Relationships between Soil Nematode Communities and Soil Quality as Affected by Land-Use Type



 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Region and Experimental Design
2.2. Soil Sampling and Analyses
2.3. Data Analysis
3. Results
3.1. Soil Nematode Community
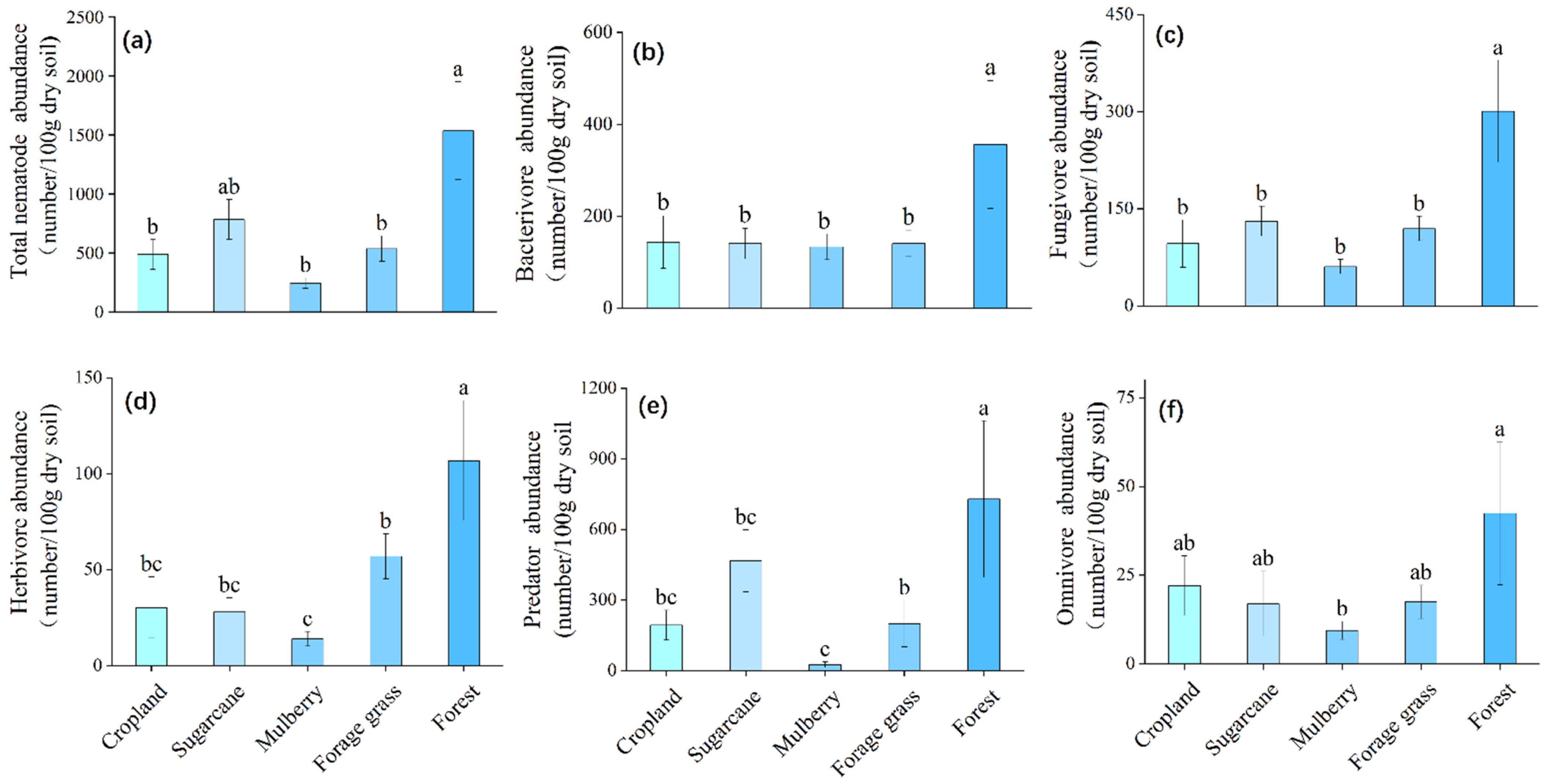
3.1.1. Total Number of Soil Nematodes and Trophic Groups
3.1.2. Ecological Index and Metabolic Footprint of The Soil Nematode Communities
3.1.3. Relationship between Soil Nematode Communities and Soil Physicochemical Properties
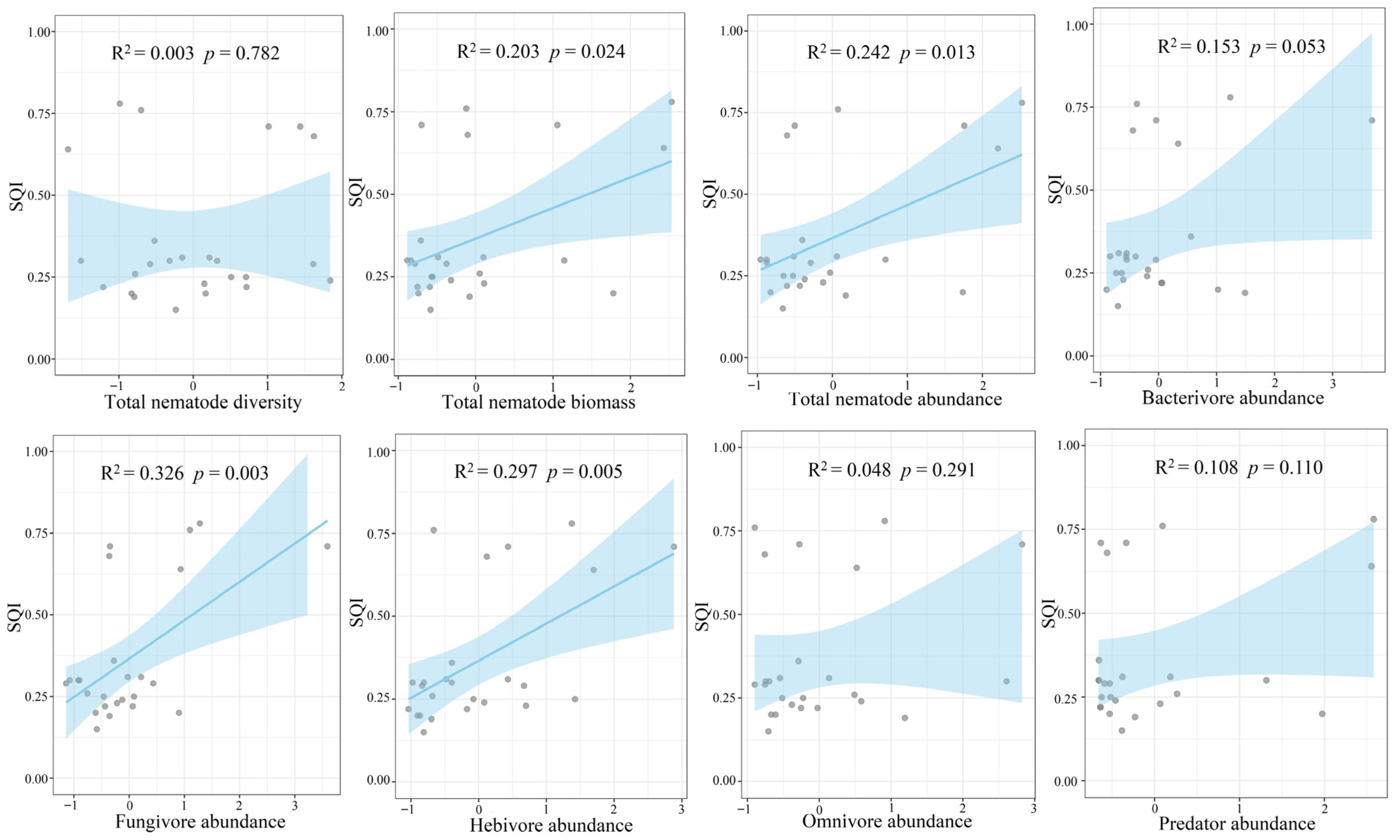
3.2. Relationship between Nematode Properties and Soil Quality
4. Discussion
4.1. Effects of Land-Use Type on Soil Nematode Communities
4.2. Relationship between Nematodes and Soil Quality
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andrews, S.S.; Flora, C.B.; Mitchell, J.P.; Karlen, D.L. Growers’ perceptions and acceptance of soil quality indices. Geoderma 2003, 114, 187–213. [Google Scholar] [CrossRef]
- Andrews, S.S.; Karlen, D.L.; Mbardella, C.C. The Soil Management Assessment Framework. Soil Sci. Soc. Am. J. 2004, 68, 1945. [Google Scholar] [CrossRef]
- Baker, J.M.; Ochsner, T.E.; Venterea, R.T.; Griffis, T.J. Tillage and soil carbon sequestration—What do we really know? Agric. Ecosyst. Environ. 2007, 118, 1–5. [Google Scholar] [CrossRef]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Barker, K.R. Nematode Extraction and Bioassays. Adv. Treatise Meloidogyne 1985, 2, 19–35. [Google Scholar]
- Barrios, E.; Delve, R.J.; Bekunda, M.; Mowo, J.; Agunda, J.; Ramisch, J.; Trejo, M.T.; Thomas, R.J. Indicators of soil quality: A South–South development of a methodological guide for linking local and technical knowledge. Geoderma 2006, 135, 248–259. [Google Scholar] [CrossRef]
- Berkelmans, R.; Ferris, H.; Tenuta, M.; van Bruggen, A.H.C. Effects of long-term crop management on nematode trophic levels other than plant feeders disappear after 1 year of disruptive soil management. Appl. Soil Ecol. 2003, 23, 223–235. [Google Scholar] [CrossRef]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Bongers, T.; Ferris, H. Nematode community structure as a bioindicator in environmental monitoring. Trends Ecol. Evol. 1999, 14, 224–228. [Google Scholar] [CrossRef]
- Bünemann, E.K.; Bongiorno, G.; Bai, Z.; Creamer, R.E.; De Deyn, G.; de Goede, R.; Fleskens, L.; Geissen, V.; Kuyper, T.W.; Mäder, P.; et al. Soil quality—A critical review. Soil Biol. Biochem. 2018, 120, 105–125. [Google Scholar] [CrossRef]
- Carter, M.; Gregorich, E. Soil Sampling and Methods of Analysis; Lewis Publishers: Boca Raton FL, USA, 2006; pp. 481–483. [Google Scholar]
- Ditzler, C.A.; Tugel, A.J. Soil Quality Field Tools. Agron. J. 2002, 94, 33–38. [Google Scholar]
- Doran, J.W. The Dynamics of Soil Quality as a Measure of Sustainable Management; American Society of Agronomy: Madison, WI, USA, 1994. [Google Scholar]
- Doran, J.W.; Parkin, T.B. Defining and Assessing Soil Quality. In Defining Soil Quality for a Sustainable Environment; Doran, J.W., Coleman, D.C., Bezdicek, D.F., Stewart, B.A., Eds.; SSSA Special Publications: Madison, WI, USA, 1994; pp. 3–21. [Google Scholar]
- Doran, J.W.; Parkin, T.B. Quantitative Indicators of Soil Quality, a Minimum Data Set; Doran, J.W., Jones, A.J., Eds.; Methods for Assessing Soil Quality, Special Publication No. 49; Soil Science Society of America: Madison, WI, USA, 1996; pp. 25–37. [Google Scholar]
- Dumanski, J.; Pieri, C. Land quality indicators: Research plan. Agric. Ecosyst. Environ. 2000, 81, 93–102. [Google Scholar] [CrossRef]
- Elliott, E.T.; Pankhurst, C.E.; Doube, B.M.; Gupta, V.; Grace, P.R. The Potential Use of Soil Biotic Activity as an Indicator of Productivity, Sustainability, and Pollution; CSIRO Publications: Melbourne, Australia, 1994. [Google Scholar]
- Ferris, H.; Bongers, T.; de Goede, R.G.M. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Frostegård, A.; Bååth, E. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol. Fertil. Soils 1996, 22, 59–65. [Google Scholar] [CrossRef]
- Fu, B.J.; Liu, S.L.; Lu, Y.H.; Chen, L.D.; Ma, K.M.; Liu, G.H. Comparing the soil quality changes of different land uses determined by two quantitative methods. J. Environ. Sci. 2003, 15, 167–172. [Google Scholar]
- Fu, S.L.; Coleman, D.C.; Hendrix, P.F.; Crossley, D.A. Responses of trophic groups of soil nematodes to residue application under conventional tillage and no-till regimes. Soil Biol. Biochem. 2000, 32, 1731–1741. [Google Scholar] [CrossRef]
- Gao, D.; Wang, F.; Li, J.; Yu, S.; Li, Z.; Zhao, J. Soil nematode communities as indicators of soil health in different land use types in tropical area. Nematology 2020, 22, 595–610. [Google Scholar] [CrossRef]
- Gao, X.; Wu, P.; Zhao, X.; Wang, J.; Shi, Y. Effects of land use on soil moisture variations in a semi-arid catchment: Implications for land and agricultural water management. Land Degrad. Dev. 2014, 25, 163–172. [Google Scholar] [CrossRef]
- Gaston, K.J. Biodiversity and extinction: The importance of being common. Prog. Phys. Geogr. Earth Environ. 2008, 32, 73–79. [Google Scholar] [CrossRef]
- Gong, W.Z. Assessment and analysis of soil quality changes after eleven years of reclamation in subtropical China. Geoderma 1998, 81, 339–355. [Google Scholar]
- Guan, P.; Zhang, X.; Yu, J.; Cheng, Y.; Li, Q.; Andriuzzi, W.S.; Liang, W. Soil microbial food web channels associated with biological soil crusts in desertification restoration: The carbon flow from microbes to nematodes. Soil Biol. Biochem. 2018, 116, 82–90. [Google Scholar] [CrossRef]
- Gudelj, I.; Weitz, J.S.; Ferenci, T.; Claire Horner-Devine, M.; Marx, C.J.; Meyer, J.R.; Forde, S.E. An integrative approach to understanding microbial diversity: From intracellular mechanisms to community structure. Ecol. Lett. 2010, 13, 1073–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingham, R.E.; Trofymow, J.A.; Ingham, E.R.; Coleman, D.C. Interactions of bacteria, fungi, and their nematode grazers—Effects on nutrient cycling and plant-growth. Ecol. Monogr. 1985, 55, 119–140. [Google Scholar] [CrossRef]
- Larson, W.E.; Pierce, F. Conservation and Enhancement of Soil Quality. Evaluation for Sustainable Land Management in the Developing World; Dumanski, J., Pushparajah, E., Latham, M., Myers, R., Eds.; Board for Soil Research and Management: Bangkok, Thailand, 1991; pp. 175–203. [Google Scholar]
- Li, D.; Liu, J.; Chen, H.; Zheng, L.; Wang, K. Soil gross nitrogen transformations in responses to land use conversion in a subtropical karst region. J. Environ. Manag. 2018, 212, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, J.; Chen, H.; Zheng, L.; Wang, K. Soil microbial community responses to forage grass cultivation in degraded karst soils, Southwest China. Land Degrad. Dev. 2018, 29, 4262–4270. [Google Scholar] [CrossRef]
- Li, D.J.; Wen, L.; Yang, L.Q.; Luo, P.; Xiao, K.C.; Chen, H.; Zhang, W.; He, X.Y.; Chen, H.S.; Wang, K.L. Dynamics of soil organic carbon and nitrogen following agricultural abandonment in a karst region. J. Geophys. Res. Biogeosciences 2017, 122, 230–242. [Google Scholar] [CrossRef]
- Li, Q.; Liang, W.; Ou, W. Response of nematode community to land use pattern in aquic brown soil. Biodivers. Sci. 2007, 15, 8. [Google Scholar]
- Liu, J.; Wu, L.; Chen, D.; Li, M.; Wei, C. Soil quality assessment of different Camellia oleifera stands in mid-subtropical China. Appl. Soil Ecol. 2017, 113, 29–35. [Google Scholar] [CrossRef]
- Liu, L.; Gundersen, P.; Zhang, T.; Mo, J. Effects of phosphorus addition on soil microbial biomass and community composition in three forest types in tropical China. Soil Biol. Biochem. 2012, 44, 31–38. [Google Scholar] [CrossRef]
- Mamehpour, N.; Rezapour, S.; Ghaemian, N. Quantitative assessment of soil quality indices for urban croplands in a calcareous semi-arid ecosystem. Geoderma 2021, 382, 114781. [Google Scholar] [CrossRef]
- Masto, R.E.; Chhonkar, P.K.; Singh, D.; Patra, A.K. Alternative soil quality indices for evaluating the effect of intensive cropping, fertilisation and manuring for 31 years in the semi-arid soils of India. Environ. Monit. Assess. 2008, 136, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Mcmahon, S.K.; Williams, M.A.; Bottomley, P.J.; Myrold, D.D. Dynamics of Microbial Communities during Decomposition of Carbon-13 Labeled Ryegrass Fractions in Soil. Soil Sci. Soc. Am. J. 2005, 69, 1238–1247. [Google Scholar] [CrossRef] [Green Version]
- Neher, D.A. Role of nematodes in soil health and their use as indicators. J. Nematol. 2001, 33, 161–168. [Google Scholar] [PubMed]
- Neher, D.A.; Campbell, C.L. Nematode communities and microbial biomass in soils with annual and perennial crops. Appl. Soil Ecol. 1994, 1, 17–28. [Google Scholar] [CrossRef]
- Neher, D.A.; Wu, J.; Barbercheck, M.E.; Anas, O. Ecosystem type affects interpretation of soil nematode community measures. Appl. Soil Ecol. 2005, 30, 47–64. [Google Scholar] [CrossRef]
- Ngo-Mbogba, M.; Yemefack, M.; Nyeck, B. Assessing soil quality under different land cover types within shifting agriculture in South Cameroon. Soil Tillage Res. 2015, 150, 124–131. [Google Scholar] [CrossRef]
- Okada, H.; Kadota, I. Host status of 10 fungal isolates for two nematode species, Filenchus misellus and Aphelenchus avenae. Soil Biol. Biochem. 2003, 35, 1601–1607. [Google Scholar] [CrossRef]
- Olsson, P.A. Signature fatty acids provide tools for determination of distribution and interactions of mycorrhizal fungi in soil. FEMS Microbiol. Ecol. 2006, 29, 303–310. [Google Scholar] [CrossRef]
- Ou, W.; Liang, W.; Jiang, Y.; Li, Q.; Wen, D. Vertical distribution of soil nematodes under different land use types in an aquic brown soil. Pedobiologia 2005, 49, 139–148. [Google Scholar] [CrossRef]
- Paz-Ferreiro, J.; Fu, S. Biological Indices for Soil Quality Evaluation: Perspectives and Limitations. Land Degrad. Dev. 2013, 27, 14–25. [Google Scholar] [CrossRef]
- Porazinska, D.L.; Duncan, L.W.; McSorley, R.; Graham, J.H. Nematode communities as indicators of status and processes of a soil ecosystem influenced by agricultural management practices. Appl. Soil Ecol. 1999, 13, 69–86. [Google Scholar] [CrossRef]
- Quist, C.W.; Schrama, M.; de Haan, J.J.; Smant, G.; Bakker, J.; van der Putten, W.H.; Helder, J. Organic farming practices result in compositional shifts in nematode communities that exceed crop-related changes. Appl. Soil Ecol. 2016, 98, 254–260. [Google Scholar] [CrossRef]
- Röty, M.; Huhta, V. Earthworms and pH affect communities of nematodes and enchytraeids in forest soil. Biol. Fertil. Soils 2003, 38, 52–58. [Google Scholar] [CrossRef]
- Smith, J.L.; Halvorson, J.J.; Papendick, R.I. Using Multiple-Variable Indicator Kriging for Evaluating Soil Quality. Soil Sci. Soc. Am. J. 1993, 57, 743–749. [Google Scholar] [CrossRef]
- van den Hoogen, J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.A.; de Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.S.; et al. Soil nematode abundance and functional group composition at a global scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Viketoft, M. Determinants of small-scale spatial patterns: Importance of space, plants and abiotics for soil nematodes. Soil Biol. Biochem. 2013, 62, 92–98. [Google Scholar] [CrossRef]
- Wall, J.; Skene, K.; Neilson, R. Nematode community and trophic structure along a sand dune succession. Biol. Fertil. Soils 2002, 35, 293–301. [Google Scholar] [CrossRef]
- Wang, K.L.; Yue, Y.M.; Chen, H.S.; Wu, X.B.; Xiao, J.; Qi, X.K.; Zhang, W.; Du, H. The comprehensive treatment of karst rocky desertification and its regional restoration effects. Acta Ecol. Sin. 2019, 39, 7432–7440. [Google Scholar]
- Wardle, D.A. Impacts of Disturbance on Detritus Food Webs in Agro-Ecosystems of Contrasting Tillage and Weed Management Practices. Adv. Ecol. Res. 1995, 26, 105–185. [Google Scholar]
- Wen, L.; Li, D.; Yang, L.; Luo, P.; Chen, H.; Xiao, K.; Song, T.; Zhang, W.; He, X.; Chen, H.; et al. Rapid recuperation of soil nitrogen following agricultural abandonment in a karst area, southwest China. Biogeochemistry 2016, 129, 341–354. [Google Scholar] [CrossRef]
- Yang, L.; Luo, P.; Wen, L.; Li, D. Soil organic carbon accumulation during post-agricultural succession in a karst area, southwest China. Sci. Rep. 2016, 6, 37118. [Google Scholar] [CrossRef] [PubMed]
- Yeates, G.W. Populations of nematode genera in soils under pasture. N. Z. J. Agric. Res. 1978, 21, 321–330. [Google Scholar] [CrossRef]
- Yeates, G.W. Soil nematodes in terrestrial ecosystems. J. Nematol. 1979, 11, 213–229. [Google Scholar] [PubMed]
- Yeates, G.W.; Bongers, T. Nematode diversity in agroecosystems. Agric. Ecosyst. Environ. 1999, 74, 113–135. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T.; Freckman, R.G.D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera-an outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Zhang, C.; Liu, G.B.; Xue, S.; Wang, G.L. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of abandoned farmland in the Loess Plateau. Soil Biol. Biochem. 2016, 97, 40–49. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, S.; Hu, S.; Mao, L.; Han, R. Discussion on Fertilization Techniques of Mulberry in Summer and Autumn. Agric. Technol. Equip. 2021, 1, 135–136. [Google Scholar]
- Zhao, J.; Li, S.; He, X.; Liu, L.; Wang, K. The soil biota composition along a progressive succession of secondary vegetation in a karst area. PLoS ONE 2014, 9, e112436. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Neher, D.A. Soil energy pathways of different ecosystems using nematode trophic group analysis: A meta analysis. Nematology 2014, 16, 379–385. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, K. Methods for cleaning turbid nematode suspensions collected from different land-use types and soil types. Soil Ecol. Lett. 2021, 4, 429–434. [Google Scholar] [CrossRef]
- Zhao, J.; Zeng, Z.; He, X.; Chen, H.; Wang, K. Effects of monoculture and mixed culture of grass and legume forage species on soil microbial community structure under different levels of nitrogen fertilization. Eur. J. Soil Biol. 2015, 68, 61–68. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, W.; Wang, K.; Song, T.; Du, H. Responses of the soil nematode community to management of hybrid napiergrass: The trade-off between positive and negative effects. Appl. Soil Ecol. 2014, 75, 134–144. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cropland | Sugarcane | Mulberry | Forage Grass | Forest | |
|---|---|---|---|---|---|
| EI | 44.75 ± 1.92 | 41.81 ± 7.6 | 38.41 ± 2.94 | 43.74 ± 4.87 | 41.71 ± 1.94 |
| SI | 85.05 ± 5.14 a | 82.19 ± 19.34 ab | 60.71 ± 15.27 b | 63.27 ± 24.39 ab | 83.19 ± 10.54 ab |
| NCR | 0.58 ± 0.10 | 0.57 ± 0.11 | 0.7 ± 0.09 | 0.53 ± 0.08 | 0.51 ± 0.05 |
| H’ | 2.22 ± 0.12 | 2.09 ± 0.13 | 2.37 ± 0.21 | 2.55 ± 0.17 | 2.28 ± 0.28 |
| SR | 2.98 ± 0.32 | 2.6 ± 0.37 | 3.2 ± 0.44 | 3.68 ± 0.38 | 3.35 ± 0.26 |
| λ | 0.16 ± 0.02 | 0.2 ± 0.03 | 0.15 ± 0.04 | 0.14 ± 0.03 | 0.24 ± 0.08 |
| MI | 3.06 ± 0.1 ab | 3.15 ± 0.27 a | 2.49 ± 0.1 b | 2.65 ± 0.23 ab | 3.11 ± 0.18 a |
| PPI | 3.53 ± 0.26 | 2.66 ± 0.35 | 3.56 ± 0.20 | 3.33 ± 0.38 | 3.25 ± 0.41 |
| Footprint | 55.46 ± 11.57 b | 101.52 ± 33.28 ab | 35.71 ± 10.01 b | 60.94 ± 15.22 b | 163.87 ± 33.15 a |
| BD b | SWC | SOC | TN | C:N | pH | Ca | Mg | AN | NN | |
|---|---|---|---|---|---|---|---|---|---|---|
| Total a | −0.480 * | 0.299 | 0.579 ** | 0.496 * | 0.615 ** | 0.394 | 0.22 | 0.710 ** | −0.175 | 0.379 |
| Ba | −0.364 | 0.26 | 0.430 * | 0.366 | 0.470 * | 0.193 | 0.144 | 0.487 * | 0.013 | 0.213 |
| Fu | −0.523 ** | 0.408 * | 0.596 ** | 0.521 ** | 0.623 ** | 0.189 | 0.089 | 0.680 ** | −0.309 | 0.500 * |
| He | −0.573 ** | 0.439 * | 0.492 * | 0.483 * | 0.460 * | 0.319 | 0.261 | 0.530 ** | −0.281 | 0.328 |
| Pr | −0.332 | 0.165 | 0.435 * | 0.364 | 0.467 * | 0.384 | 0.196 | 0.570 ** | −0.152 | 0.29 |
| Om | −0.219 | 0.125 | 0.246 | 0.23 | 0.266 | 0.3 | 0.172 | 0.298 | 0.121 | 0.024 |
| EI | 0.109 | −0.161 | −0.188 | −0.252 | −0.167 | −0.509 ** | −0.577 ** | −0.101 | −0.046 | 0.201 |
| SI | −0.052 | −0.055 | 0.123 | 0.025 | 0.178 | 0.265 | −0.081 | 0.269 | 0.062 | 0.027 |
| NCR | 0.142 | −0.181 | −0.082 | −0.038 | −0.061 | 0.273 | 0.322 | −0.123 | 0.218 | −0.305 |
| H’ | −0.019 | 0.159 | −0.049 | −0.033 | −0.064 | −0.098 | −0.015 | −0.188 | −0.106 | −0.023 |
| SR | −0.147 | 0.232 | 0.049 | 0.05 | 0.023 | −0.093 | −0.06 | −0.013 | −0.135 | 0.031 |
| λ | −0.262 | 0.077 | 0.303 | 0.279 | 0.308 | 0.256 | 0.165 | 0.442 * | −0.005 | 0.183 |
| MI | −0.121 | −0.017 | 0.163 | 0.073 | 0.221 | 0.359 | 0.017 | 0.297 | 0.064 | 0.017 |
| PPI | 0.108 | 0.077 | −0.094 | −0.101 | −0.11 | −0.143 | −0.084 | −0.146 | 0.1 | −0.192 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Chen, X.; Li, J.; Liao, X.; Li, D.; He, X.; Zhang, W.; Zhao, J. Relationships between Soil Nematode Communities and Soil Quality as Affected by Land-Use Type. Forests 2022, 13, 1658. https://doi.org/10.3390/f13101658
Li Z, Chen X, Li J, Liao X, Li D, He X, Zhang W, Zhao J. Relationships between Soil Nematode Communities and Soil Quality as Affected by Land-Use Type. Forests. 2022; 13(10):1658. https://doi.org/10.3390/f13101658
Chicago/Turabian StyleLi, Zhilei, Xiaomei Chen, Jiangnan Li, Xionghui Liao, Dejun Li, Xunyang He, Wei Zhang, and Jie Zhao. 2022. "Relationships between Soil Nematode Communities and Soil Quality as Affected by Land-Use Type" Forests 13, no. 10: 1658. https://doi.org/10.3390/f13101658
APA StyleLi, Z., Chen, X., Li, J., Liao, X., Li, D., He, X., Zhang, W., & Zhao, J. (2022). Relationships between Soil Nematode Communities and Soil Quality as Affected by Land-Use Type. Forests, 13(10), 1658. https://doi.org/10.3390/f13101658