
Nitrogen and Phosphorus Distribution and Relationship in Soils and Plants under Different Aged Chinese Fir Plantation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling
2.2. Biomass Estimation of Chinese Fir
2.3. Analysis of Total Nitrogen and Phosphorous
2.4. Statistical Analysis
3. Results
3.1. Nitrogen and Phosphorus in Soil
3.2. Nitrogen and Phosphorus in Chinese Fir
3.3. Biomass and Nutrient Pool
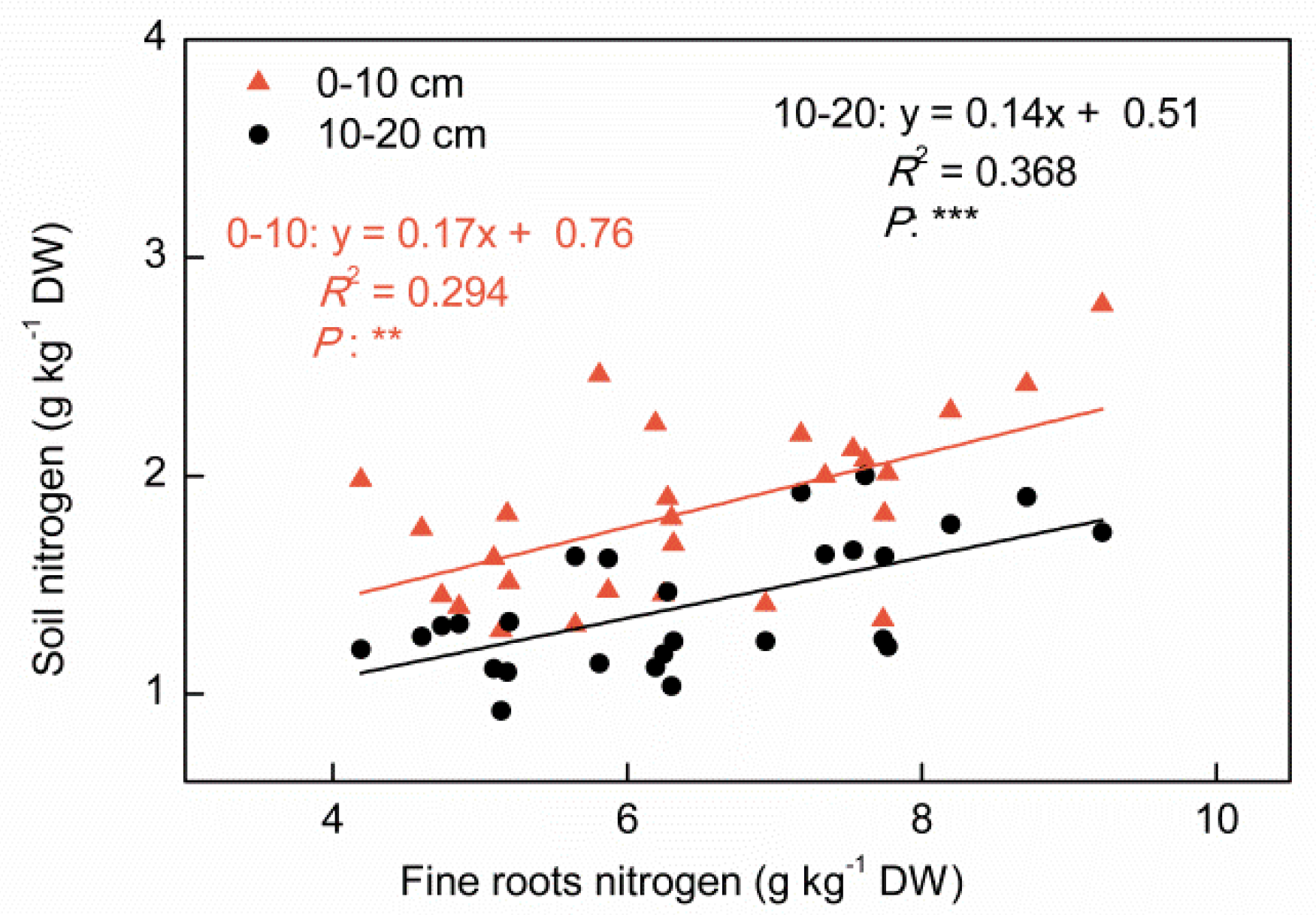
3.4. Correlation between Nutrients
4. Discussion
4.1. Spatial and Temporal Variation in Soil N and P Content
4.2. Nitrogen and Phosphorous Concentrations in Different Organs and N–P Coupling
4.3. Biomass Allocation and Distribution Patterns of Nutrient Pools
4.4. Relationship between Surface Soil and Nutrient Concentrations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crous, K.Y.; Ósvaldsson, A.; Ellsworth, D.S. Is phosphorus limiting in a mature Eucalyptus woodland? Phosphorus fertilisation stimulates stem growth. Plant Soil 2015, 391, 293–305. [Google Scholar] [CrossRef]
- López-Arredondo, D.L.; Leyva-González, M.A.; González-Morales, S.I.; López-Bucio, J.; Herrera-Estrella, L. Phosphate nutrition: Improving low-phosphate tolerance in crops. Annu. Rev. Plant Biol. 2014, 65, 95–123. [Google Scholar] [CrossRef] [PubMed]
- Rennenberg, H.; Wildhagen, H.; Ehlting, B. Nitrogen nutrition of poplar trees. Plant Biol. 2010, 12, 275–291. [Google Scholar] [CrossRef]
- Yang, X.J.; Huang, Z.Y.; Zhang, K.L.; Cornelissen, J.H.C. C:N:P stoichiometry of Artemisia species and close relatives across Northern China: Unraveling effects of climate, soil and taxonomy. J. Ecol. 2015, 103, 1020–1031. [Google Scholar] [CrossRef]
- Shen, J.B.; Yuan, L.X.; Zhang, J.L.; Li, H.G.; Bai, Z.H.; Chen, X.P.; Zhang, W.F.; Zhang, F.S. Phosphorus dynamics: From soil to plant. Plant Physiol. 2011, 156, 997–1005. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Liu, W.C.; Xu, M.P.; Deng, J.; Han, X.H.; Yang, G.H.; Feng, Y.Z.; Ren, G.X. Response of forest growth to C:N:P stoichiometry in plants and soils during Robinia pseudoacacia afforestation on the Loess Plateau, China. Geoderma 2019, 337, 280–289. [Google Scholar] [CrossRef]
- Kerkhoff, A.J.; Fagan, W.F.; Elser, J.J.; Enquist, B.J. Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants. Am. Nat. 2006, 168, 103–122. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.J. Biomass and nutrient distribution in a Chinese-fir plantation chronosequence in southwest Hunan, China. For. Ecol. Manag. 1998, 105, 209–216. [Google Scholar] [CrossRef]
- Ma, X.Q.; Liu, C.J.; Ilvesniemi, H.; Carl, J.W.; Liu, A.Q. Biomass, litterfall and the nutrient fluxes in Chinese fir stands of different age in subtropical China. J. For. Res. 2002, 13, 165–170. [Google Scholar]
- Ma, X.Q.; Heal, K.V.; Liu, A.Q.; Jarvis, P.G. Nutrient cycling and distribution in different-aged plantations of Chinese fir in Southern China. For. Ecol. Manag. 2007, 243, 61–74. [Google Scholar] [CrossRef]
- Xue, L. Nutrient cycling in a Chinese-fir (Cunninghamia lanceolata) stand on a poor site in Yishan, Guangxi. For. Ecol. Manag. 1996, 89, 115–123. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, X.Y.; Zeng, D.H. Aboveground biomass and nutrient allocation in an age-sequence of Larix olgensis plantations. J. For. Res. 2011, 22, 71–76. [Google Scholar] [CrossRef]
- Li, L.; Zerbe, S.; Han, W.; Thevs, N.; Li, W.; He, P.; Schmitt, A.O.; Liu, Y.; Ji, C. Nitrogen and phosphorus stoichiometry of common reed (Phragmites australis) and its relationship to nutrient availability in northern China. Aquat. Bot. 2014, 112, 84–90. [Google Scholar] [CrossRef]
- Zhang, W.D.; Wang, X.F.; Wang, S.L. Fate of Chinese-fir litter during decomposition as a result of inorganic N additions. Appl. Soil Ecol. 2014, 74, 30–36. [Google Scholar] [CrossRef]
- Zhou, L.; Shalom, A.-D.D.; Wu, P.; Li, S.; Jia, Y.; Ma, X. Litterfall production and nutrient return in different-aged Chinese fir (Cunninghamia lanceolata) plantations in South China. J. For. Res. 2015, 26, 79–89. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, S.L.; Feng, Z.W.; Ouyang, Z.Y.; Wang, X.; Feng, Z. Changes in soil quality due to introduction of broad-leaf trees into clear-felled Chinese fir forest in the mid-subtropics of China. Soil Use Manag. 2004, 20, 418–425. [Google Scholar] [CrossRef]
- Wang, Q.K.; Wang, S.L. Soil microbial properties and nutrients in pure and mixed Chinese fir plantations. J. For. Res. 2008, 19, 131–135. [Google Scholar] [CrossRef]
- Wang, Q.K.; Wang, S.L.; Yu, X.J. Decline of soil fertility during forest conversion of secondary forest to Chinese fir plantations in subtropical China. Land Degrad. Dev. 2011, 22, 444–452. [Google Scholar] [CrossRef]
- Wang, W.J.; Qiu, L.; Zu, Y.G.; Su, D.X.; An, J.; Wang, H.Y.; Zheng, G.Y.; Sun, W.; Chen, X.Q. Changes in soil organic carbon, nitrogen, pH and bulk density with the development of larch (Larix gmelinii) plantations in China. Glob. Chang. Biol. 2011, 17, 2657–2676. [Google Scholar]
- Tong, R.; Zhou, B.Z.; Jiang, L.N.; Ge, X.G.; Cao, Y.H. Spatial patterns of leaf carbon, nitrogen, and phosphorus stoichiometry and nutrient resorption in Chinese fir across subtropical China. Catena 2021, 201, 105221. [Google Scholar] [CrossRef]
- Kalliokoski, T.; Nygren, P.; Sievanen, R. Coarse root architecture of three boreal tree species growing in mixed stands. Silva Fenn. 2008, 42, 189–210. [Google Scholar] [CrossRef] [Green Version]
- Hellsten, S.; Helmisaari, H.S.; Melin, Y.; Skovsgaard, J.P.; Kaakinen, S.; Kukkola, M.; Saarsalmi, A.; Petersson, H.; Akselsson, C. Nutrient concentrations in stumps and coarse roots of Norway spruce, Scots pine and silver birch in Sweden, Finland and Denmark. For. Ecol. Manag. 2013, 290, 40–48. [Google Scholar] [CrossRef]
- Cao, Y.; Chen, Y.M. Coupling of plant and soil C:N:P stoichiometry in black locust (Robinia pseudoacacia) plantations on the Loess Plateau, China. Trees 2017, 31, 1559–1570. [Google Scholar] [CrossRef]
- Li, H.; Li, J.; He, Y.L.; Li, S.J.; Liang, Z.S.; Peng, C.H.; Polle, A.; Luo, Z.B. Changes in carbon, nutrients and stoichiometric relations under different soil depths, plant tissues and ages in black locust plantations. Acta Physiol. Plant. 2013, 35, 2951–2964. [Google Scholar] [CrossRef]
- Zhou, J.; Jiao, F.C.; Wu, Z.C.; Li, Y.Y.; Wang, X.M.; He, X.W.; Zhong, W.Q.; Wu, P. OsPHR2 is involved in phosphate-starvation signaling and excessive phosphate accumulation in shoots of plants. Plant Physiol. 2008, 146, 1673–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.W.; Zhang, X.K.; Bao, X.L.; Wang, J.K. Spatial distribution of soil nutrients after the establishment of sand-fixing shrubs on sand dune. Plant Soil Environ. 2009, 55, 288–294. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.C.; McCormack, M.L.; Fan, H.B.; Wang, H.M.; Wu, J.P.; Tu, J.; Liu, W.F.; Guo, D.L. Relation of fine root distribution to soil C in a Cunninghamia lanceolata plantation in subtropical China. Plant Soil 2014, 381, 225–234. [Google Scholar] [CrossRef]
- Luan, J.W.; Xiang, C.H.; Liu, S.R.; Luo, Z.S.; Gong, Y.B.; Zhu, X.L. Assessments of the impacts of Chinese fir plantation and natural regenerated forest on soil organic matter quality at Longmen mountain, Sichuan, China. Geoderma 2010, 156, 228–236. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, P.; Chen, Y.M. Soil C:N:P stoichiometry in plantations of N-fixing black locust and indigenous pine, and secondary oak forests in Northwest China. J. Soil. Sediment. 2018, 18, 1478–1489. [Google Scholar] [CrossRef]
- Houlton, B.Z.; Morford, S.L. A new synthesis for terrestrial nitrogen inputs. Soil 2014, 1, 497–540. [Google Scholar]
- Blanco, J.A.; Imbert, J.B.; Castillo, F.J. Nutrient return via litterfall in two contrasting Pinus sylvestris forests in the Pyrenees under different thinning intensities. For. Ecol. Manag. 2008, 256, 1840–1852. [Google Scholar] [CrossRef]
- Kavvadias, V.A.; Alifragis, D.; Tsiontsis, A.; Brofas, G.; Stamatelos, G. Litterfall, litter accumulation and litter decomposition rates in four forest ecosystems in northern Greece. For. Ecol. Manag. 2001, 144, 113–127. [Google Scholar] [CrossRef]
- Yang, Y.S.; Guo, J.F.; Chen, G.; Xie, J.S.; Gao, R.; Li, Z.; Jin, Z. Litter production, seasonal pattern and nutrient return in seven natural forests compared with a plantation in southern China. Forestry 2005, 78, 403–415. [Google Scholar] [CrossRef] [Green Version]
- Long, M.; Wu, H.H.; Smith, M.D.; Pierre La, K.J.; Lü, X.T.; Zhang, H.Y.; Han, X.G.; Yu, Q. Nitrogen deposition promotes phosphorus uptake of plants in a semi-arid temperate grassland. Plant Soil 2016, 408, 475–484. [Google Scholar] [CrossRef]
- Yu, Q.; Elser, J.J.; He, N.P.; Wu, H.H.; Chen, Q.S.; Zhang, G.M.; Han, X.G. Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland. Oecologia 2011, 166, 1–10. [Google Scholar] [CrossRef]
- Helmisaari, H.S.; Siltala, T. Variation in nutrient concentrations of Pinus sylvestris stems. Scand. J. For. Res. 1989, 4, 443–451. [Google Scholar] [CrossRef]
- Iivonen, S.; Kaakinen, S.; Jolkkonen, A.; Vapaavuori, E.; Linder, S. Influence of long-term nutrient optimization on biomass, carbon, and nitrogen acquisition and allocation in Norway spruce. Can. J. For. Res. 2006, 36, 1563–1571. [Google Scholar] [CrossRef]
- Liu, X.L.; Xu, H.Y.; Berninger, F.; Luukkanen, O.; Li, C.Y. Nutrient distribution in Picea likiangensis trees growing in a plantation in West Sichuan, Southwest China. Silva Fenn. 2004, 38, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.; Walters, M.; Tjoelker, M.; Vanderklein, D.; Buschena, C. Photosynthesis and respiration rates depend on leaf and root morphology and nitrogen concentration in nine boreal tree species differing in relative growth rate. Funct. Ecol. 1998, 12, 395–405. [Google Scholar] [CrossRef]
- Xiang, W.H.; Wu, W.; Tong, J.; Deng, X.W.; Tian, D.L.; Zhang, L.; Liu, C.; Peng, C.H. Differences in fine root traits between early and late-successional tree species in a Chinese subtropical forest. Forestry 2013, 86, 343–351. [Google Scholar] [CrossRef]
- Borja, I.; De Wit, H.A.; Steffenrem, A.; Majdi, H. Stand age and fine root biomass, distribution and morphology in a Norway spruce chronosequence in southeast Norway. Tree Physiol. 2008, 28, 773–784. [Google Scholar] [CrossRef]
- Guo, L.; Chen, J.; Cui, X.H.; Fan, B.H.; Lin, H. Application of ground penetrating radar for coarse root detection and quantification: A review. Plant Soil 2013, 362, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Peri, P.L.; Gargaglione, V.; Pastur, G.M. Dynamics of above- and below-ground biomass and nutrient accumulation in an age sequence of Nothofagus antarctica forest of Southern Patagonia. For. Ecol. Manag. 2006, 233, 85–99. [Google Scholar] [CrossRef]
- Yang, Y.H.; Luo, Y.Q. Carbon: Nitrogen stoichiometry in forest ecosystems during stand development. Glob. Ecol. Biogeogr. 2011, 20, 354–361. [Google Scholar] [CrossRef]
- Yan, Z.B.; Kim, N.Y.; Han, W.X.; Guo, Y.L.; Han, T.S.; Du, E.Z.; Fang, J.Y. Effects of nitrogen and phosphorus supply on growth rate, leaf stoichiometry, and nutrient resorption of Arabidopsis thaliana. Plant Soil 2014, 388, 147–155. [Google Scholar] [CrossRef]
- Ladanai, S.; Ågren, G.I.; Olsson, B.A. Relationships between tree and soil properties in Picea abies and Pinus sylvestris forests in Sweden. Ecosystems 2010, 13, 302–316. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Liu, G.B.; Li, P.; Xue, S. Ecological stoichiometry of plant-soil-enzyme interactions drives secondary plant succession in the abandoned grasslands of Loess Plateau, China. Catena 2021, 202, 105302. [Google Scholar] [CrossRef]
- Gusewell, S.; Koerselman, W. Variation in nitrogen and phosphorus concentrations of wetland plants. Perspect. Plant Ecol. 2002, 5, 37–61. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Years) | Fine Roots | Coarse Roots | Stems | Leaves | Total | p-Value |
|---|---|---|---|---|---|---|
| 10 | 7.8 ± 1.8 B b (1.9) | 70.8 ± 27.0 C b (17.1) | 292.2 ± 84.0 B a (70.6) | 43.0 ± 13.6 C b (10.4) | 413.7 ± 71.8 C a (100) | *** |
| 20 | 8.9 ± 1.1 A d (0.7) | 261.7 ± 10.6 B c (20.7) | 896.9 ± 91.5 B b (70.9) | 98.1 ± 15.1 B d (7.7) | 1265.6 ±67.1 B a (100) | *** |
| 30 | 14.0 ± 3.0 A e (0.6) | 473.1 ± 40.0 A c (19.9) | 1738.1 ± 98.1 A b (73.0) | 156.3 ± 15.0 A d (6.5) | 2381.5 ± 47.5 A a (100) | *** |
| p-value | * | *** | ns | *** | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Luo, Z. Nitrogen and Phosphorus Distribution and Relationship in Soils and Plants under Different Aged Chinese Fir Plantation. Forests 2021, 12, 1271. https://doi.org/10.3390/f12091271
Yang Z, Luo Z. Nitrogen and Phosphorus Distribution and Relationship in Soils and Plants under Different Aged Chinese Fir Plantation. Forests. 2021; 12(9):1271. https://doi.org/10.3390/f12091271
Chicago/Turabian StyleYang, Zhen’an, and Zhibin Luo. 2021. "Nitrogen and Phosphorus Distribution and Relationship in Soils and Plants under Different Aged Chinese Fir Plantation" Forests 12, no. 9: 1271. https://doi.org/10.3390/f12091271
APA StyleYang, Z., & Luo, Z. (2021). Nitrogen and Phosphorus Distribution and Relationship in Soils and Plants under Different Aged Chinese Fir Plantation. Forests, 12(9), 1271. https://doi.org/10.3390/f12091271