
Comparative Morphological, Anatomical and Physiological Analyses Explain the Difference of Wounding-Induced Agarwood Formation between Ordinary Agarwood Nongrafted Plants and Five Grafted Qi-Nan Clones (Aquilaria sinensis)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Agarwood Yield and Essential Oil Content
2.3. Determination of Leaf
2.4. Gas Exchange Parameter Measurement
2.5. Xylem Structure
2.6. Determination of Non-Structural Carbohydrates
2.7. Statistical Analysis
3. Results
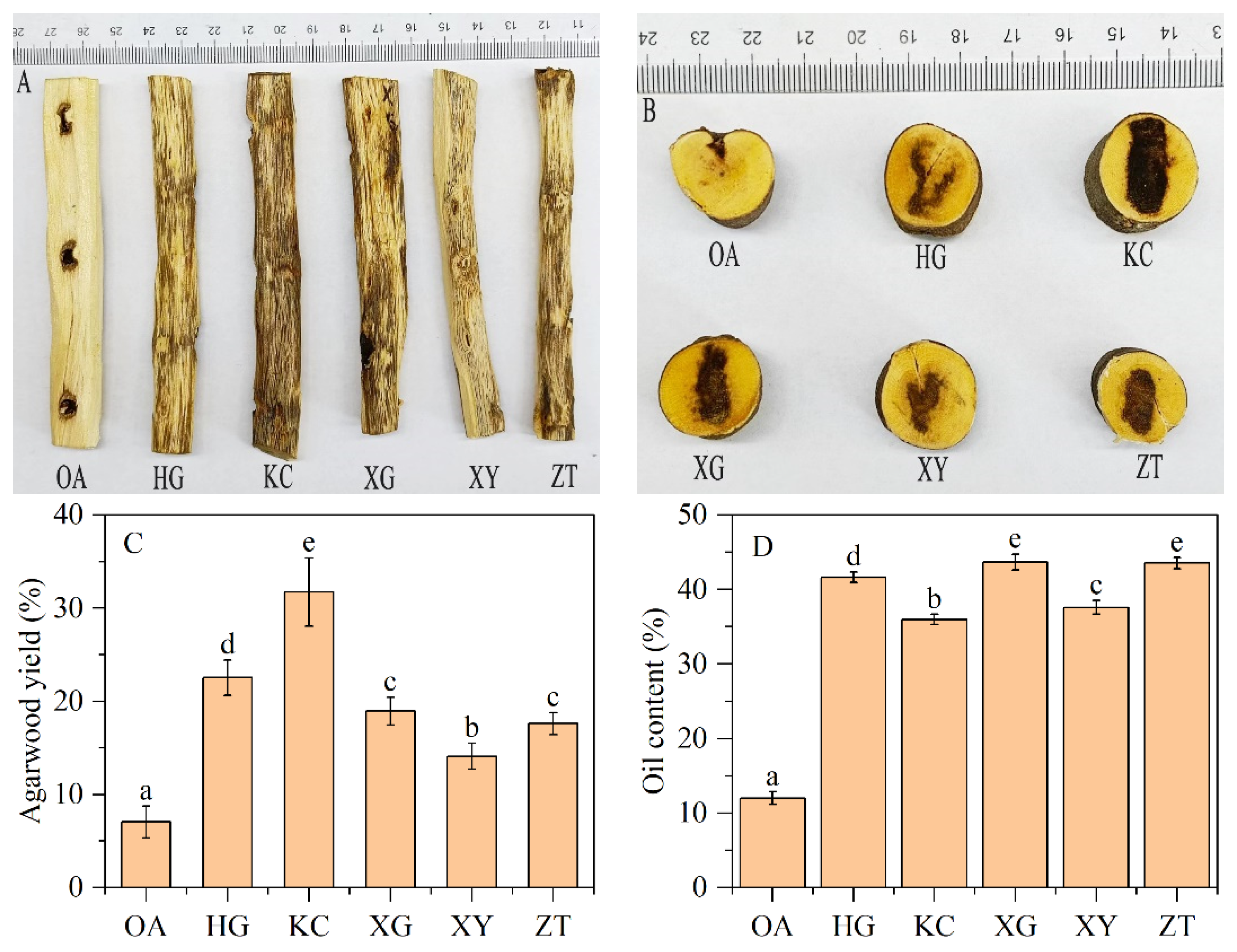
3.1. Agarwood Yield and Essential Oil Content
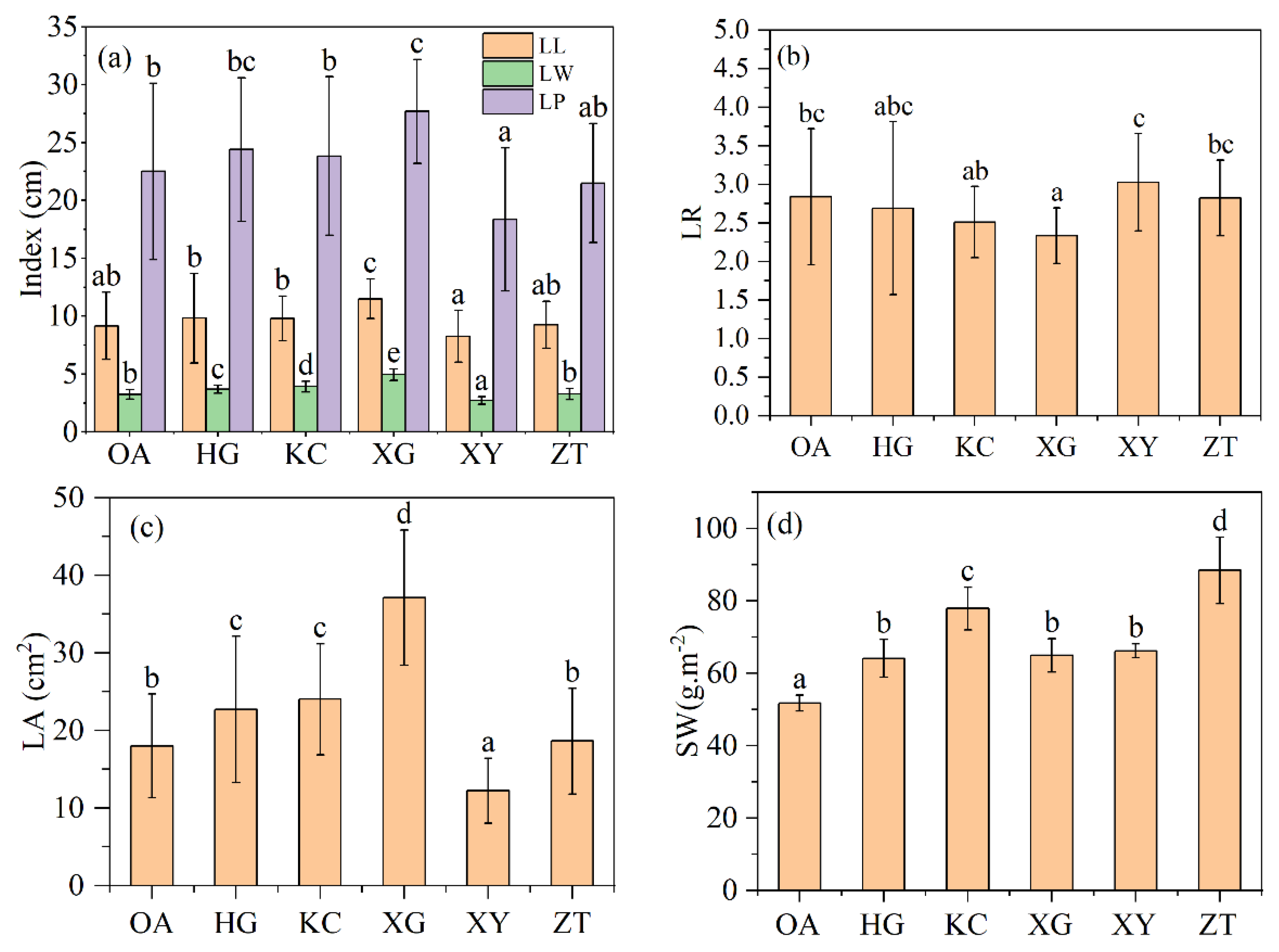
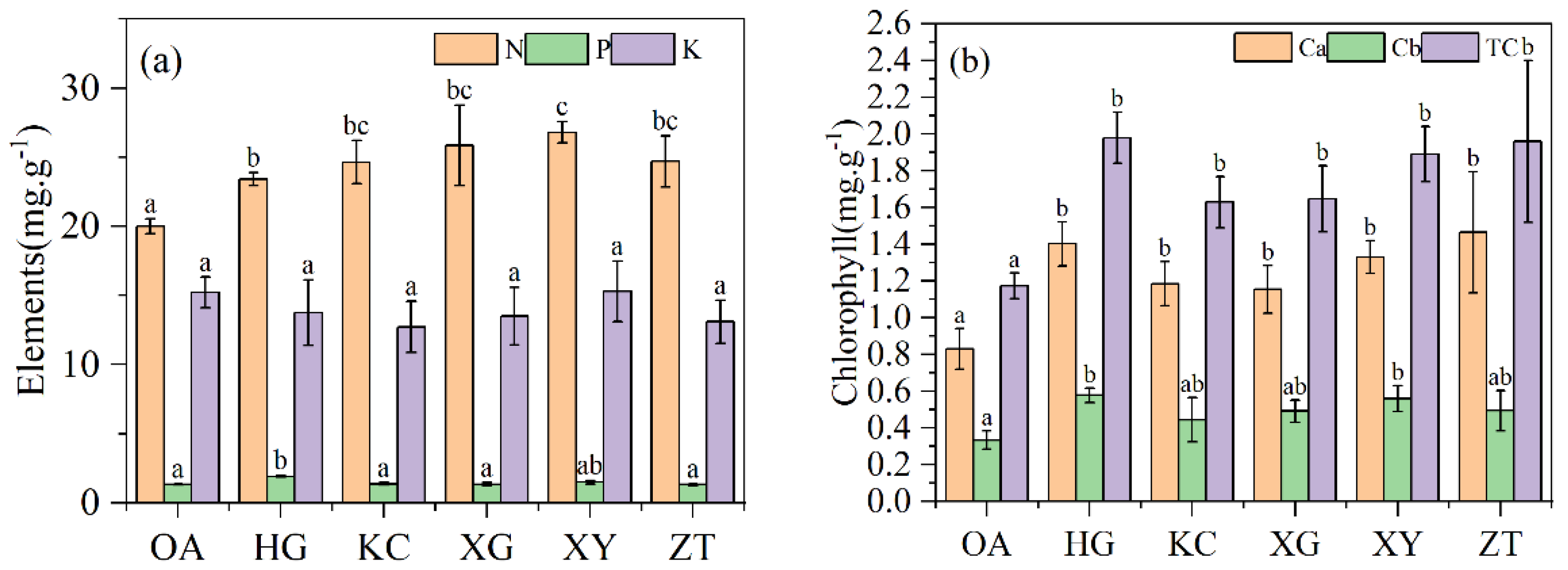
3.2. Leaf Morphological and Physiological Parameters
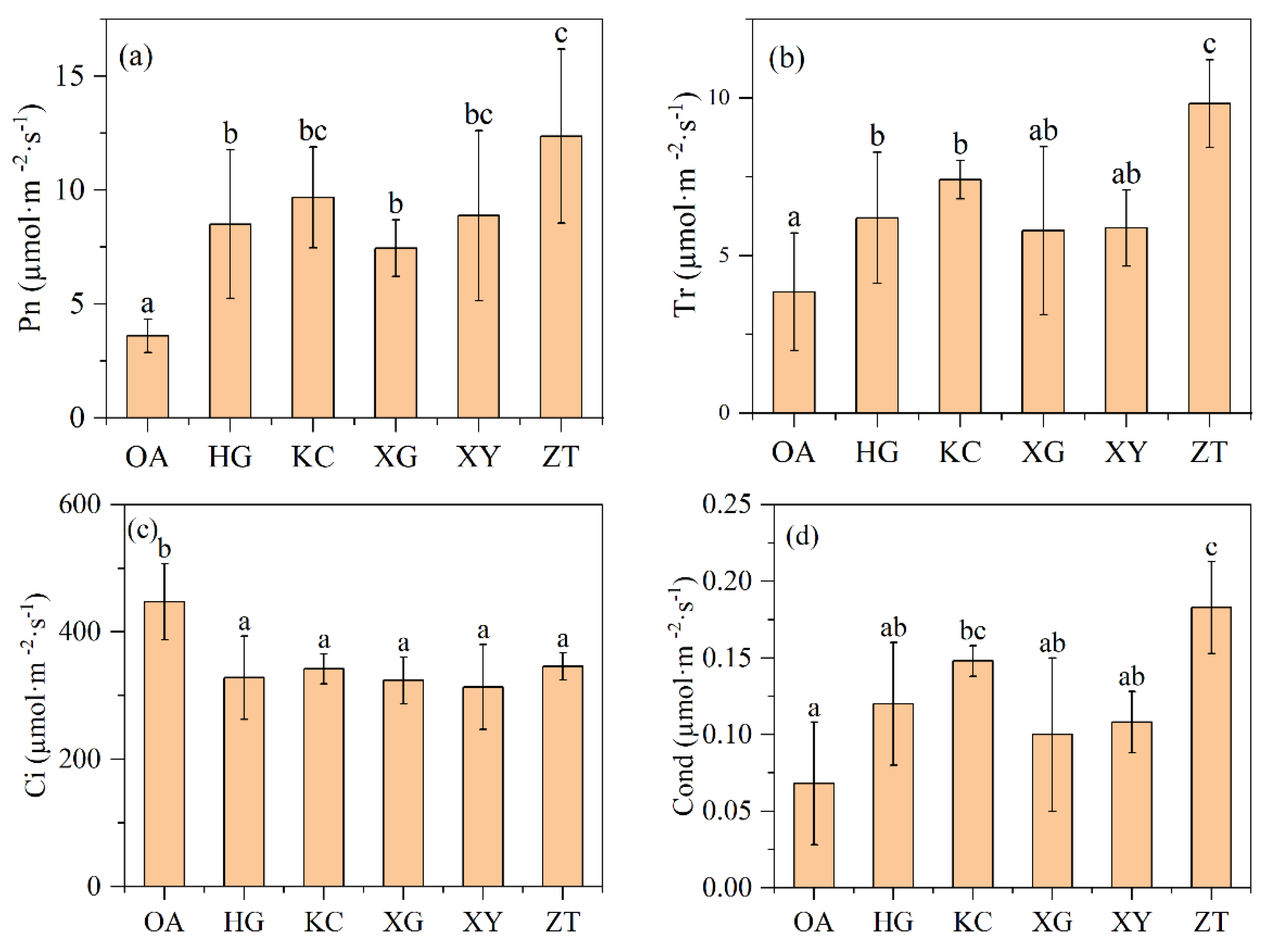
3.3. Gas Exchange Parameters
3.4. Xylem Structure
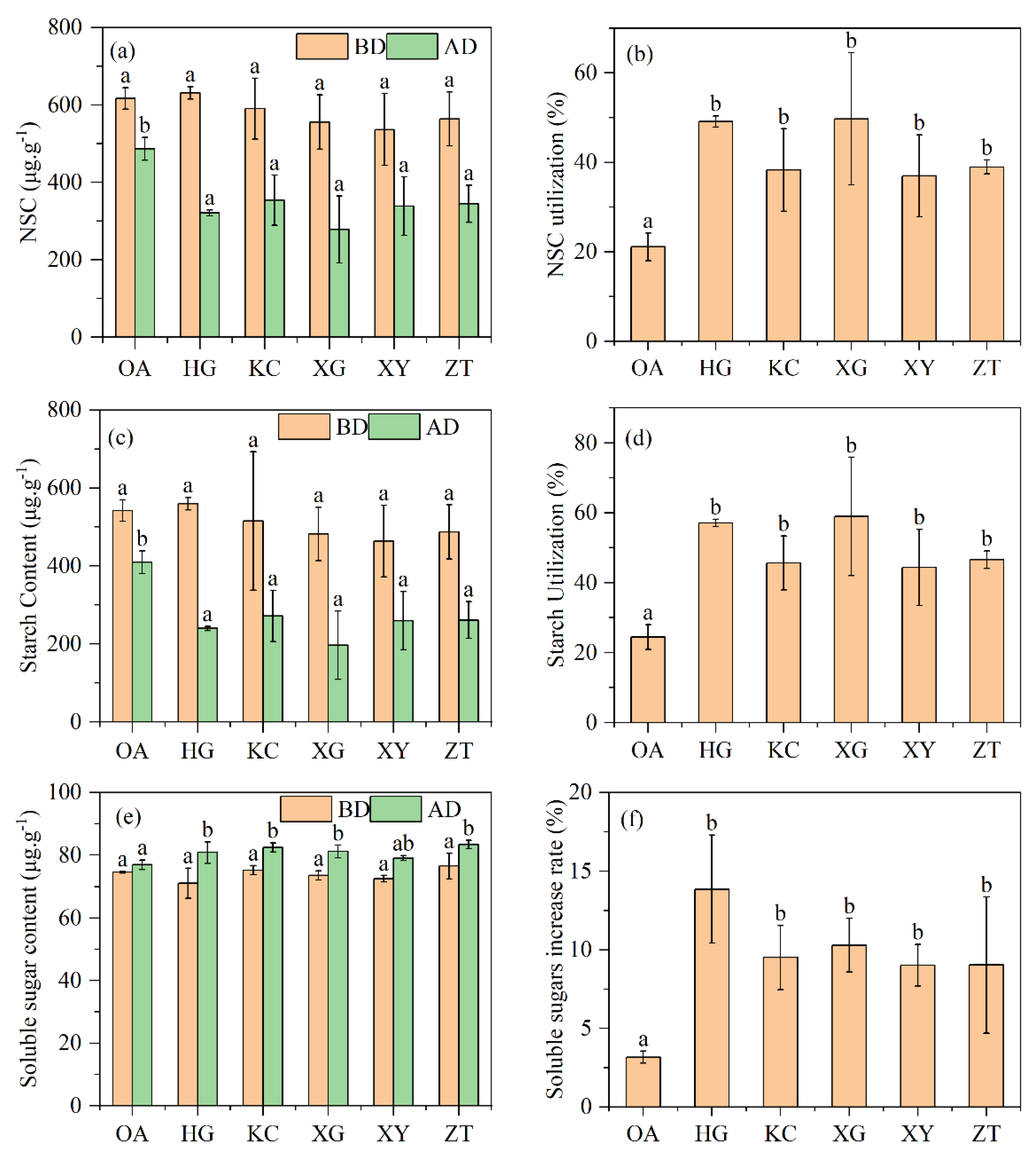
3.5. Non-Structural Carbohydrates
3.6. Correlation Analysis
4. Discussion
4.1. Relationships between Leaf Morphology and Photophysiological Characteristics and Agarwood Formation
4.2. Relationship between the Anatomical Characteristics of Xylem and Agarwood Formation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tan, C.S.; Isa, N.M.; Zainal, Z. Agarwood induction: Current developments and future perspectives. Front. Plant Sci. 2019, 10, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.P.; Mei, W.L.; Lin, Q.; Wang, H.; Wang, J.; Peng, S.Q.; Li, H.L.; Zhu, J.H.; Li, W.; Wang, P. Genome sequence of the agarwood tree Aquilaria sinensis (Lour.) Spreng: The first chromosome-level draft genome in the Thymelaeceae family. GigaScience 2020, 9, a13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Yang, J.; Jiang, C.; Zhou, J.H.; Zhao, Y.Y.; Huang, L.Q. Volatile organic compound and endogenous phytohormone characteristics during callus browning in Aquilaria sinensis. Ind. Crops Prod. 2021, 168, 113605. [Google Scholar] [CrossRef]
- Azren, P.D.; Lee, S.Y.; Emang, D.; Mohamed, R. History and perspectives of induction technology for agarwood production from cultivated Aquilaria in Asia: A review. J. Forestry Res. 2019, 30, 1–11. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.F.; Cui, Z.Y.; Xu, D.P. Morphological, physiological, biochemical and molecular analyses reveal wounding-induced agarwood formation mecha-nism in two types of Aquilaria sinensis (Lour.) Spreng. Ind. Crops Prod. 2022, 178, 114603. [Google Scholar] [CrossRef]
- Yu, M.; Liu, Y.Y.; Feng, J.; Chen, D.; Yang, Y.; Liu, P.W.; Yu, Z.X.; Wei, J.H. Remarkable Phytochemical Characteristics of Chi-Nan Agarwood Induced from New-Found Chi-Nan Germplasm of Aquilaria sinensis Compared with Ordinary Agarwood. Int. J. Anal. Chem. 2021, 10, 5593730. [Google Scholar] [CrossRef]
- Li, W.; Chen, H.Q.; Wang, H.; Mei, W.L.; Dai, H.F. Natural products in agarwood and Aquilaria plants: Chemistry, biological activities and biosynthesis. Nat. Prod. Rep. 2020, 38, 528–565. [Google Scholar] [CrossRef]
- Yang, L.; Yang, J.L.; Dong, W.H.; Wang, Y.L.; Zeng, J.; Yuan, J.Z.; Wang, H.; Mei, W.L.; Dai, H.F. The Characteristic fragrant sesquiterpenes and 2-(2-phenylethyl) chromones in wild and cultivated “Qi-Nan” agarwood. Molecules 2021, 26, 436. [Google Scholar] [CrossRef]
- Jumrani, K.; Bhatia, V.S.; Pandey, G.P. Impact of elevated temperatures on specific leaf weight, stomatal density, photosynthesis and chlorophyll fluorescence in soybean. Photosynth. Res. 2017, 131, 333–350. [Google Scholar] [CrossRef]
- Kanchan, J.; Virender, S.B. Identification of drought tolerant genotypes using physiological traits in soybean. Physiol. Mol. Biol. Plants 2019, 25, 697–711. [Google Scholar] [CrossRef]
- Olano, J.M.; Muñoz, N.G.; Arzac, A.; Rozas, V. Sex determines xylem anatomy in a dioecious conifer: Hydraulic consequences in a drier world. Tree Physiol. 2017, 11, 1493–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.X.; Qiao, M.J.; Fu, Y.L.; Wei, P.L.; Li, Y.J.; Liu, Z.G. Tissue structure changes of Aquilaria sinensis xylem after fungus induction. Forests 2022, 13, 43. [Google Scholar] [CrossRef]
- Nguyen, T.T.N.; Trinh, N.Y.; Le, P.K. Recovery yield and bioactivities evaluation on essential oil and ethanolic extract of star anise (Illicium verum Hook. f.). Chem. Eng. J. 2021, 83, 205–210. [Google Scholar] [CrossRef]
- Wang, Z.T.; Zhang, Z.; Tang, H.X.; Zhang, Q.; Lia, X.G.; Zhou, G.F. Genetic variation in leaf characters of F1 hybrids of Chinese Jujube. Sci. Hortic. 2019, 244, 372–378. [Google Scholar] [CrossRef]
- Abubakar, D.; Zahir, A.Z.; Muhammad, I.; Atif, M.; Atif, J.; Azhar, H.; Bushra Maqshoof, A. Efficacy of rhizobacterial exopolysaccharides in improving plant growth, physiology, and soil properties. Environ. Monit. Assess. 2012, 193, 515. [Google Scholar] [CrossRef]
- Yang, L.Z.; Pan, C.X.; Shao, S.L.; Tao, C.Y.; Wan, G.W.; Ying, Y.Q. Effect of PP333 and drought-stress on the activity photosynthetic characteristics and nonstructura1 carbohydrates of Phyllostachy sedulis seedings. Acta Ecol. Sinica 2018, 38, 2082–2091. [Google Scholar] [CrossRef]
- Johansen, D. Plant Microtechnique; McGrawHill: New York, NY, USA, 1940; pp. 55–60. [Google Scholar]
- Scholz, A.; Klepsch, M.; Karimi, Z.; Jansen, S. How to quantify conduits in wood? Front. Plant Sci. 2013, 4, 56. [Google Scholar] [CrossRef] [Green Version]
- AbdElgawad, H.; Avramova, V.; Baggerman, G.; Raemdonck, G.V.; Valkenborg, D.; Ostade, X.V.; Guisez, Y.; Prinsen, E.; Asard, H.; Ende, W.V.D.; et al. Starch biosynthesis contributes to the maintenance of photosynthesis and leaf growth under drought stress in maize. Plant Cell Environ. 2020, 43, 2254–2271. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Wang, C.K.; Wang, X.C. Spatial variability of non-structural carbohydrate concentrations in canopy branches and leaves of five temperate tree species. Acta Ecol. Sin. 2015, 35, 6496–6506. [Google Scholar] [CrossRef] [Green Version]
- Khadka, K.; Earl, H.J.; Raizada, M.N.; Navabi, A. A physio-morphological trait-based approach for breeding drought tolerant wheat. Front. Plant Sci. 2020, 11, 715. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, H.Y.; Wu, W.L.; Li, W.L. Effect of drought stress on physiological changes and leaf surface morphology in the blackberry, Braz. J. Bot. 2017, 40, 625–634. [Google Scholar] [CrossRef]
- Alves, A.A.C.; Setter, T.L. Response of cassava leaf area expansion to water deficit: Cell proliferation, cell expansion and delayed development. Ann. Bot. 2004, 94, 605–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.Q.; Wei, J.H.; Yang, J.S.; Zhang, Z.; Yang, Y.; Gao, Z.H.; Sui, C.; Gong, B. Chemical constituents of agarwood originating from the endemic genus Aquilaria plants. Chem. Biodivers. 2012, 9, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, N.P.; Hou, J.H.; Xu, L.; Liu, C.C.; Zhang, J.H.; Wang, Q.F.; Zhang, X.M.; Wu, X.Q. Factors influencing leaf chlorophyll content in natural forests at the biome scale. Front. Ecol. Evol. 2018, 6, 64. [Google Scholar] [CrossRef] [Green Version]
- Liang, L.; Di, L.P.; Huang, T.; Wang, J.H.; Lin, L.; Wang, L.J.; Yang, M.H. Estimation of leaf nitrogen content in wheat using new hyperspectral indices and a random forest regression algorithm. Remote Sens. 2018, 10, 1940. [Google Scholar] [CrossRef] [Green Version]
- Jia, M.; Colombo, R.; Rossini, M.; Celesti, M. Estimation of leaf nitrogen content and photosynthetic nitrogen use efficiency in wheat using sun-induced chlorophyll fluorescence at the leaf and canopy scales. Eur. J. Agron. 2021, 122, 126192. [Google Scholar] [CrossRef]
- Nobuchi, T.; Somkid, S. Preliminary Observation of Aquliaria crassna Wood Associated with the Formation of Aloeswood Bult. Kyoto Univ. For. 1991, 63, 226–235. Available online: http://hdl.handle.net/2433/191992 (accessed on 10 May 2022).
- Kono, Y.; Ishida, A.; Saiki, S.T.; Yoshimura, K.; Dannoura, M.; Yazaki, K.; Kimura, F.; Yoshimura, J.; Aikawa, S. Initial hydraulic failure followed by late-stage carbon starvation leads to drought-induced death in the tree Trema orientalis. Commun. Biol. 2019, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, H.; Trumbore, S. Understanding the roles of nonstructural carbohydrates in forest trees—From what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.H.; Liang, J.; Tang, G.M.; Wang, X.F.; Liu, F.C.; Zhao, D.C. Drought stress affects on growth, water use efficiency, gas exchange and chlorophyll fluorescence of Juglans rootstocks. Sci. Hortic. 2019, 250, 230–235. [Google Scholar] [CrossRef]
- Adams, S.J.; Manohara, T.N.; Krishnamurthy, K.V.; Kumar, T.S. Hito-Chemical Studies on Fungal-Induced Agarwood. Indian J. Plant Sci. 2016, 5, 102–110. Available online: https://www.researchgate.net/publication/300056960 (accessed on 12 July 2021).
- Naoki, T.; Tatsuya, A.; Miyuki, T.N.; Yuzou, S.; Shingo, S.; Nobutaka, M.; Toru, T. Populus NST/SND orthologs are key regulators of secondary cell wall formation in wood fibers, phloem fibers and xylem ray parenchyma cells. Tree Physiol. 2019, 39, 514–525. [Google Scholar] [CrossRef]
- Rubin, E.M. Genomics of cellulosic biofuels. Nature 2008, 454, 841–845. [Google Scholar] [CrossRef] [PubMed]
- Studer, M.H.; DeMartini, J.D.; Davis, M.F.; Sykes, R.W.; Davison, B.; Keller, M.; Tuskan, G.A.; Wyman, C.E. Lignin content in natural Populus variants affects sugar release. Proc. Natl. Acad. Sci. USA 2011, 108, 6300–6305. [Google Scholar] [CrossRef] [Green Version]
- Rao, K.R.; Dayal, R. The secondary xylem of Aquilaria agallocha (Thymelaeaceae) and the formation of ‘agar’. IAWA J. 1992, 13, 163–172. [Google Scholar] [CrossRef]
- Angyalossy, V.; Pace, M.R.; Lima, A.C. Liana Anatomy: A Broad Perspective on Structural Evolution of the Vascular System; Wiley-Blackwell Publishers: Oxford, UK, 2015; pp. 253–287. [Google Scholar] [CrossRef]
- Carlquist, S. Interxylary phloem: Diversity and functions. Brittonia 2013, 62, 477–495. [Google Scholar] [CrossRef]
- Liu, P.W.; Zhang, X.L.; Yang, Y.; Sui, C.; Xu, Y.H.; Wei, J.H. Interxylary phloem and xylem rays are the structural foundation of agarwood resin formation in the stems of Aquilaria sinensis. Trees 2019, 33, 533–542. [Google Scholar] [CrossRef]
- Cui, Z.Y.; Li, X.F.; Xu, D.P.; Yang, Z.J.; Zhang, N.N.; Liu, X.J.; Hong, Z. Physiological changes during heartwood formation induced by plant growth regulators in Dalbergia odorifera (Leguminosae). IAWA J. 2021, 42, 217–234. [Google Scholar] [CrossRef]
- Bacete, L.; Mélida, H.; Miedes, E.; Molina, A. Plant cell wall-mediated immunity: Cell wall changes trigger disease resistance responses. Plant J. 2018, 93, 614–636. [Google Scholar] [CrossRef]
- De Lorenzo, G.; Ferrarim, S.; Cervone, F.; Okun, E. Extracellular DAMPs in Plants and Mammals: Immunity, Tissue Damage and Repair. Trends Immunol. 2018, 39, 937–950. [Google Scholar] [CrossRef]
- Molina, A.; Miedesa, E.; Bacete, L.; Rodríguez, T.; Mélida, H.; Denancé, N.; Sánchez, V.A.; Rivière, M.P.; López, G.; Freydier, A. Arabidopsis cell wall composition determines disease resistance specificity and fitness. Proc. Natl. Acad. Sci. USA 2020, 118, e2010243118. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.A.S.; Li, S.D.; Lin, A.Z.; Boutrot, F.; Grossmann, G.; Zipfel, C.; Somerville, S.C. Cellulose-derived oligomers act as damage-associated molecular patterns and trigger defense-like responses. Plant Physiol. 2017, 173, 2383–2398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelsdorf, T.; Kjaer, L.; Bisceglia, N.G.; Vaahtera, L.; Bauer, S.; Miedes, E.; Wormit, A.; James, L.; Chairam, I.; Molina, A. Functional characterization of genes mediating cell wall metabolism and responses to plant cell wall integrity impairment. BMC Plant Biol. 2019, 19, 320. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aquilaria sinensis | OA | HG | KC | XG | XY | ZT |
|---|---|---|---|---|---|---|
| Tree height (m) | 3.06 ± 0.26 | 3.09 ± 0.47 | 3.32 ± 0.28 | 3.57 ± 0.54 | 3.13 ± 0.42 | 3.58 ± 0.64 |
| Base diameter at 20 cm above ground (cm) | 5.63 ± 0.44 | 5.49 ± 0.52 | 5.76 ± 0.33 | 6.00 ± 0.35 | 5.52 ± 0.49 | 5.80 ± 0.35 |
| Aquilaria sinensis | TNV | NSV | NVP | VWT | VD | VA | TAV |
|---|---|---|---|---|---|---|---|
| OA | 23.00 ± 1.00a | 4.67 ± 2.08a | 7.33 ± 1.53ab | 9.61 ± 2.18d | 51.40 ± 6.46bcd | 2630.74 ± 393.35c | 60,886.58 ± 6576.12b |
| HG | 30.33 ± 5.69a | 6.67 ± 2.52a | 8.67 ± 1.15ab | 5.11 ± 0.93a | 59.22 ± 2.77d | 3629.67 ± 373.92d | 116,443.47 ± 10,983.00d |
| KC | 29.33 ± 11.93a | 6.33 ± 1.53a | 7.00 ± 3.61ab | 6.00 ± 1.89b | 51.88 ± 5.50cd | 3110.24 ± 432.15c | 96,219.33 ± 4612.71c |
| XG | 19.67 ± 0.58a | 5.00 ± 1.00a | 5.67 ± 0.58a | 7.17 ± 1.69c | 42.48 ± 2.84ab | 2193.02 ± 296.69ab | 44,511.03 ± 5202.55a |
| XY | 44.67 ± 8.33b | 6.33 ± 1.53a | 10.33 ±0.58b | 7.28 ± 1.35c | 44.72 ± 6.93abc | 2439.53 ± 503.52b | 122,569.73 ± 7665.42d |
| ZT | 60.67 ± 6.51c | 8.00 ± 2.65a | 17.00 ±2.65c | 5.10 ± 1.08a | 40.45 ± 2.69a | 1841.65 ± 277.38a | 110,664.33 ± 7402.35d |
| Aquilaria sinensis | RH | RW | RD | RCL | RCW | RCWT | IPA |
| OA | 337.43 ± 141.20cd | 20.98 ± 3.60b | 23.43 ±3.36ab | 34.69 ± 1.23a | 25.91 ± 1.32a | 4.83 ± 0.05a | 71,592.19 ± 21,859.73a |
| HG | 242.13 ± 100.15a | 17.34 ± 4.37a | 23.33 ±1.51ab | 37.74 ± 3.12a | 22.54 ± 0.44a | 5.72 ± 0.33b | 200,308.43 ± 36,001.94c |
| KC | 364.84 ± 118.00d | 30.70 ± 9.01d | 22.83 ±2.14ab | 36.90 ± 3.48a | 25.05 ± 1.03a | 5.46 ± 0.34b | 111,693.37 ± 18,575.93b |
| XG | 303.49 ± 93.59bc | 25.86 ± 6.65c | 21.83 ± 1.17a | 35.27 ± 3.10a | 23.58 ± 0.45a | 6.71 ± 0.52c | 105,941.93 ± 6539.21b |
| XY | 288.38 ± 87.66ab | 26.40 ± 7.87c | 25.00 ± 1.41b | 36.24 ± 4.64a | 27.02 ± 0.64a | 5.50 ± 0.27b | 108,637.07 ± 4499.48b |
| ZT | 265.20 ± 96.79ab | 18.89 ± 3.66ab | 33.17 ± 2.64c | 35.01 ± 2.88a | 24.27 ± 1.56a | 5.41 ± 0.21b | 140,071.33 ± 14,646.67b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Cui, Z.; Liu, X.; Hong, Z.; Zhang, P.; Xu, D. Comparative Morphological, Anatomical and Physiological Analyses Explain the Difference of Wounding-Induced Agarwood Formation between Ordinary Agarwood Nongrafted Plants and Five Grafted Qi-Nan Clones (Aquilaria sinensis). Forests 2022, 13, 1618. https://doi.org/10.3390/f13101618
Li X, Cui Z, Liu X, Hong Z, Zhang P, Xu D. Comparative Morphological, Anatomical and Physiological Analyses Explain the Difference of Wounding-Induced Agarwood Formation between Ordinary Agarwood Nongrafted Plants and Five Grafted Qi-Nan Clones (Aquilaria sinensis). Forests. 2022; 13(10):1618. https://doi.org/10.3390/f13101618
Chicago/Turabian StyleLi, Xiaofei, Zhiyi Cui, Xiaojin Liu, Zhou Hong, Peng Zhang, and Daping Xu. 2022. "Comparative Morphological, Anatomical and Physiological Analyses Explain the Difference of Wounding-Induced Agarwood Formation between Ordinary Agarwood Nongrafted Plants and Five Grafted Qi-Nan Clones (Aquilaria sinensis)" Forests 13, no. 10: 1618. https://doi.org/10.3390/f13101618