
Identification and Characterization of Nectria pseudotrichia Associated with Camellia Canker Disease in China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Field Surveys of Canker Disease in C. japonica
2.2. Fungal Isolation
2.3. Morphological Characterization
2.4. DNA Extraction, Amplification, Sequencing, and Phylogenetic Analysis
2.5. Pathogenicity Tests
3. Results
3.1. Field Surveys of Canker Disease in C. japonica
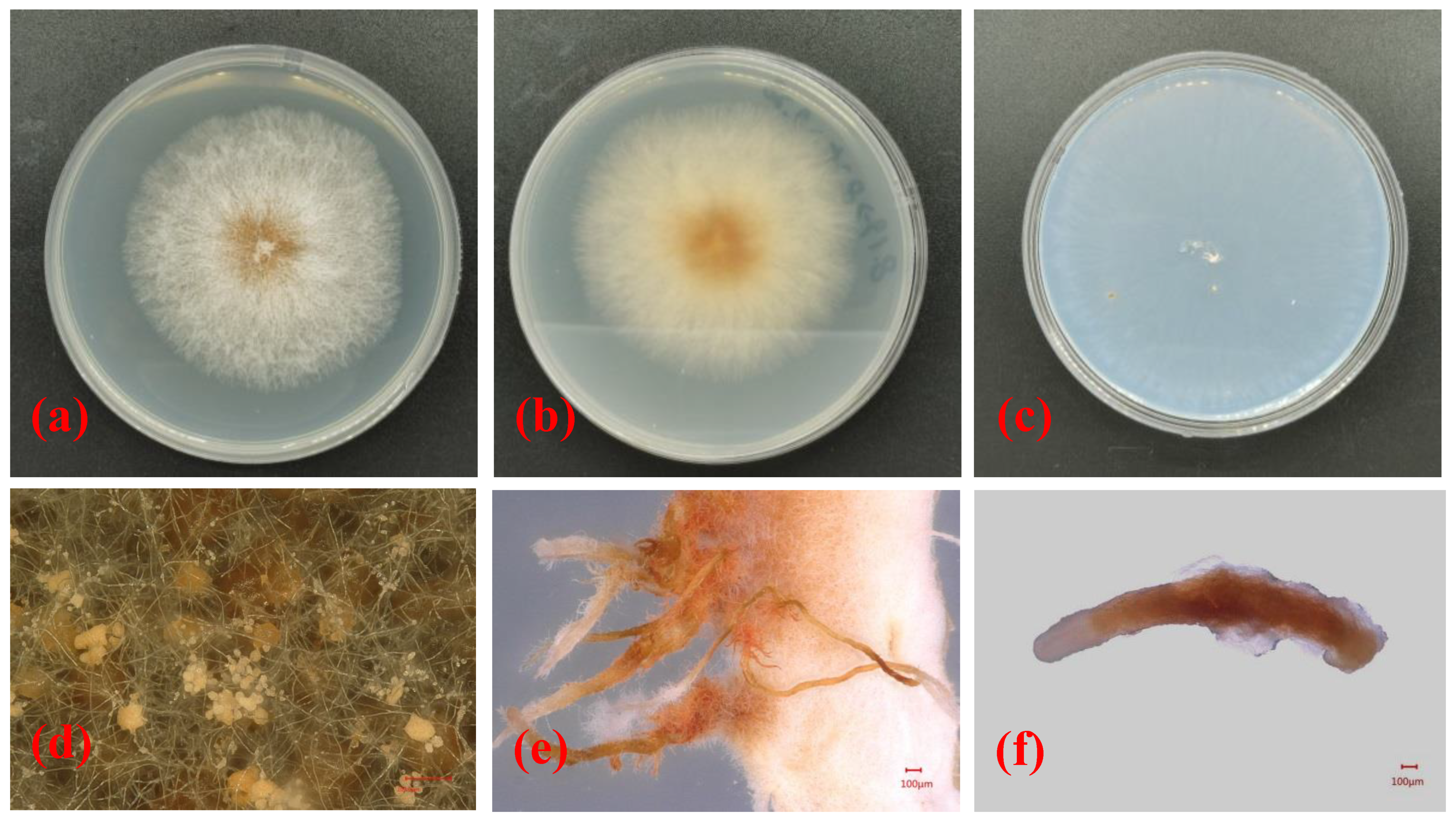
3.2. Isolation, Morphological Structures and Cultural Characteristics of the Pathogenic Fungus of Camellia Canker
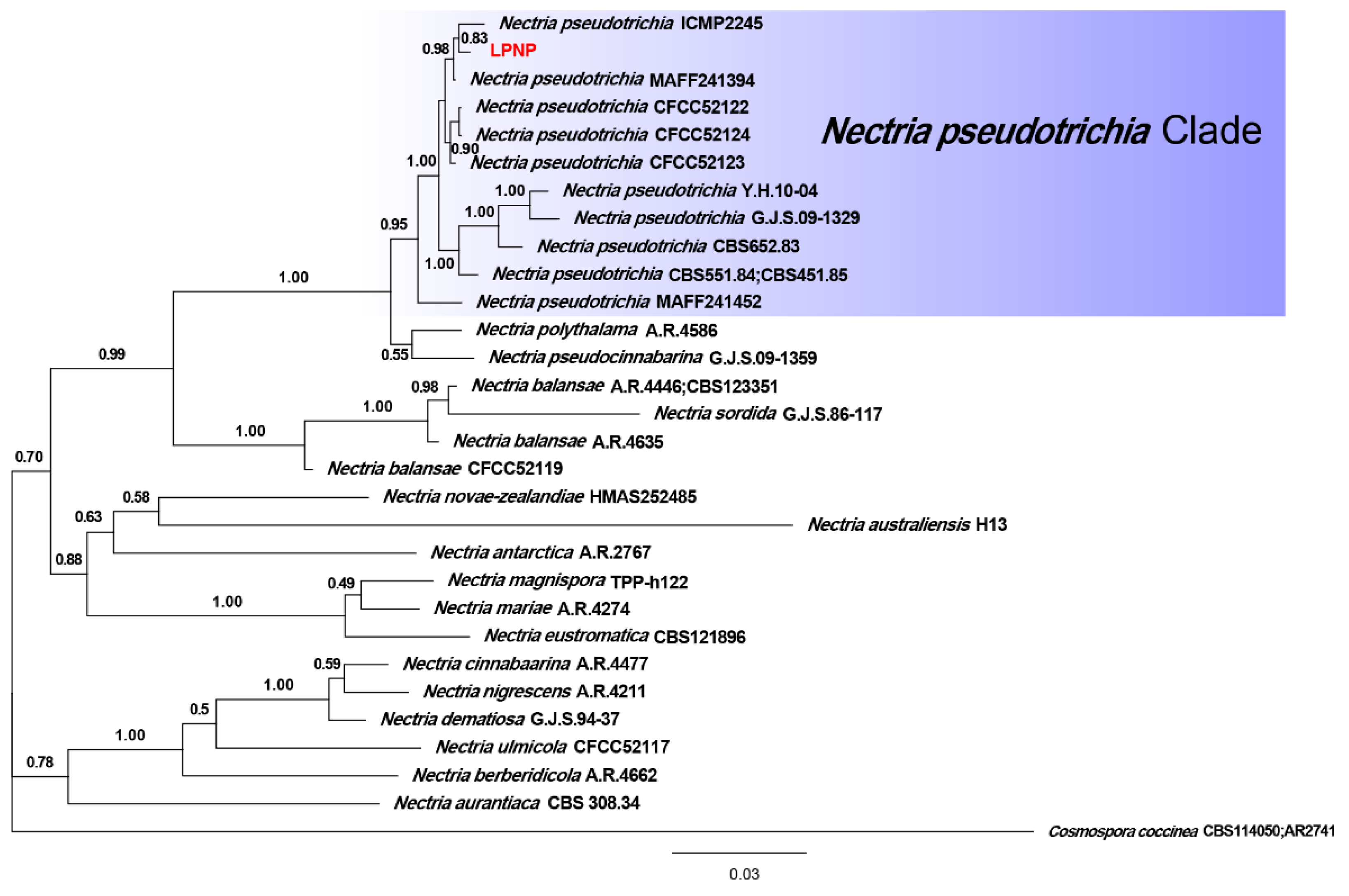
3.3. Phylogenetic Analysis
3.4. Pathogenicity Tests
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lim, T.K. (Ed.) Camellia japonica. In Edible Medicinal and Non Medicinal Plants; Springer: Dordrecht, The Netherlands, 2014; Volume 8, pp. 764–776. [Google Scholar]
- Gao, J.Y.; Parks, C.R.; Du, Y.Q. Collected Species of the Genus Camellia: An Illustrated Outline; Zhejiang Science and Technology Press: Hangzhou, China, 2005; pp. 43–44. [Google Scholar]
- Vela, P.; Salinero, C.; Sainz, M. Phenological growth stages of Camellia japonica. Ann. Appl. Biol. 2012, 162, 182–190. [Google Scholar] [CrossRef]
- Garcia-Jares, C.; Sanchez-Nande, M.; Lamas, J.P.; Lores, M. Profiling the Fatty Acids Content of Ornamental Camellia Seeds Cultivated in Galicia by an Optimized Matrix Solid-Phase Dispersion Extraction. Bioengineering 2017, 4, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.-L.; Yin, C.-P.; Kong, L.-C.; Jiang, D.-H. Extraction optimisation, purification and major antioxidant component of red pigments extracted from Camellia japonica. Food Chem. 2011, 129, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Yoon, I.-S.; Park, D.-H.; Kim, J.-E.; Yoo, J.-C.; Bae, M.-S.; Oh, D.-S.; Shim, J.-H.; Choi, C.-Y.; An, K.-W.; Kim, E.-I.; et al. Identification of the biologically active constituents of Camellia japonica leaf and anti-hyperuricemic effect in vitro and in vivo. Int. J. Mol. Med. 2017, 39, 1613–1620. [Google Scholar] [CrossRef] [Green Version]
- Baxter, L.W. Studies on Control of Root Rot and Dieback of Camellias; Louisiana State University: Baton Rouge, FL, USA, 1955. [Google Scholar]
- Dickens, J.S.W.; Cook, R.T.A. Glomerella cingulata on camellia. Plant Pathol. 1989, 38, 75–85. [Google Scholar] [CrossRef]
- Pintos, C.; Redondo, V.; Chaves, M.; Rial, C.; Mansilla, P. Camellia japonica dieback caused by Neofusicoccum luteum and N. parvum in Spain. In Proceedings of the International Camellia Congress, Chuxiong, China, 5–9 February 2012. [Google Scholar]
- Ngo, H.C.; Baxter, L.W.; Fagan, S.G. The status of our knowledge in 1978 of twig blight, canker and dieback of camellias caused by a strain of Glomerella cingulata. Am. Camellia Yearb. 1978, 33, 75–95. [Google Scholar]
- Linaldeddu, B.; Bottecchia, F.; Bregant, C.; Maddau, L.; Montecchio, L. Diplodia fraxini and Diplodia subglobosa: The Main Species Associated with Cankers and Dieback of Fraxinus excelsior in North-Eastern Italy. Forests 2020, 11, 883. [Google Scholar] [CrossRef]
- Geng, X.S.; Shu, J.P.; Sheng, J.L.; Zhang, W.; Peng, H. Isolation and identification of the pathogens causing witches’ broom disease of five bamboo species of non-Phyllostachys. Sci. Silvae Sin. 2020, 56, 82–89. [Google Scholar]
- Leonard, L.M. Melzer’s, lugol’s or iodine for identification of white-spored Agaricales? McIlvainea 2006, 16, 43–51. [Google Scholar]
- Eberhardt, U. Methods for DNA Barcoding of Fungi. In Springer Protocols Handbooks; Springer: Singapore, 2012; Volume 858, pp. 183–205. [Google Scholar]
- White, T.; Bruns, T.; Lee, S.; Taylor, F.J.R.M.; White, T.; Lee, S.H.; Taylor, L.; Shawetaylor, J. Amplification and direct se-quencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, A.M., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Bellemain, E.; Carlsen, T.; Brochmann, C.; Coissac, E.; Taberlet, P.; Kauserud, H. ITS as an environmental DNA barcode for fungi: An in silico approach reveals potential PCR biases. BMC Microbiol. 2010, 10, 189. [Google Scholar] [CrossRef] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zhou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2019, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elvira-Recuenco, M.; Cacciola, S.O.; Sanz-Ros, A.V.; Garbelotto, M.; Aguayo, J.; Solla, A.; Mullett, M.; Drenkhan, T.; Oskay, F.; Kaya, A.G.A.; et al. Potential Interactions between Invasive Fusarium circinatum and Other Pine Pathogens in Europe. Forests 2019, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier: Burlington, MA, USA, 2005; pp. 473–483. [Google Scholar]
- Bedker, P.J.; Blanchette, R.A. Identification and control of cankers caused by Nectria cinnabarina of honey locust. J. Arboric. 1984, 10, 33–39. [Google Scholar]
- Pan, M.; Zhu, H.; Bonthond, G.; Tian, C.; Fan, X. High Diversity of Cytospora Associated With Canker and Dieback of Rosaceae in China, with 10 New Species Described. Front. Plant Sci. 2020, 11, 690. [Google Scholar] [CrossRef] [PubMed]
- Bartnik, C.; Boroń, P.; Michalcewicz, J.; Ciach, M. The first record of Botryodiplodia canker in Poland. For. Pathol. 2019, 49, e12528. [Google Scholar] [CrossRef]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Pers. Mol. Phylogeny Evol. Fungi 2013, 31, 1–41. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Du, Z.; Liang, Y.-M.; Tian, C.-M. Molecular phylogeny of Nectria species associated with dieback and canker diseases in China, with a new species described. Phytotaxa 2018, 356, 199–214. [Google Scholar] [CrossRef]
- Zhuang, W.Y. Nectriaceae et Bionectriaceae. In Flora Fungorum Sinicorum; Science Press: Beijing, China, 2013; Volume 47, pp. 110–125. [Google Scholar]
- Hirooka, Y.; Rossman, A.; Samuels, G.; Lechat, C.; Chaverri, P. A monograph of Allantonectria, Nectria, and Pleonectria (Nectriaceae, Hypocreales, Ascomycota) and their pycnidial, sporodochial, and synnematous anamorphs. Stud. Mycol. 2012, 71, 1–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, W.F. Nectria pseudotrichia, como agente causal de cancro de ramos, ocorrendo em pereira japonesa no Brasil. Fitopatol. Bras. 2003, 28, 107. [Google Scholar] [CrossRef] [Green Version]
- Akinsanmi, O.A.; Drenth, A. First report ofTubercularia lateritiaas the causal agent of canker on macadamia. Australas. Plant Dis. Notes 2006, 1, 49–51. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.Q.; Duan, K.; Gao, Q.H.; Song, L.L. First Report of Nectria pseudotrichia Causing Crown Rot of Strawberry in China. Plant Dis. 2018, 102, 1655. [Google Scholar] [CrossRef]
- Wanjiku, E.K.; Waceke, J.W.; Wanjala, B.W.; Mbaka, J.N. Identification and Pathogenicity of Fungal Pathogens Associated with Stem End Rots of Avocado Fruits in Kenya. Int. J. Microbiol. 2020, 2020, 1–8. [Google Scholar] [CrossRef]
- Jiang, G.Z.; Li, L.; Liu, Y.X.; Zhou, M. Identification of the pathogen causing stem brown spots canker of rubber tree and screening of fungicides. Trop. Agric. Sci. Technol. 2019, 42, 15–20. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain 1 | Geographic Origin | GenBank Accession No. | ||
|---|---|---|---|---|---|
| ITS | LSU | tef1-α | |||
| Nectria antarctica | A.R.2767 | USA | HM484556 | HM484560 | HM484516 |
| N. aurantiaca | CBS 308.34 | UK | JF832628 | JF832682 | JF832519 |
| N. balansae | A.R. 4446; CBS 123351 | France | HM484552 | GQ505996 | HM484525 |
| N. balansae | A.R. 4635 | China | JF832653 | JF832711 | JF832522 |
| N. berberidicola | A.R. 4662 | France | JF832662 | JF832712 | JF832538 |
| N. cinnabarina | A.R. 4477 | France | HM484548 | HM484562 | HM484527 |
| N. dematiosa | G.J.S. 94-37 | USA | HM484557 | HM484561 | HM484534 |
| N. magnispora | TPP-h122 | Japan | JF832663 | JF832683 | JF832539 |
| N. mariae | A.R. 4274 | France | JF832629 | JF832684 | JF832542 |
| N. nigrescens | A.R. 4211 | USA | HM484707 | HM484720 | HM484672 |
| N. polythalama | A.R. 4586 | New Zealand | JF832638 | JF832695 | JF832523 |
| N. pseudocinnabarina | G.J.S. 09-1359 | Venezuela | JF832642 | JF832697 | JF832533 |
| N. pseudotrichia | LPNP | China | OK562743 | OK562730 | OL440127 |
| N. pseudotrichia | CBS551.84; CBS 451.85 | Japan; Honduras | HM484554 | GQ506000 | HM484532 |
| N. pseudotrichia | MAFF 241394 | Japan | JF832639 | JF832705 | JF832527 |
| N. pseudotrichia | ICMP 2245 | Japan | JF832640 | JF832701 | JF832532 |
| N. pseudotrichia | MAFF 241452 | Papua New Guinea | JF832649 | JF832706 | JF832531 |
| N. pseudotrichia | CBS 652.83 | Japan | JF832648 | JF832703 | JF832528 |
| N. pseudotrichia | Y.H. 10-04 | Venezuela | JF832650 | JF832704 | JF832529 |
| N. pseudotrichia | G.J.S. 09-1329 | USA | JF832647 | JF832702 | JF832530 |
| N. pseudotrichia | CFCC 52122 | Venezuela | MG231953 | MG231974 | MG232016 |
| N. pseudotrichia | CFCC 52123 | China | MG231954 | MG231975 | MG232017 |
| N. pseudotrichia | CFCC 52124 | China | MG231955 | MG231976 | MG232018 |
| N. novae-zealandiae | HMAS 252485 | China | KM026503 | KM026504 | KM026506 |
| N. ulmicola | CFCC 52117 | China | MG231959 | MG231980 | MG232022 |
| N. australiensis | H13 | China | GU075855 | DQ119569 | HM054076 |
| N. eustromatica | CBS 121896 | China | HM534896 | HM534896 | HM534875 |
| N. sordida | G.J.S. 86-117 | Croatia | HM484857 | HM484868 | HM484848 |
| N. balansae | CFCC 52119 | French Guiana | MG231956 | MG231977 | MG232019 |
| Cosmospora coccinea | CBS 114050; AR2741 | China | FJ474072 | GQ505990 | AY489629 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geng, X.; Liu, Y.; Li, J.; Li, Z.; Shu, J.; Wu, G. Identification and Characterization of Nectria pseudotrichia Associated with Camellia Canker Disease in China. Forests 2022, 13, 29. https://doi.org/10.3390/f13010029
Geng X, Liu Y, Li J, Li Z, Shu J, Wu G. Identification and Characterization of Nectria pseudotrichia Associated with Camellia Canker Disease in China. Forests. 2022; 13(1):29. https://doi.org/10.3390/f13010029
Chicago/Turabian StyleGeng, Xiansheng, Ying Liu, Jiyuan Li, Zhihong Li, Jinping Shu, and Guiyang Wu. 2022. "Identification and Characterization of Nectria pseudotrichia Associated with Camellia Canker Disease in China" Forests 13, no. 1: 29. https://doi.org/10.3390/f13010029
APA StyleGeng, X., Liu, Y., Li, J., Li, Z., Shu, J., & Wu, G. (2022). Identification and Characterization of Nectria pseudotrichia Associated with Camellia Canker Disease in China. Forests, 13(1), 29. https://doi.org/10.3390/f13010029