 5, 147–156. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, B.B.; Ghimire, B.; Lekhak, H.D.; Jha, P.K. Regeneration of treeline birch (Betula utilis D. Don) forest in a trans-Himalayan dry valley in Central Nepal. Mt. Res. Dev. 2007, 27, 259–267. [Google Scholar] [CrossRef]
- Tiwari, A.; Fan, Z.-X.; Jump, A.S.; Li, S.-F.; Zhou, Z.-K. Gradual expansion of moisture sensitive Abies spectabilis forest in the trans-Himalayan zone of central Nepal associated with climate change. Dendrochronologia 2017, 41, 34–43. [Google Scholar] [CrossRef]
- Tiwari, A.; Jha, P.K. An overview of treeline response to environmental changes in Nepal Himalaya. Trop. Ecol. 2018, 59, 273–285. [Google Scholar]
- Shrestha, K.B.; Hofgaard, A.; Vandvik, V. Recent treeline dynamics are similar between dry and mesic areas of Nepal, central Himalaya. J. Plant Ecol. 2015, 8, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Chhetri, P.K.; Bista, R.; Shrestha, K.B. How Does the stand structure of treeline-forming species shape the treeline ecotone in different regions of the Nepal Himalayas? J. Mt. Sci. 2020, 17, 2354–2368. [Google Scholar] [CrossRef]
- Takahashi, K.; Tokumitsu, Y.; Yasue, K. Climatic factors affecting the tree-ring width of Betula ermanii at the timberline on Mount Norikura, Central Japan. Ecol. Res. 2005, 20, 445–451. [Google Scholar] [CrossRef] [Green Version]
- Miehe, G.; Miehe, S.; Böhner, J.; Bäumler, R.; Ghimire, S.K.; Bhattarai, K.; Chaudhary, R.P.; Subedi, M.; Jha, P.K.; Pendry, C. Vegetation ecology. In Nepal. An Introduction to the Natural History, Ecology and Human Environment of the Himalayas. A Companion to the Flora of Nepal; Miehe, G., Pendry, C., Chaudhary, R.P., Eds.; Royal Botanic Garden: Edinburgh, UK, 2015; pp. 144–168. [Google Scholar]
- Carnicer, J.; Barbeta, A.; Sperlich, D.; Coll, M.; Peñuelas, J. Contrasting trait syndromes in angiosperms and conifers are associated with different responses of tree growth to temperature on a large scale. Front. Plant Sci. 2013, 4, 409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawadi, B.; Liang, E.; Tian, L.; Devkota, L.P.; Yao, T. Pre-monsoon precipitation signal in tree rings of timberline Betula utilis in the central Himalayas. Quat. Int. 2013, 283, 72–77. [Google Scholar] [CrossRef]
- Liang, E.; Dawadi, B.; Pederson, N.; Eckstein, D. Is the growth of birch at the upper timberline in the Himalayas limited by moisture or by temperature? Ecology 2014, 95, 2453–2465. [Google Scholar] [CrossRef] [Green Version]
- Chhetri, P.K.; Cairns, D.M. Low recruitment above treeline indicates treeline stability under changing climate in Dhorpatan Hunting Reserve, western Nepal. Phys. Geogr. 2018, 39, 329–342. [Google Scholar] [CrossRef]
- Department of National Parks and Wildlife Conservation (DNPWC); Dhorpatan Hunting Reserve. Available online: http://www.dnpwc.gov.np/en/conservation-area-detail/61/ (accessed on 14 December 2020).
- Bobrowski, M.; Gerlitz, L.; Schickhoff, U. Modelling the potential distribution of Betula utilis in the Himalaya. Glob. Ecol. Conserv. 2017, 11, 69–83. [Google Scholar] [CrossRef]
- University of East Anglia Climatic Research Unit; Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. CRU TS4.04: Climatic Research Unit (CRU) Time-Series (TS) Version 4.04 of high-resolution gridded data of month-by-month variation in climate (January 1901–December 2019). Cent. Environ. Data Anal. 2020. Available online: https://catalogue.ceda.ac.uk/uuid/89e1e34ec3554dc98594a5732622bce9 (accessed on 16 November 2020).
- Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 2020, 7, 109. [Google Scholar] [CrossRef] [Green Version]
- Guijarro, J.A. Climatol: Climate Tools (Series Homogenization and Derived Products). R Package Version 3.1.2. 2019. Available online: https://CRAN.R-project.org/package=climatol (accessed on 11 December 2020).
- Schweingruber, F.H.; Kairiukstis, L.; Shiyatov, S. Sample selection. In Methods of Dendrochronology; Cook, E.R., Kairiukstis, L.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; pp. 23–35. [Google Scholar]
- Phipps, R.L. Collecting, Preparing, Crossdating, and Measuring Tree Increment Cores; Department of the Interior, U.S. Geological Survey: Reston, VA, USA, 1985.
- Pilcher, J.R. Sample preparation, cross-dating, and measurement. In Methods of Dendrochronology; Cook, E.R., Kairiukstis, L.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; pp. 40–51. [Google Scholar]
- Speer, J.H. Fundamentals of Tree-Ring Research; University of Arizona Press: Tucson, AZ, USA, 2010. [Google Scholar]
- Bunn, A.G. A dendrochronology program library in R (DplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Version 4.0.3; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Cook, E.R.; Briffa, K.R.; Shiyatov, S.; Mazepa, V. Tree-ring standardization and growth-trend estimation. In Methods of Dendrochronology; Cook, E.R., Kairiukstis, L.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; pp. 104–123. [Google Scholar]
- Cook, E.R.; Shiyatov, S.; Mazepa, V. Estimation of the mean chronology. In Methods of Dendrochronology; Cook, E.R., Kairiukstis, L.A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; pp. 123–132. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- St. George, S. An Overview of Tree-Ring Width Records across the Northern Hemisphere. Quat. Sci. Rev. 2014, 95, 132–150. [Google Scholar] [CrossRef]
- Biondi, F.; Qeadan, F. A theory-driven approach to tree-ring standardization: Defining the biological trend from expected basal area increment. Tree-Ring Res. 2008, 64, 81–96. [Google Scholar] [CrossRef] [Green Version]
- Weiner, J.; Thomas, S.C. The nature of tree growth and the “age-related decline in forest productivity”. Oikos 2001, 94, 374–376. [Google Scholar] [CrossRef] [Green Version]
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; Mérian, P.; Qeadan, F.; Zang, C. DplR: Dendrochronology Program Library in R. R Package Version 1.7.1. 2020. Available online: https://CRAN.R-project.org/package=dplR (accessed on 16 November 2020).
- Phipps, R.L.; Field, M.L. Computer Programs to Calculate Basal Area Increment from Tree Rings; Department of the Interior, U.S. Geological Survey: Reston, VA, USA, 1989.
- Fritts, H.C. Tree Rings and Climate; Academic Press: New York, NY, USA, 1976. [Google Scholar]
- Zang, C.; Biondi, F. Treeclim: An R package for the numerical calibration of proxy-climate relationships (Version 2.0.5.1). Ecography 2014, 38, 431–436. [Google Scholar] [CrossRef]
- Pandey, S.; Carrer, M.; Castagneri, D.; Petit, G. Xylem anatomical responses to climate variability in Himalayan birch trees at one of the world’s highest forest limit. Perspect. Plant Ecol. Evol. Syst. 2018, 33, 34–41. [Google Scholar] [CrossRef]
- Kharal, D.K.; Thapa, U.K.; St. George, S.; Meilby, H.; Rayamajhi, S.; Bhuju, D.R. Tree-climate relations along an elevational transect in Manang Valley, central Nepal. Dendrochronologia 2017, 41, 57–64. [Google Scholar] [CrossRef]
- Lusk, C.H.; Jorgensen, M.A.; Bellingham, P.J. A conifer–angiosperm divergence in the growth vs. Shade tolerance trade-off underlies the dynamics of a New Zealand warm-temperate rain forest. J. Ecol. 2015, 103, 479–488. [Google Scholar] [CrossRef]
- Abrams, M.D.; Orwig, D.A. Structure, radial growth dynamics and recent climatic variations of a 320-year-old Pinus rigida rock outcrop community. Oecologia 1995, 101, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, K.B.; Hofgaard, A.; Vandvik, V. Tree-growth response to climatic variability in two climatically contrasting treeline ecotone areas, central Himalaya, Nepal. Can. J. For. Res. 2015, 45, 1643–1653. [Google Scholar] [CrossRef]
- Gaire, N.P.; Fan, Z.-X.; Braeuning, A.; Panthi, S.; Rana, P.; Shrestha, A.; Bhuju, D.R. Abies spectabilis shows stable growth relations to temperature, but changing response to moisture conditions along an elevation gradient in the central Himalaya. Dendrochronologia 2020, 60, 125675. [Google Scholar] [CrossRef]
- Shrestha, K.B.; Chhetri, P.K.; Bista, R. Growth responses of Abies spectabilis to climate variations along an elevational gradient in Langtang National Park in the central Himalaya, Nepal. J. For. Res. 2017, 22, 274–281. [Google Scholar] [CrossRef]
- Herrera-Soto, G.; González-Cásares, M.; Pompa-García, M.; Camarero, J.J.; Solís-Moreno, R. Growth of Pinus cembroides Zucc. in response to hydroclimatic variability in four sites forming the species latitudinal and longitudinal distribution limits. Forests 2018, 9, 440. [Google Scholar] [CrossRef] [Green Version]
- Liang, E.; Balducci, L.; Ren, P.; Rossi, S. Xylogenesis and moisture stress. In Secondary Xylem Biology; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2016; pp. 45–58. [Google Scholar]
- Begum, S.; Nakaba, S.; Yamagishi, Y.; Oribe, Y.; Funada, R. Regulation of cambial activity in relation to environmental conditions: Understanding the role of temperature in wood formation of trees. Physiol. Plant. 2013, 147, 46–54. [Google Scholar] [CrossRef]
- Rathgeber, C.B.K.; Cuny, H.E.; Fonti, P. Biological basis of tree-ring formation: A crash course. Front. Plant Sci. 2016, 7, 734. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Deslauriers, A.; Griçar, J.; Seo, J.-W.; Rathgeber, C.B.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- Sohar, K.; Altman, J.; Lehečková, E.; Doležal, J. Growth–climate relationships of Himalayan conifers along elevational and latitudinal gradients. Int. J. Climatol. 2017, 37, 2593–2605. [Google Scholar] [CrossRef]
- Ren, P.; Ziaco, E.; Rossi, S.; Biondi, F.; Prislan, P.; Liang, E. Growth rate rather than growing season length determines wood biomass in dry environments. Agric. For. Meteorol. 2019, 271, 46–53. [Google Scholar] [CrossRef]
- D’Arrigo, R.; Wilson, R.; Liepert, B.; Cherubini, P. On the ‘divergence problem’ in northern forests: A review of the tree-ring evidence and possible causes. Glob. Planet. Chang. 2008, 60, 289–305. [Google Scholar] [CrossRef]
- Peltier, D.M.P.; Ogle, K. Tree growth sensitivity to climate is temporally variable. Ecol. Lett. 2020, 23, 1561–1572. [Google Scholar] [CrossRef] [PubMed]
- Langan, S.J.; Ewers, F.W.; Davis, S.D. Xylem dysfunction caused by water stress and freezing in two species of co-occurring chaparral shrubs. Plant Cell Environ. 1997, 20, 425–437. [Google Scholar] [CrossRef]
- Yadav, R.R.; Singh, J.; Dubey, B.; Chaturvedi, R. Varying strength of relationship between temperature and growth of high-level fir at marginal ecosystems in western Himalaya, India. Curr. Sci. 2004, 86, 1152–1156. [Google Scholar]
- Rollinson, C.R.; Alexander, M.R.; Dye, A.W.; Moore, D.J.P.; Pederson, N.; Trouet, V. Climate sensitivity of understory trees differs from overstory trees in temperate mesic forests. Ecology 2021, 102, e03264. [Google Scholar] [CrossRef]
- Ryan, M.G.; Yoder, B.J. Hydraulic limits to tree height and tree growth. BioScience 1997, 47, 235–242. [Google Scholar] [CrossRef] [Green Version]
- D’Arrigo, R.D.; Kaufmann, R.K.; Davi, N.; Jacoby, G.C.; Laskowski, C.; Myneni, R.B.; Cherubini, P. Thresholds for warming-induced growth decline at elevational tree line in the Yukon Territory, Canada. Glob. Biogeochem. Cycles 2004, 18, GB3021. [Google Scholar] [CrossRef]
- Davi, N.K.; Jacoby, G.C.; Wiles, G.C. Boreal temperature variability inferred from maximum latewood density and tree-ring width data, Wrangell Mountain region, Alaska. Quat. Res. 2003, 60, 252–262. [Google Scholar] [CrossRef]

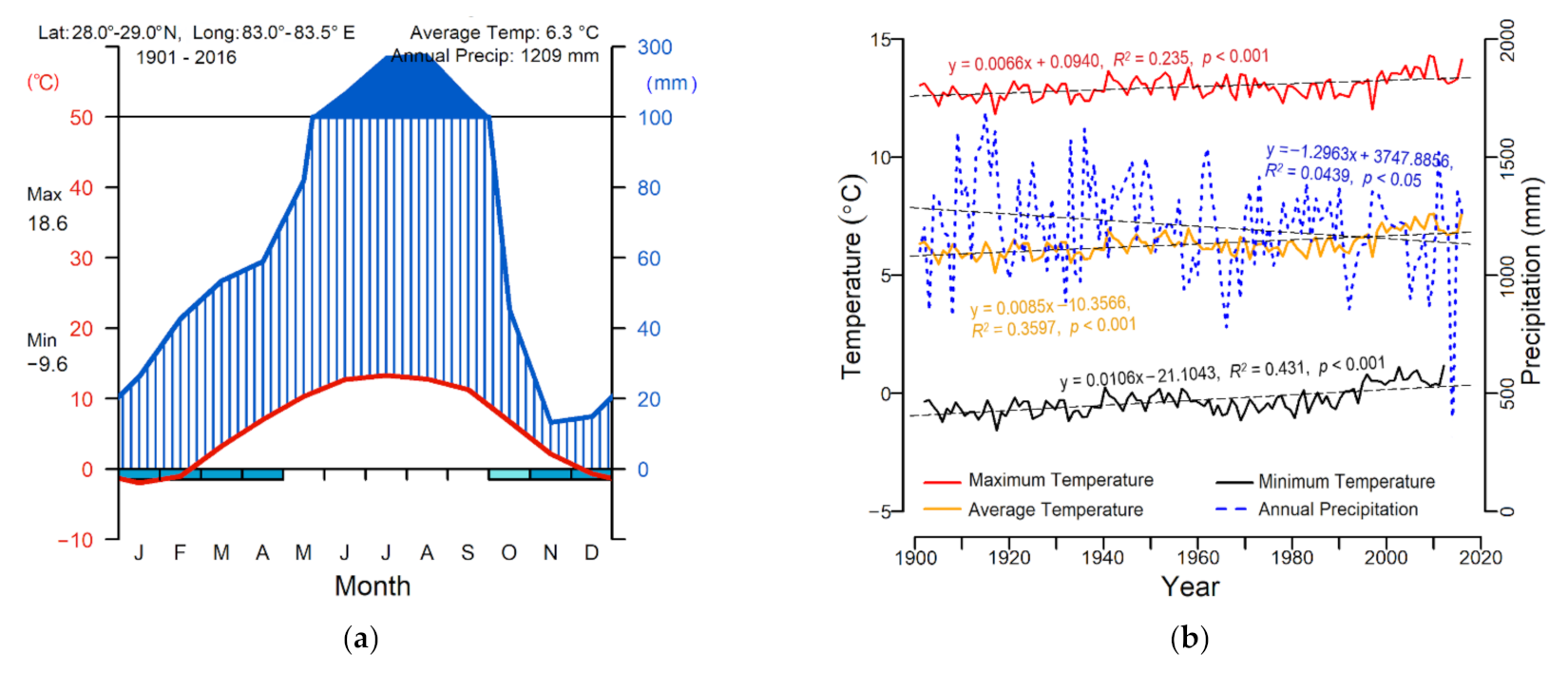




| Statistic | Species | |
|---|---|---|
| Abies spectabilis | Betula utilis | |
| Series number (tree number) | 53 (28) | 44 (24) |
| Span (Year) | 1909–2015 | 1721–2015 |
| Average mean sensitivity (MS) | 0.21 | 0.35 |
| First order autocorrelation (Ar1) | 0.70 | 0.76 |
| Mean inter-series correlation (R) | 0.54 | 0.55 |
| Mean within-tree correlation (Rbarwt) | 0.55 | 0.53 |
| Mean between-tree correlation (Rbarbt) | 0.31 | 0.33 |
| Expressed population signal (EPS) | 0.91 | 0.92 |
| Year after which EPS > 0.85 | 1919 | 1808 |
| Signal-to-noise ratio (SNR) | 10.28 | 10.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bista, R.; Chhetri, P.K.; Johnson, J.S.; Sinha, A.; Shrestha, K.B. Climate-Driven Differences in Growth Performance of Cohabitant Fir and Birch in a Subalpine Forest in Dhorpatan Nepal. Forests 2021, 12, 1137. https://doi.org/10.3390/f12091137
Bista R, Chhetri PK, Johnson JS, Sinha A, Shrestha KB. Climate-Driven Differences in Growth Performance of Cohabitant Fir and Birch in a Subalpine Forest in Dhorpatan Nepal. Forests. 2021; 12(9):1137. https://doi.org/10.3390/f12091137
Chicago/Turabian StyleBista, Raju, Parveen K. Chhetri, Jeremy S. Johnson, Ashish Sinha, and Krishna B. Shrestha. 2021. "Climate-Driven Differences in Growth Performance of Cohabitant Fir and Birch in a Subalpine Forest in Dhorpatan Nepal" Forests 12, no. 9: 1137. https://doi.org/10.3390/f12091137
APA StyleBista, R., Chhetri, P. K., Johnson, J. S., Sinha, A., & Shrestha, K. B. (2021). Climate-Driven Differences in Growth Performance of Cohabitant Fir and Birch in a Subalpine Forest in Dhorpatan Nepal. Forests, 12(9), 1137. https://doi.org/10.3390/f12091137