
Understanding Species Diversity, Phenology and Environmental Implications of Different Life Forms in Coniferous Forests: A Case Study from Bhallesa Hills of Pir Panjal Mountain, Western Himalaya, India



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.2.1. Plant Surveys, Collection and Identification
2.2.2. Presentation of Data
2.3. Data Analysis
2.4. Literature Sources
3. Results and Discussion
3.1. Floristic Composition and Diversity of Species and Families
3.2. Species Diversity in Different Growth Forms
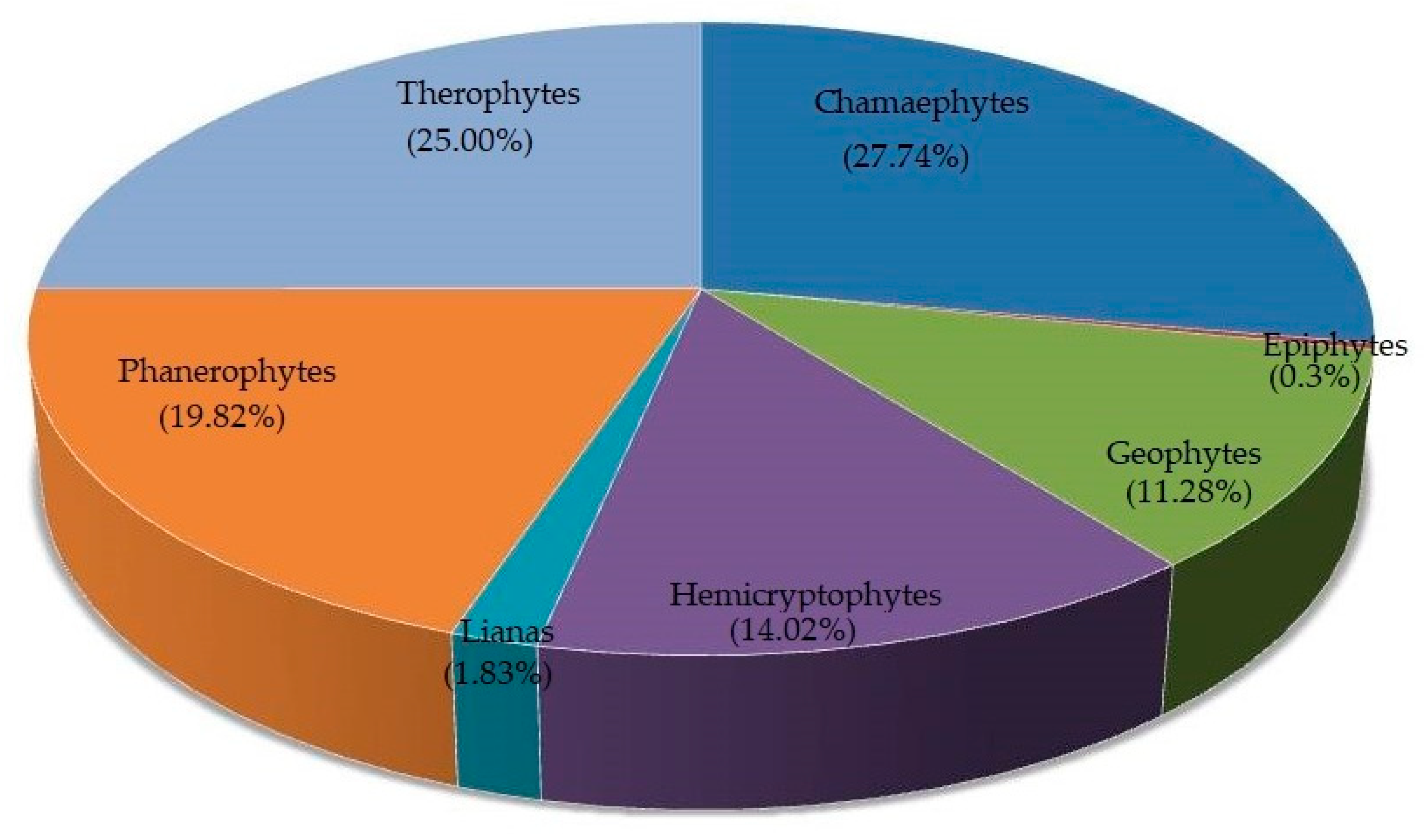
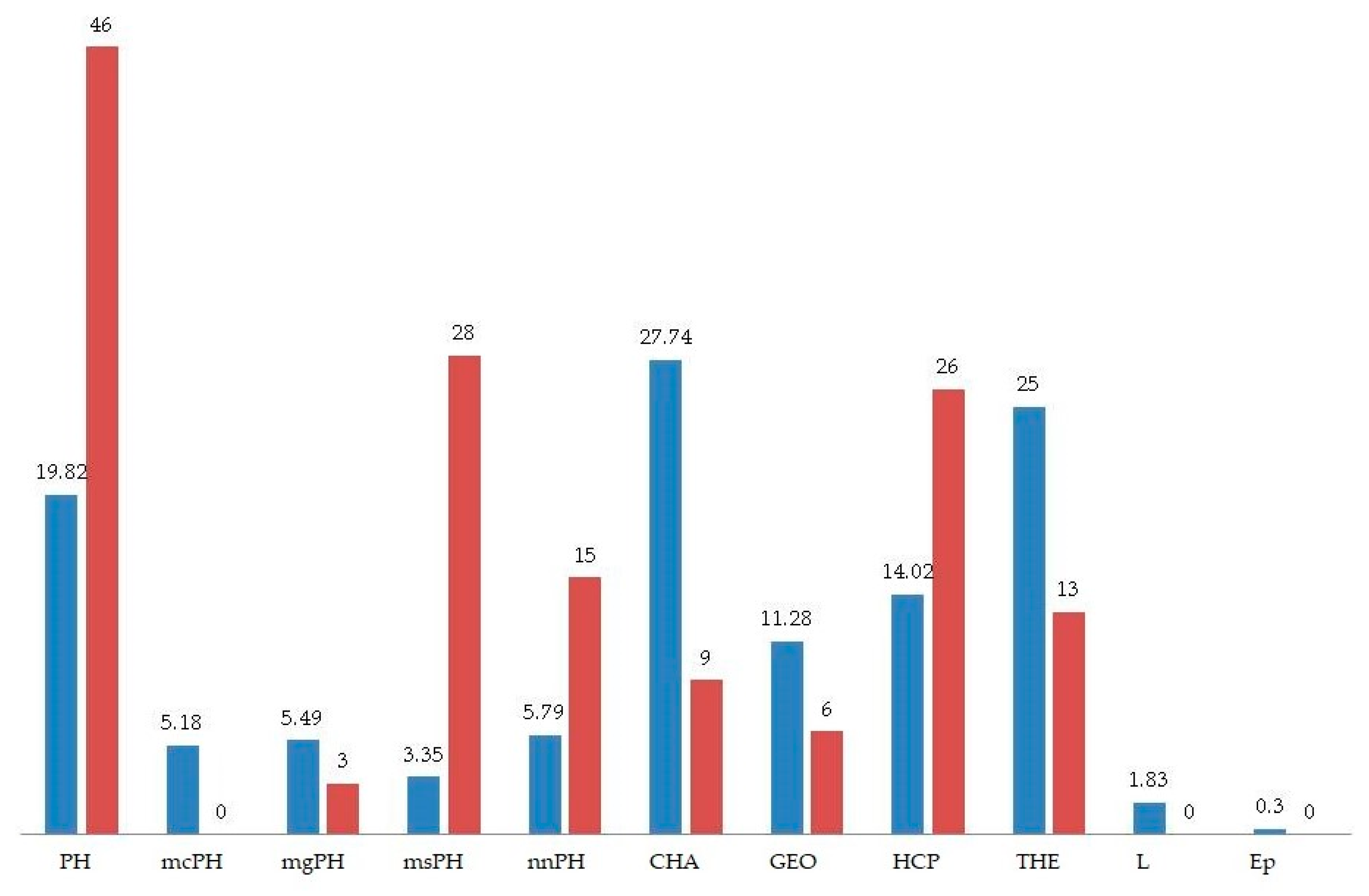
3.3. Life Forms and Biological Spectrum
3.4. Leaf Size Spectrum
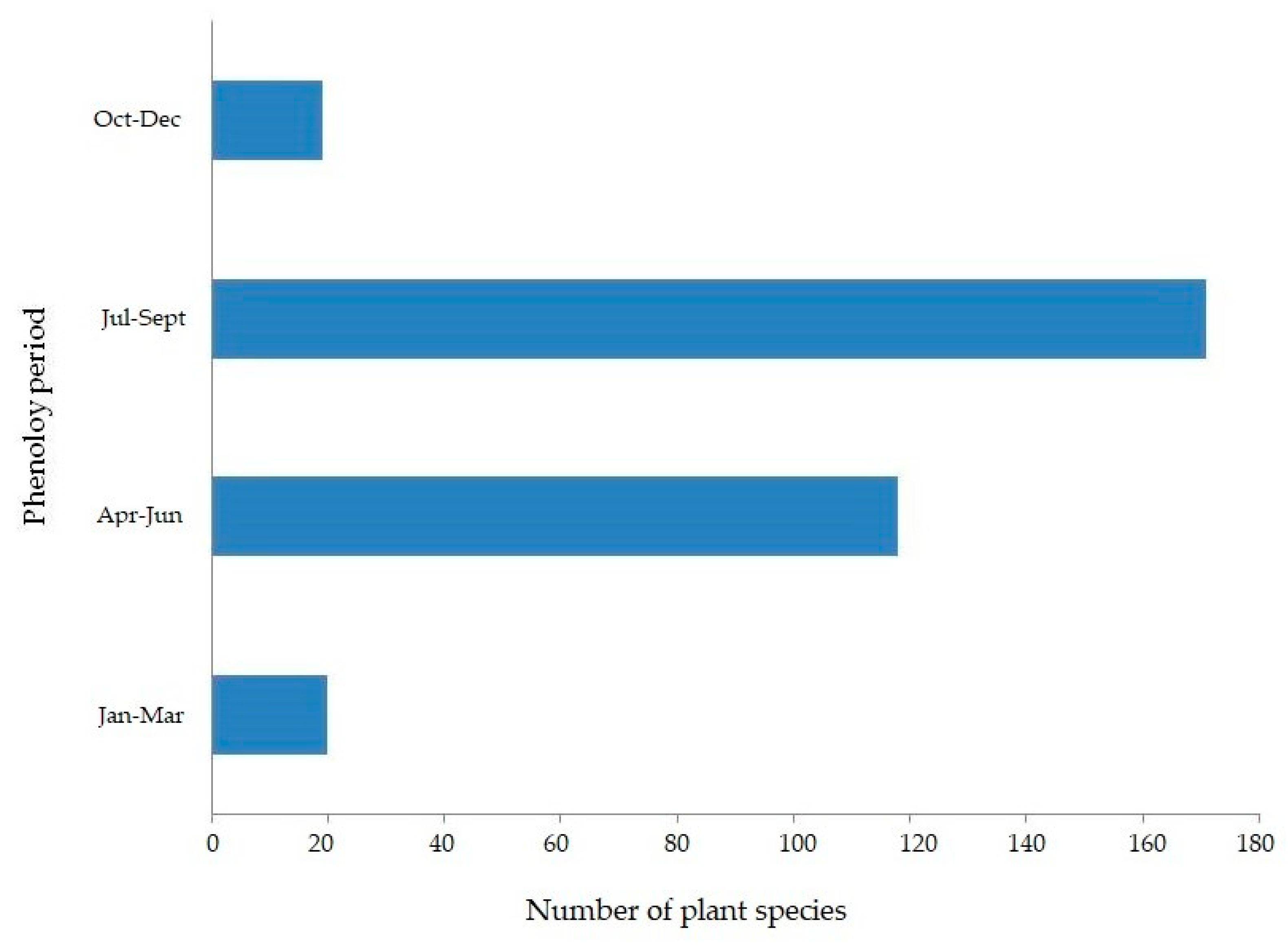
3.5. Phenological Periods
3.6. Alien Taxa
3.7. Red List IUCN and Conservation Status
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rudawska, M.; Leski, T.; Stasińska, M.; Karliński, L.; Wilgan, R.; Kujawska, M. The contribution of forest reserves and managed forests to the diversity of macrofungi of different trophic groups in European mixed coniferous forest ecosystem. For. Ecol. Manag. 2022, 518, 120274. [Google Scholar] [CrossRef]
- Hooper, D.; Adair, E.C.; Caedinale, B.J.; Byrnes, J.E.; Hungate, B.A.; Matulich, K.L.; O’Conner, M.I. A global synthesis reveals biodiversity loss as major driver of ecosystem, change. Nature 2012, 486, 1050108. [Google Scholar] [CrossRef] [PubMed]
- daSilva, L.P.; Heleno, R.H.; Costa, J.M.; Valente, M.; Mata, V.A.; Gonçalves, S.C.; Ramos, J.A. Natural woodlands hold more diverse, abundant, and unique biota than novel anthropogenic forests: A multi-group assessment. Eur. J. For. Res. 2019, 138, 461–472. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, W.; Niu, K.; Du, G. Realistic loss of plant species diversity decreases soil quality in a Tibetan alpine meadow. Agric. Ecosyst. Environ. 2019, 279, 25–32. [Google Scholar] [CrossRef]
- Gong, P.; Pu, R.; Yu, B. Conifer species recognition: An exploratory analysis of in-situ hyperspectral data. Remote Sens. Environ. 1997, 62, 189–200. [Google Scholar] [CrossRef]
- Halme, P.; Holec, J.; Heilmann-Clausen, J. The history and future of fungi as biodiversity surrogates in forests. Fungal Ecol. 2017, 27, 193–201. [Google Scholar] [CrossRef]
- Tomao, A.; Bonet, J.A.; Castano, C.; de-Miguel, S. How does forest management affect fungal diversity and community composition? Current knowledge and future perspectives for the conservation of forest fungi. Forest Ecol. Manag. 2020, 457, 117678. [Google Scholar] [CrossRef]
- Mendes, P.; Meireles, C.; Vila-Viçosa, C.; Musarella, C.; Pinto-Gomes, C. Best management practices to face degraded territories occupied by Cistus ladanifer shrublands—Portugal case study. Plant Biosyst. 2015, 149, 494–502. [Google Scholar] [CrossRef]
- Moreno, C.E.; Calderon-Patron, J.M.; Martin-Regalado, N.; Martinez-Falcon, A.P.; Ortega-Martinez, I.J.; Rios-Diaz, C.L.; Rosas, F. Measuring species diversity in the tropics: A review of methodological approaches and framework for future studies. Biotropica 2018, 50, 929–941. [Google Scholar] [CrossRef]
- Pauw, K.D.; Meeussen, C.; Govaert, S.; Sanczuk, P.; Vanneste, T.; Bernhardt-Römermann, M.; Bollmann, K.; Brunet, J.; Calders, K.; Cousins, S.A.O.; et al. Taxonomic, phylogenetic and functional diversity of understorey plants respond differently to environmental conditions in European forest edges. J. Ecol. 2021, 109, 2627–2648. [Google Scholar]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.L.; Gardiner, B.; González-Olabarria, J.R.; Jactel, H. Forest Biodiversity, ecosystem, functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef]
- Lelli, C.; Brunn, H.H.; Chiarucci, A.; Donati, D.; Frascaroli, F.; Fritz, O.; Goldberg, I.; Nascimbene, J.; Tottrup, A.P.; Rahbek, C.; et al. Biodiversity response to forest structure and management: Comparing species richness, conservation relevant species and functional diversity as metrics in forests. Forest Ecol. Manag. 2019, 432, 707–717. [Google Scholar] [CrossRef]
- Perrino, E.V.; Musarella, C.M.; Magazzini, P. Management of grazing Italian river buffalo to preserve habitats defined by Directive 92/43/EEC in a protected wetland area on the Mediterranean coast: PaludeFrattarolo, Apulia, Italy. Euro-Mediterr. J. Environ. Integr. 2021, 6, 32. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef]
- Crouzeilles, R.; Curram, M.; Ferreira, M.S.; Lindernmayer, D.B.; Grelle, C.E.V.; Rey Benayas, J.M. A global meta-analysis on the ecological drivers of forest restoration success. Nat. Commun. 2016, 7, 11666. [Google Scholar] [CrossRef]
- Leader-Williams, N.; Heywood, V.H. Global biodiversity assessment. J. Anim. Ecol. 1996, 65, 853–854. [Google Scholar] [CrossRef]
- Adhikari, D.; Tiwari, R.; Singh, P.P.; Upadhaya, K.; Singh, B.; Haridashan, K.E.; Bhat, B.B.; Chetri, A.; Barik, S.K. Ecological niche modelling as a cumulative environmental impact assessment tool for biodiversity assessment and conservation planning: A case study of critically endangered plant Lagerstroemia minuticarpa in the Eastern Himalaya. J. Environ. Manag. 2019, 243, 299–307. [Google Scholar] [CrossRef]
- Corona, P.; Blasi, C.; Chirici, G.; Facioni, L.; Fattorini, L.; Ferrari, B. Monitoring and assessing old-growth forest stands by plot sampling. Plant Biosyst. 2010, 144, 171–179. [Google Scholar] [CrossRef]
- Dansereau, P.; Guinochet, M.; Tomaselli, R. Two new European commentaries on phytosociology. Ecology 1957, 38, 364–366. [Google Scholar] [CrossRef]
- Singh, B.; Singh, B.; Singh, S.; Bhanwaria, R.; Chandra, S. Biological spectrum and floral diversity of Western Himalaya: A case study of Nandini Wildlife Sanctuary in J&K State. In Angiosperm Systematics: Recent Trends and Issues; Agnihotri, P., Kuraijam, J.S., Eds.; Bishen Singh Mahendra Pal Singh: Dehradun, India, 2018; pp. 589–605. [Google Scholar]
- Blasi, C.; Biondi, E.; Izco, J. 100 years of plant sociology: A celebration. Plant Biosyst. 2011, 145, 1–3. [Google Scholar] [CrossRef]
- Torresani, M.; Rocchini, D.; Sonnenschein, R.; Zebisch, M.; Marcantonio, M.; Ricotta, C.; Tonon, G. Estimating tree species diversity from space in an alpine conifer forest: The Rao’s Q diversity index meets the spectral variation hypothesis. Ecol. Inform. 2019, 52, 26–34. [Google Scholar] [CrossRef]
- Raunkiaer, C.; Gilbert-Carter, H.; Fausboll, A.; Tansley, A.G. The Life Forms of Plants and Statistical Plant Geography; The Clarendron Press: Oxford, MI, USA, 1934. [Google Scholar]
- daCosta, C.R.; De Araújo, S.F.; Lima-Verde, W.L. Flora and life-form spectrum in an area of deciduous thorn woodland (caatinga) in northeastern, Brazil. J. Arid Environ. 2007, 68, 237–247. [Google Scholar]
- Al-Yemeni, M.; Sher, H. Biological spectrum with some other ecological attributes of the flora and vegetation of the Asir Mountain of South West, Saudi Arabia. Afr. J. Biotechnol. 2010, 9, 5550–5559. [Google Scholar]
- Reddy, S.C.; Krishna, P.H.; Meena, S.L.; Bhardwaj, R.; Sharma, K.C. Composition of life forms and biological spectrum along climatic gradient in Rajasthan, India. Int. J. Environ. Sci. 2011, 1, 1632–1639. [Google Scholar]
- Badshah, L.; Hussain, F.; Sher, Z. Floristic inventory, ecological characteristics and biological spectrum of rangeland, district Tank, Pakistan. Pak. J. Bot. 2013, 45, 1159–1168. [Google Scholar]
- Hussain, F.; Shah, S.M.; Badshah, L.; Durrani, M.J. Diversity and ecological characteristics of flora of Mastuj valley, district Chitral, Hindukush range, Pakistan. Pak. J. Bot. 2015, 47, 495–510. [Google Scholar]
- Cano, E.; Musarella, C.M.; Cano-Ortiz, A.; Piñar Fuentes, J.C.; Spampinato, G.; Pinto Gomes, C.J. Morphometric analysis and bioclimatic distribution of Glebionis coronarias L. (Asteraceae) in the Mediterranean area. PhytoKeys 2017, 81, 103–126. [Google Scholar] [CrossRef]
- Musarella, C.M.; Cano-Ortiz, A.; Piñar Fuentes, J.C.; Navas-Ureña, J.; Pinto Gomes, C.J.; Quinto-Canas, R.; Cano, E.; Spampinato, G. Similarity analysis between species of the genus Quercus L. (Fagaceae) in southern Italy based on the fractal dimension. PhytoKeys 2018, 113, 79–95. [Google Scholar] [CrossRef]
- Zarezadeh, A.; Mirvakili, S.M.; Mirhossaini, A. Introduction to the flora, life form and plant geographical distribution of Darrah Damgahan in Mehriz (Yazd province), Pajouhesh. Sazandegi 2007, 20, 129–137. [Google Scholar]
- Wright, I.J.; Dong, N.; Maire, V.; Prentice, I.C.; Westoby, M.; Díaz, S.; Gallagher, R.V.; Jacobs, B.F.; Kooyman, R.; Law, E.A.; et al. Global climatic drivers of leaf size. Science 2017, 357, 917–921. [Google Scholar] [CrossRef]
- Greig-Smith, P.; Shimwell, D.W. The Description and classification of vegetation. J. Ecol. 1973, 61, 313. [Google Scholar] [CrossRef]
- Mehmood, A.; Shah, A.H.; Shah, A.H.; Khan, S.M.; Rahman, I.U.; Ahmad, H. Floristic List and Indigenous Uses of Poaceae Family in District Tor Ghar, Khyber Pakhtunkhwa, Pakistan. J. Appl. Environ. Biol. Sci. 2017, 7, 169–177. [Google Scholar]
- Lechowicz, M.J. The Earth System: Biological and Ecological Dimensions of Global Environmental Change; Wiley: London, UK, 2001. [Google Scholar]
- Saini, A.; Singh, B. Tribal knowledge on ethnobotanical plants of Uttarakhand Himalaya: Review research documentary. In Plants for Novel Drug Molecules-Ethnobotany to Ethnopharmacology; Singh, B., Sharma, Y.P., Eds.; New India Publishing Agency: New Delhi, India, 2021; pp. 91–145. [Google Scholar]
- Gillison, A.N. Latitudinal Variation in Plant Functional Types; Springer International Publishing AG: Cham, Switzerland, 2018. [Google Scholar]
- Cain, S.A.; Castro, G.D.O. Manual of Vegetation Analysis; Harper and Brothers: New York, NY, USA, 1959. [Google Scholar]
- Meher-Homji, V.M. Life forms and biological spectra as epharmonic criteria of aridity and humidity in tropics. J. Indian Bot. Soc. 1964, 43, 424–430. [Google Scholar]
- Rana, T.S.; Datt, B.; Rao, R.R. Life forms and biological spectrum of the flora of Tons valley, Garhwal Himalaya (Uttaranchal) India. Taiwania 2002, 47, 164–169. [Google Scholar]
- Austin, M.P. The potential contribution of vegetation ecology to biodiversity research. Ecography 1999, 22, 465–484. [Google Scholar] [CrossRef]
- Abd El-Ghani, M.M.; Amer, W.M. Soil-vegetation relationships in a coastal desert plain of southern Sinai, Egypt. J. Arid Environ. 2003, 55, 607–628. [Google Scholar] [CrossRef]
- Körner, C. Mountain biodiversity, its causes and function. Ambio 2004, 33, 11–17. [Google Scholar] [CrossRef]
- Ortiz, A.C.; Canas, R.Q.; Fuentes, J.C.P.; del Rio, S.; Gomes, C.J.P.; Cano, E. Endemic Hemicryptophyte Grasslands of the High Mountains of the Caribbean. Res. J. Ecol. Environ. Sci. 2022, 2, 1–20. [Google Scholar] [CrossRef]
- Hajra, P.K.; Rao, R.R. Distribution of vegetation types in Northwest Himalayas with brief remarks on phytogeography and floral resource conservation. Proc. Natl. Acad. Sci. India Sect. B-Biol. Sci. 1990, 100, 236–277. [Google Scholar] [CrossRef]
- Musarella, C.M.; Brullo, S.; Del Galdo, G.G. Contribution to the orophilous cushion-like vegetation of central-southern and insular Greece. Plants 2020, 9, 1678. [Google Scholar] [CrossRef]
- Ighbareyeh, J.M.H.; Suliemieh, A.A.-R.A.; Sheqwarah, M.; Cano-Ortiz, A.; Carmona, E.C. Flora and Phytosociological of Plant in Al-Dawaimah of Palestine. Res. J. Ecol. Environ. Sci. 2022, 2, 58–91. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389. [Google Scholar] [CrossRef] [PubMed]
- Thuiller, W.; Richardson, D.M.; Midgley, G.F. Will Climate Change Promote Alien Plant Invasions? Springer: Berlin/Heidelberg, Germany, 2008; pp. 197–211. [Google Scholar]
- Del Río, S.; Canas, R.; Cano, E.; Cano-Ortiz, A.; Musarella, C.; Pinto-Gomes, C.; Penas, A. Modelling the impacts of climate change on habitat suitability and vulnerability in deciduous forests in Spain. Ecol. Indic. 2021, 131, 108202. [Google Scholar] [CrossRef]
- Stinca, A.; Musarella, C.M.; Rosati, L.; Laface, V.L.A.; Licht, W.; Fanfarillo, E.; Wagensommer, R.P.; Galasso, G.; Fascetti, S.; Esposito, A.; et al. Italian vascular flora: New findings, updates and exploration of floristic similarities between regions. Diversity 2021, 13, 600. [Google Scholar] [CrossRef]
- Cano, E.; Cano-Ortiz, A.; Fuentes, J.C.P.; Quinto-Canas, R.; Igbareyeh, J.; del Río, S.; Gomes, C.J.P. Ecological and Syntaxonomic Analysis of Pinus halepensis Mill. in the Iberian Peninsula and Balearic Islands. Land 2022, 11, 369. [Google Scholar] [CrossRef]
- Cano, E.; Veloz Ramirez, A.; Cano Ortiz, A. Phytosociological study of the Pinus occidentalis forests in the Dominican Republic. Plant Biosyst. 2011, 145, 286–297. [Google Scholar] [CrossRef]
- Cano, E.; Rodríguez Torres, A.; Pinto Gomes, C.J.; García Fuentes, A.; Torres, J.A.; Salazar, C.; Ruiz, L.; Cano Ortiz, A.; Montilla, R. Analysis of the Juniperus oxycedrus L. in the centre and south of the Iberian peninsula (Spain and Portugal). Acta Bot. Gallica 2007, 154, 79–99. [Google Scholar] [CrossRef]
- Ortiz, A.C.; Spampinato, G.; Fuentes, J.C.P.; Gomes, C.J.P.; Quinto-Canas, R.; Cano, E. Taxonomy, Ecology and Distribution of Juniperus oxycedrus L. Group in the Mediterranean Basin Using Bioclimatic, Phytochemical and Morphometric Approaches, with Special Reference to the Iberian Peninsula. Forests 2021, 12, 703. [Google Scholar] [CrossRef]
- Cano, E.; Musarella, C.M.; Ortiz, A.C.; Fuentes, J.C.P.; Torres, A.R.; Del Río González, S.; Gomes, C.J.P.; Quinto-Canas, R.; Spampinato, G. Geobotanical Study of the Microforests of Juniperus oxycedrus subsp. badia in the Central and Southern. Sustainability 2019, 11, 1111. [Google Scholar] [CrossRef]
- Cano Ortiz, A.; Piñar, J.C.; Pinto Gomes, C.J.; Musarella, C.M.; Cano, E. Expansion of the Juniperus genus due to anthropic activity. In Old-Growth Forest and Coniferous Forests; Weber, R.P., Ed.; Nova Science Publishers: New York, NY, USA, 2015; pp. 55–65. [Google Scholar]
- Sahib, N.; Boumediene, M.; Abid, M.; Mihamou, A.; Serghini-Caid, H.; Elamrani, A.; Hano, C.; Addi, M. Phenotypic Comparison of Three Populations of Juniperus turbinata Guss. in North-Eastern Morocco. Forests 2022, 13, 287. [Google Scholar] [CrossRef]
- García Morote, F.A.; Andrés Abellán, M.; Rubio, E.; Pérez Anta, I.; García Saucedo, F.; López Serrano, F.R. Stem CO2 Efflux as an Indicator of Forests’ Productivity in Relict Juniper Woodlands (Juniperus thurifera L.) of Southern Spain. Forests 2021, 12, 1340. [Google Scholar] [CrossRef]
- Hooker, J.D. The Flora of British India; L. Leeve& Co.: London, UK; Volume 7, pp. 1872–1879.
- Bargalii, H.; Kumar, A.; Singh, P. Plant studies in Uttarakhand, Western Himalaya: A comprehensive review. Trees Forest. People 2022, 8, 100203. [Google Scholar] [CrossRef]
- Singh, A.K.; Goswami, M. Life in the Himalaya: An ecosystem at risk. Delhi Bus. Rev. 2018, 19, 121–122. [Google Scholar] [CrossRef]
- Singh, B.; Singh, B.; Kishor, A.; Singh, S.; Bhat, M.N.; Surmal, O.; Musarella, C.M. Exploring plant-based ethnomedicine and quantitative ethnopharmacology: Medicinal plants utilized by the population of Jasrota Hill in Western Himalaya. Sustainability 2020, 12, 7526. [Google Scholar] [CrossRef]
- Don, D. Prodomus Florae Nepalensis; Govt. Press: Dehradun, India, 1825. [Google Scholar]
- Atkinson, E.T. The Himalayan Gazatteer; Cosmo Publication: Delhi, India, 1882. [Google Scholar]
- Duthie, J.F. Catalogue of the Plants of Kumaun and of the Adjacent Portions of Garhwal and Tibet on the Collections Made by Strachey and Winterbottom during the Years 1864–1849, on the Catalogue Originally Prepared in 1852 by Sir Richard Stachey; Lovell Reeve & Co. Limited: London, UK, 1906. [Google Scholar]
- Strachey, R.; Winterbottom, J.E. Scientific botany. In The Himalayan Districts of the NW provinces of India; Atkinson, E.T. Cosmo Publication: New Delhi, India, 1882. [Google Scholar]
- Kanoyer, I.A. Forest formation and succession in Satlej Valley, Kumaun Himalaya. J. Indian Bot. Soc. 1921, 2, 8–9. [Google Scholar]
- Osmaston, A.E. Notes on forest communities of Gharwal Himalaya. J. Ecol. 1922, 10, 129–167. [Google Scholar] [CrossRef]
- Kashyap, S.R. Vegetation of Western Himalaya and Western Tibet in relation to their climates. J. Indian Bot. Soc. 1932, 4, 327–334. [Google Scholar]
- Giorrie, R.M. The Sutlej Deodar, its ecology and timber production. Indian forest production. Indian For. Rec. 1933, 17, 1–240. [Google Scholar]
- Puri, G.S.; Gupta, A.C. The Himalayan conifers II: The ecology of Humans in conifer forests of Kullu Himalaya. Indian For. 1951, 77, 124–129. [Google Scholar]
- Champion, H.G.; Seth, S.K. A Revised Survey of Forest Types of India; Govt. of India: Calcutta, India, 1968.
- Mani, M.S. Biogeography evolution in India. In Ecology and Biogeography in India; Mani, M.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1974; pp. 698–724. [Google Scholar]
- Clarke, R. Ecological of Nanda Devi area. Am. Alp. J. 1979, 53, 281. [Google Scholar]
- Polunin, O.; Stainton, A. Flowers of Himalaya; Oxford University Press: Delhi, India; Bombay, India; Calcutta, India; Madras, India, 1984. [Google Scholar]
- Adhikari, B.S.; Rikhari, H.C.; Rawat, Y.S.; Singh, S.P. High altitude forest: Composition diversity and profile structure in a part of KumaunHimalya. Trop. Ecol. 1991, 32, 86–97. [Google Scholar]
- Bankoti, N.S.; Rawal, R.S.; Samant, S.S.; Pangtey, Y.P.S. Forest vegetation of inner hill ranges in Komaun, Central Himalaya. Trop. Ecol. 1992, 33, 41–53. [Google Scholar]
- Ram, J. Effect of clipping on aboveground biomass and total herbage yield in grassland above tree line in central Himalaya. India. Arc. Alp. Res. 1922, 24, 456–468. [Google Scholar]
- Dar, G.H.; Khuroo, A.A. Floristic diversity in the Kashmir Himalaya: Progress, problems and prospects. Sains Malays. 2013, 42, 1377–1386. [Google Scholar]
- Chawla, A.; Rajkumar, S.; Singh, K.N.; Lal, B.; Singh, R.D.; Thukral, A.K. Plant Species Diversity along an Altitudinal Gradient of Bhabha Valley in Western Himalaya. J. Mt. Sci. 2008, 5, 157–177. [Google Scholar] [CrossRef]
- Samant, S.S.; Dhar, U.; Palni, L.M.S. Medicinal Plants of Indian Himalaya: Diversity, Distribution and Potential Values; GyanodayaPrakashan: Nainital, India, 1998. [Google Scholar]
- Rawat, G.S. Alpine Meadows of Uttaranchal; Bishen Singh Mahenda Pal Singh: Dehradun, India, 2005. [Google Scholar]
- Kumar, A.; Mitra, M.; Adhikari, B.S.; Rawat, G.S. Flora of Niti valley: A cold arid region of Nanda Devi Biosphere Reserve, Western Himalaya, India. Check List 2016, 12, 1824. [Google Scholar] [CrossRef]
- Singh, B.; Singh, S.; Kishor, A.; Singh, B. Traditional usage of medicinal plants in humans and animal health care and their chemical constituents from hills and valleys of Jammu province, Western Himalaya. Indian J. Nat. Prod. Resour. 2021, 22, 84–100. [Google Scholar]
- Singh, S.; Singh, B.; Surmal, O.; Bhat, M.N.; Singh, B.; Musarella, C.M. Fragmented forest patches in Indian Himalaya preserves unique components of biodiversity: Investigation on floristic and phytoclimate of unexplored Bani valley. Sustainability 2021, 13, 6063. [Google Scholar] [CrossRef]
- Singh, S.; Sneha; Singh, B. Pyrrosiasarthalensis, a new species of Polypodiaceae from Kathua District, Jammu and Kashmir, Western Himalaya, India. Phytotaxa 2022, 522, 249–255. [Google Scholar] [CrossRef]
- Bhat, M.N.; Singh, B.; Surmal, O.; Singh, B.; Shivgotra, V.; Musarella, C.M. Ethnobotany of the Himalayas: Safeguarding medical practices and traditional uses of Kashmir regions. Biology 2021, 10, 851. [Google Scholar] [CrossRef]
- Singh, B. Therapeutic Himalayan herbs: Folklore uses, bioactive phytochemicals, and biological activities medicinal orchids used by Nomads. Indian J. Nat. Prod. Resour. 2022, 23, 94–104. [Google Scholar]
- Haq, S.M.; Hamid, M.; Lone, F.A.; Singh, B. Himalayan Hotspot with Alien Weeds: A Case Study of Biological Spectrum, Phenology, and Diversity of Weedy Plants of High-Altitude Mountains in District Kupwara of J&K Himalaya. Proc. Natl. Acad. Sci. India Sect. B-Biol. Sci. 2021, 91, 139–152. [Google Scholar]
- Mir, A.H.; Javaid, M.D.; Singh, B.; Kamili, A.N. Passive restoration considerably improved the community structure, soil health and carbon stock in the Pine forests of Kashmir Himalaya, India. Ecol. Eng. 2022, 176, 106535. [Google Scholar] [CrossRef]
- Rudolph, S.; Maciá-Vicente, J.G.; Lotz-Winter, H.; Schleuning, M.; Piepenbring, M. Temporal variation of fungal diversity in a mosaic landscape in Germany. Stud. Mycology 2018, 89, 95–104. [Google Scholar]
- Jain, S.K.; Rao, R.R. A Handbook of Field and Herbarium Methods; Today and Tomorrow’s Printers: New Delhi, India, 1977. [Google Scholar]
- Theirs, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff; New York Botanical Garden’s Virtual Herbarium: New York, NY, USA, 2019. [Google Scholar]
- Swami, A.; Gupta, R.K. Flora of Udhampur; Bishen Singh Mahendra Pal Singh: Dehradun, India, 1998. [Google Scholar]
- Sharma, B.M.; Kachroo, P. Flora of Jammu and Plants of Neighbourhood; Bishen Singh Mahendra Pal Singh: Dehradun, India, 1981. [Google Scholar]
- Kapur, S.K.; Sarin, Y.K. Flora of Trikuta Hills (Shri Vaishno Devi Shrine) with Special Reference to the Distribution Pattern of Minor Forest Products; Bishen Singh Mahendra Pal Singh: Dehradun, India, 1990. [Google Scholar]
- Sharma, B.M. Illustrations of Jammu Plants: A Supplement of Flora of Jammu and Plants of Neighbourhood with General Key and Annotation; Bishen Singh Mahendra Pal Singh: Dehradun, India, 2010. [Google Scholar]
- Perez-Harguindeguy, N.; Daiz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Ali, A.; Badshah, L.; Hussain, F.; Shinwari, Z.K. Floristic composition and ecological characteristics of plants of Chail valley, district Swat, Pakistan. Pak. J. Bot. 2013, 48, 1013–1026. [Google Scholar]
- Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; Stevens, P.F.; et al. An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar]
- Misra, R. Ecology Workbook; Oxford and IBH Publication Co.: Calcutta, India, 1968. [Google Scholar]
- Weaver, W.; Shannon, C.E. The Mathematical Theory of Communication; University of lllinois Press: Urbana, IL, USA, 1963. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Science: Oxford, UK, 2004. [Google Scholar]
- Pieluo, E.C. Ecological Diversity; Wiley & Sons Publications: New York, NY, USA, 1975. [Google Scholar]
- Ford, R. Flowers of the Himalaya, by Oleg Polunin & Adam Stainton; Oxford University Press: Delhi, India; Bombay, India; Calcutta, India; Madras, India; Oxford, UK, 1984. [Google Scholar]
- Sharma, P.; Rana, J.C.; Devi, U.; Randhawa, S.S.; Kumar, R. Floristic diversity and distribution pattern of plant communities along altitudinal gradient in Sangla valley, northwest Himalaya. Sci. World J. 2014, 2014, 264878. [Google Scholar] [CrossRef]
- Agrawal, A.K. Floristic composition and phenology of temperate grasslands of Western Himalaya as affected by scraping, fire and heavy grazing. Vegetatio 1990, 88, 177–187. [Google Scholar] [CrossRef]
- Shaheen, H.; Ullah, Z.; Khan, S.M.; Harper, D.M. Species composition and community structure of western Himalayan moist temperate forests in Kashmir. For. Ecol. Manag. 2012, 278, 138–145. [Google Scholar] [CrossRef]
- Haq, S.M.; Malik, A.H.; Khuroo, A.A.; Rashid, I. Floristic composition and biological spectrum of Keran—A remote valley of northwestern Himalaya. Acta Ecol. Sin. 2019, 39, 372–379. [Google Scholar] [CrossRef]
- Wen, J.; Qin, R.; Zhang, S.; Yang, X.; Xu, M. Effects of long-term warming on the aboveground biomass and species diversity in an alpine meadow on the Qinghai-Tibetan Plateau of China. J. Arid. Land. 2020, 12, 252–266. [Google Scholar] [CrossRef]
- Malik, Z.A.; Bhatt, A.B. Regeneration status of tree species and survival of their seedlings in Kedarnath Wildlife Sanctuary and its adjoining area in Western Himalaya, India. Trop. Ecol. 2016, 57, 677–690. [Google Scholar]
- Tiwari, O.P.; Rana, Y.S.; Krishan, R.; Sharma, C.M.; Bhandari, B.S. Regeneration dynamics, population structure and forest composition in some ridge forests of the Western Himalaya, India. For. Sci. Technol. 2018, 14, 66–75. [Google Scholar] [CrossRef]
- Haq, S.M.; Calixto, E.S.; Kumar, M. Assessing biodiversity and productivity over a small-scale gradient in the protected forests of Indian Western Himalayas. J. Sustain. For. 2021, 40, 675–694. [Google Scholar] [CrossRef]
- Subramani, S.; Vaneet, J.; Verma, R. Floristic composition, life-forms and biological spectrum of Renuka Wildlife Sanctuary, Himachal Pradesh. Indian For. 2007, 133, 79–92. [Google Scholar]
- Saha, S. Determination of lifeform patterns in the Singalila range of Darjeeling. Environ. Ecol. 2008, 26, 588–590. [Google Scholar]
- Rawat, G.S.; Adhikari, B.S. Floristics and distribution of plant communities across moisture and topographic gradients in Tso Kar basin, Changtnang plateau, eastern Ladakh. Arctic, Antarct. Alp. Res. 2008, 37, 539–544. [Google Scholar]
- Namgail, T.; Rawat, G.S.; Mishra, C.; Wieren, S.E.V.; Prins, H.H.T. Biomass and diversity of dry alpine plant communities along altitudinal gradients in the Himalayas. J. Plant Res. 2012, 125, 93–101. [Google Scholar] [CrossRef]
- Pharswan, K.; Mehta, J.P. Floristic composition and biological spectrum of vegetation in alpine meadows of Kedarnath: Garhwal Himalaya. October 2010, 8, 109–115. [Google Scholar]
- Nautiyal, M.C.; Nautiyal, B.P.; Prakash, V. Phenology and growth form distribution in an alpine pasture at Tungnath. Mt. Res. Dev. 2001, 21, 168–174. [Google Scholar] [CrossRef]
- Mir, N.A.; Masood, T.H.; Sofi, P.A.; Husain, M.; Rather, T.A. Life form spectrum of vegetation in Betula dominant tree stands along the available altitudinal gradient in north western Himalayas of Kashmir. J. Pharmacogn. Phytochem. 2017, 6, 267–272. [Google Scholar]
- Dogra, K.S.; Sood, S.K.; Sharma, R. Distribution, Biology and Ecology of Parthenium hysterophorus L. (Congress Grass) an invasive species in the North-Western Indian Himalaya (Himachal Pradesh). Afr. J. Plant Sci. 2011, 5, 682–687. [Google Scholar]
- Gaston, A.J.; Garson, P.J.; Hunter, M.L. The status and conservation of forest wildlife in Himachal Pradesh, Western Himalayas. Biol. Conserv. 1983, 27, 291–314. [Google Scholar] [CrossRef]
- Gairola, S.; Sharma, C.M.; Suyal, S.; Ghildiyal, S.K. Composition and diversity of five major forest types in moist temperate climate of the western Himalayas. For. Stud. China 2011, 13, 139–153. [Google Scholar] [CrossRef]
- Ricklefs, R.D. Ecology; Thomas Nelson and Sons Ltd.: Edinburgh, UK, 1979. [Google Scholar]
- Mohr, C.O.; Smith, R.L. Ecology and field biology. J. Wildl. Manag. 1967, 31, 393–395. [Google Scholar] [CrossRef]
- Deljouei, A.; Abdi, E.; Marcantonio, M.; Majnounian, B.; Amici, V.; Sohrabi, H. The impact of forest roads on understory plant diversity in temperate hornbeam-beech forests of Northern Iran. Environ. Monit. Assess. 2017, 189, 392. [Google Scholar] [CrossRef]
- Cain, S.A. Life-forms and phytoclimate. Bot. Rev. 1950, 16, 1–32. [Google Scholar] [CrossRef]
- Daubenmire, R. Plant Communities; A text book of plant synecology; Harper and Row, Publ.: New York, NY, USA, 1950. [Google Scholar]
- Vashistha, R.K.; Rawat, N.; Chaturvedi, A.K.; Nautiyal, B.P.; Prasad, P.; Nautiyal, M.C. Characteristics of life-form and growth-form of plant species in an alpine ecosystem of North-West Himalaya. J. For. Res. 2011, 22, 501–506. [Google Scholar] [CrossRef]
- Saxena, A.K.; Pandey, P.; Singh, J.S. Biological spectrum and other structural-functional attributes of the vegetation of Kumaun Himalaya. Vegetatio 1982, 49, 111–119. [Google Scholar] [CrossRef]
- Singh, B.; Bedi, Y.S. Eating from raw wild plants in Himalaya: Traditional knowledge documentary on Sheena tribe in Kashmir. Indian J. Nat. Prod. Resour. 2017, 8, 269–275. [Google Scholar]
- Das, D.S.; Rawat, D.S.; Maity, D.; Dash, S.S.; Sinha, B.K. Species Richness Patterns of Different Life-Forms along Altitudinal Gradients in the Great Himalayan National Park, Western Himalaya, India. Taiwania 2020, 65, 154–162. [Google Scholar]
- Dobhal, P.; Bisht, S.L.; Smriti, S.; Venita, J.; Joshi, S.P. Life-forms and biological spectrum of a riverine forest of the Doon valley, Uttarakhand, India. Indian J. For. 2010, 33, 585–598. [Google Scholar] [CrossRef]
- Ghildiyal, J.C.; Meenakshi, J. Biodiversity of Lal Dhang Forest Range of Lansdowne Forest Division in Garhwal Himalaya. Indian For. 2010, 136, 742–766. [Google Scholar]
- Ghildiyal, J.C.; Barthwal, N.; Khantwal, A. Life forms and biological spectrum of Deeva ka Daanda Sacred groove in Gharwal Himalaya. Indian For. 2016, 142, 167–178. [Google Scholar]
- Thakur, M.; Santvan, V.K.; Nigam, A. Floristic composition and biological spectrum of Darlaghat wildlife sanctuary Solan Himachal Pradesh, India. N. Y. Sci. J. 2012, 5, 1–14. [Google Scholar]
- Kapoor, K.S.; Singh, R. Life forms and biological spectrum of the temperate grasslands with varying biotic pressure in Shimla Hills, Himachal Pradesh. Range Manag. Agrofor. 1990, 11, 109–114. [Google Scholar]
- Sharma, J.; Raina, A.K. Lifeform classification and biological spectrum of Nandini wildlife sanctuary, Jammu, J&K, India. Environ. Conserv. J. 2017, 18, 231–237. [Google Scholar]
- Raina, A.K.; Kumar, R. Floristic composition, life form classification and biological spectrum of the catchment of Rattle HE project, District Kishtwar-J and K. Environ. Conserv. J. 2011, 12, 49–57. [Google Scholar]
- Chowdhery, H.J.; Wadhwa, B.M. Flora of Himachal Pradesh; Botanical Survey of India: Calcutta, India, 1984. [Google Scholar]
- Prance, G.T. Discovering the plant world. Taxon 2001, 50, 345–359. [Google Scholar] [CrossRef]
- Kala, C.P. Phenology of alpine plants in the valley of flowers national park and Hemkund, Western Himalaya. Indian For. 1999, 125, 581–590. [Google Scholar]
- Pangtey, Y.P.S.; Rawal, R.S.; Bankoti, N.S.; Samant, S.S. Phenology of high-altitude plants of Kumaun in Central Himalaya, India. Int. J. Biometeorol. 1990, 34, 122–127. [Google Scholar] [CrossRef]
- Singh, B.; Borthakur, S.K. Phenology and geographic extension of lycophyta and fern flora in Nokrek Biosphere Reserve of Eastern Himalaya. Proc. Natl. Acad. Sci. India Sect. B-Biol. Sci. 2015, 85, 291–301. [Google Scholar] [CrossRef]
- Maheshwari, J.K. Studies on the Naturalised Flora of India. Proc. Summer Sch. Bot. Darjeeling 2015, 1, 156–170. [Google Scholar]
- Sharma, B.D.; Pandey, D.S. Exotic Flora of Allahabad District; Botanical Survey of India: Kolkata, India, 1984. [Google Scholar]
- Pandey, R.; Parmar, P. The exotic flora of Rajasthan. J. Econ. Taxon. Bot. 1994, 18, 105–136. [Google Scholar]
- Nagar, P.S.; Pathak, S.J.; Pandya, S.M. The alien flora of the Barda hills and its surroundings in Gujarat, India. Indian For. 2004, 27, 25–38. [Google Scholar]
- Shrestha, B.B. Invasive alien plant species in Nepal. In Frontiers of Botany; Jha, P.K., Siwakoti, M., Rajbhandary, S., Eds.; Tribhuvan University: Kirtipur, Nepal; Kathmandu, Nepal, 2016; pp. 269–284. [Google Scholar]
- Walther, G.R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pysek, P.; Kuhn, I.; Zobel, M. Alien species in a warmer world. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef]
- Musarella, C.M. Solanum torvum Sw. (Solanaceae): A new alien species for Europe. Genet. Resour. Crop Evol. 2020, 67, 515–522. [Google Scholar] [CrossRef]
- Laface, V.L.A.; Musarella, C.M.; Ortiz, A.C.; Canas, R.Q.; Cannavò, S.; Spampinato, G. Three new alien taxa for Europe and a chorological update on the alien vascular flora of Calabria (Southern Italy). Plants 2020, 9, 1181. [Google Scholar] [CrossRef]
- Musarella, C.M.; Stinca, A.; Cano-Ortíz, A.; Laface, V.L.A.; Petrilli, R.; Esposito, A.; Spampinato, G. New data on the alien vascular flora of Calabria (Southern Italy). Ann. Bot. 2020, 10, 55–66. [Google Scholar] [CrossRef]
- Reddy, C.S. Catalogue of invasive alien flora of India. Life Sci. J. 2008, 5, 84–89. [Google Scholar]
- Khuroo, A.A.; Weber, E.; Malik, A.H.; Dar, G.H.; Reshi, Z.A. Taxonomic and biogeographic patterns in the native and alien woody flora of Kashmir Himalaya, India. Nord. J. Bot. 2010, 28, 685–696. [Google Scholar] [CrossRef]
- Randall, R.P. The Introduced Flora of Australia and Its Weed Status; CRC Press: Adelaide, Australia, 2007. [Google Scholar]
- Diez, J.M.; Antonio, C.M.D.; Dukes, J.S.; Grosholz, E.D.; Olden, J.D.; Sorte, C.J.B.; Blumenthal, D.M.; Bradley, B.A.; Early, R.; Ibáñez, I.; et al. Will extreme climatic events facilitate biological invasions? Front. Ecol. Environ. 2012, 10, 249–257. [Google Scholar] [CrossRef]
- Kohli, R.K.; Dogra, K.S.; Batish, D.R.; Singh, H.P. Impact of invasive plants on the structure and composition of natural negetation of northwestern Indian Himalayas. Weed Technol. 2004, 18, 1296–1300. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Categories (APG IV) | Selected Plots in Bhallesa Study Area | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gandoh | Kahal | Chaunwari | Chilly | ||||||||||
| GPS Coordinates | 33°01′56″N, 75°54′47″E | 33°01′30″N,76° 01′50″E | 32°04′ 52″N, 75 47′42″E | 33°02′60″N, 76°02′42″E | |||||||||
| Elevation (m.a.s.l.) | 1625 to 2989 | 2132 to 3799 | 1760 to 2887 | 2225 to 4000 | |||||||||
| Sub-Categories | Fam. | Gen. | Sp. | Fam. | Gen. | Sp. | Fam. | Gen. | Sp. | Fam. | Gen. | Sp. | |
| A. Angiosperms * | I. Magnoliids | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| II. Monocots | 7 | 16 | 18 | 7 | 18 | 21 | 6 | 19 | 24 | 10 | 23 | 27 | |
| II. Eudicots | 4 | 13 | 14 | 3 | 12 | 15 | 3 | 12 | 15 | 3 | 13 | 18 | |
| III. Superrosids | 2 | 2 | 2 | 3 | 6 | 9 | 3 | 3 | 3 | 3 | 6 | 10 | |
| IV. Rosids | 21 | 48 | 57 | 15 | 33 | 36 | 19 | 43 | 52 | 16 | 33 | 39 | |
| V. Superasterids | 4 | 13 | 16 | 3 | 8 | 13 | 4 | 10 | 16 | 3 | 13 | 14 | |
| VI. Asterids | 20 | 46 | 56 | 19 | 50 | 66 | 20 | 48 | 63 | 17 | 59 | 76 | |
| Sub-Total | 59 | 137 | 164 | 50 | 127 | 160 | 55 | 135 | 173 | 52 | 147 | 184 | |
| B. Gymnosperms | 2 | 4 | 5 | 2 | 4 | 4 | 2 | 4 | 4 | 3 | 6 | 7 | |
| C. Lycophytes and Ferns | 4 | 5 | 6 | 5 | 7 | 8 | 4 | 5 | 7 | 4 | 4 | 4 | |
| ** Total | 65 | 146 | 175 | 57 | 138 | 172 | 61 | 144 | 184 | 59 | 157 | 195 | |
| Categories | Tree Community | Shrub Community | Herb Community | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cl | Ka | Ch | Gd | Cl | Ka | Ch | Gd | Cl | Ka | Ch | Gd | |
| Plant species richness | 13 | 13 | 15 | 23 | 25 | 19 | 20 | 22 | 157 | 139 | 150 | 130 |
| Number of families | 9 | 9 | 10 | 15 | 18 | 16 | 16 | 17 | 45 | 43 | 43 | 41 |
| Basal area (m2 ha−1) | 144.52 ± 29.60 | 85.08 ± 20.43 | 119.24 ± 15.18 | 44.16 ± 14.29 | - | - | - | - | - | - | - | - |
| Density (ha−1) | 240 | 136 | 216 | 192 | 1056 | 816 | 672 | 656 | 191,200 | 196,800 | 206,000 | 146,400 |
| H’ | 1.95 ± 0.50 | 2.12 ± 0.47 | 2.52 ± 0.32 | 2.78 ± 0.41 | 2.81 ± 0.36 | 2.73 ± 0.87 | 2.71 ± 0.67 | 2.92 ± 0.77 | 4.77 | 4.68 | 4.67 | 4.59 |
| D | 0.77 ± 0.27 | 0.82 ± 0.13 | 0.89 ± 0.32 | 0.91 ± 0.28 | 0.91 ± 0.37 | 0.92 ± 0.45 | 0.91 ± 0.38 | 0.94 ± 0.33 | 0.99 | 0.98 | 0.98 | 0.98 |
| J | 0.54 ± 0.08 | 0.64 ± 0.04 | 0.83 ± 0.04 | 0.71 ± 0.11 | 0.66 ± 0.06 | 0.81 ± 0.13 | 0.79 ± 0.09 | 0.84 ± 0.12 | 0.75 | 0.78 | 0.72 | 0.76 |
| Taxa | Basal Area (m2 ha−1) | Density (ha−1) | IVI | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cl | Ka | Ch | Gd | Cl | Ka | Ch | Gd | Cl | Ka | Ch | Gd | |
| Abies pindrow (Royle ex D.Don) Royle | 7.68 | 0.00 | 0.00 | 0.00 | 24 | 0 | 0 | 0 | 25.84 | 0.00 | 0.00 | 0.00 |
| Acer caesium Wall. ex Br. | 0.00 | 1.84 | 4.60 | 1.84 | 0 | 8 | 20 | 8 | 0.00 | 16.76 | 22.46 | 11.07 |
| Aesculus indica (Wall. ex Camb.) Hook.f. | 3.84 | 1.92 | 5.76 | 1.68 | 8 | 4 | 12 | 4 | 11.25 | 9.55 | 17.28 | 6.15 |
| Alnus nepalensis D.Don | 5.88 | 1.96 | 0.00 | 0.00 | 12 | 4 | 0 | 0 | 14.33 | 9.59 | 0.00 | 0.00 |
| Alnus nitida (Spach) Endl. | 0.00 | 0.00 | 3.92 | 1.28 | 0 | 0 | 8 | 4 | 0.00 | 0.00 | 10.44 | 5.82 |
| Betula utilis D.Don | 2.16 | 2.16 | 0.00 | 0.00 | 4 | 4 | 0 | 0 | 5.79 | 9.83 | 0.00 | 0.00 |
| Cedrusdeodara (Roxb. ex D.Don) G.Don | 39.16 | 24.92 | 17.80 | 42.00 | 44 | 28 | 20 | 28 | 69.11 | 62.92 | 34.53 | 56.77 |
| Celtis australis L. | 0.00 | 3.00 | 6.00 | 3.44 | 0 | 4 | 8 | 4 | 0.00 | 10.82 | 12.18 | 7.58 |
| Corylus colurna L. | 0.92 | 0.00 | 0.00 | 1.98 | 4 | 0 | 0 | 8 | 4.93 | 0.00 | 0.00 | 11.13 |
| Crataegus songarica K.Koch | 1.28 | 0.00 | 0.00 | 0.00 | 4 | 0 | 0 | 0 | 5.18 | 0.00 | 0.00 | 0.00 |
| Ficus hispida L. | 0.00 | 1.36 | 4.08 | 1.32 | 0 | 4 | 12 | 4 | 0.00 | 8.89 | 12.43 | 5.86 |
| Ficus palmata Forssk. | 0.00 | 0.00 | 0.00 | 1.44 | 0 | 0 | 0 | 4 | 0.00 | 0.00 | 0.00 | 5.95 |
| Fraxinus excelsior L. | 0.00 | 1.28 | 1.28 | 1.56 | 0 | 4 | 4 | 4 | 0.00 | 8.79 | 6.37 | 6.05 |
| Juglans regia L. | 5.92 | 2.96 | 8.88 | 3.48 | 8 | 4 | 12 | 4 | 12.69 | 10.77 | 16.45 | 7.61 |
| Juniperus communis L. | 3.84 | 0.00 | 0.00 | 2.88 | 12 | 0 | 0 | 8 | 12.92 | 0.00 | 0.00 | 11.91 |
| Litsea glutinosa (Lour.) C.B.Rob. | 0.00 | 0.00 | 0.00 | 1.12 | 0 | 0 | 0 | 4 | 0.00 | 0.00 | 0.00 | 5.69 |
| Morus alba L. | 0.00 | 0.00 | 2.80 | 1.00 | 0 | 0 | 8 | 4 | 0.00 | 0.00 | 9.50 | 5.60 |
| Picea smithiana (Wall.) Boiss | 11.76 | 11.76 | 19.60 | 12.72 | 12 | 12 | 20 | 12 | 18.40 | 31.34 | 36.04 | 21.98 |
| Pinus wallichiana A.B.Jacks | 57.00 | 27.36 | 29.64 | 18.24 | 100 | 48 | 52 | 48 | 107.42 | 97.89 | 73.07 | 58.72 |
| Platanus orientalis L. | 0.00 | 0.00 | 0.00 | 1.84 | 0 | 0 | 0 | 4 | 0.00 | 0.00 | 0.00 | 6.28 |
| Populus ciliata Wall. ex Royle | 0.00 | 0.00 | 0.00 | 3.04 | 0 | 0 | 0 | 8 | 0.00 | 0.00 | 0.00 | 12.04 |
| Quercus floribunda Lindl. ex A.Camus | 0.00 | 0.00 | 5.04 | 1.96 | 0 | 0 | 12 | 4 | 0.00 | 0.00 | 1.68 | 6.38 |
| Quercus ilex L. | 0.00 | 0.00 | 1.44 | 4.64 | 0 | 0 | 4 | 8 | 0.00 | 0.00 | 6.51 | 10.63 |
| Quercus semecarpifolia Sm. | 0.00 | 2.88 | 0.00 | 0.00 | 0 | 8 | 0 | 0 | 0.00 | 13.62 | 0.00 | 0.00 |
| Rhododendron arboreum Sm. | 1.68 | 1.68 | 5.04 | 3.56 | 4 | 4 | 12 | 4 | 5.48 | 9.26 | 13.23 | 7.67 |
| Robinia pseudoacacia L. | 0.00 | 0.00 | 0.00 | 2.28 | 0 | 0 | 0 | 4 | 0.00 | 0.00 | 0.00 | 6.64 |
| Taxus wallichiana Zucc. | 3.40 | 0.00 | 0.00 | 0.00 | 4 | 0 | 0 | 0 | 6.65 | 0.00 | 0.00 | 0.00 |
| Trema orientale (L.) Blume | 0.00 | 0.00 | 0.00 | 6.96 | 0 | 0 | 0 | 8 | 0.00 | 0.00 | 0.00 | 15.22 |
| Ulmus wallichiana Planch. | 0.00 | 0.00 | 3.36 | 3.04 | 0 | 0 | 12 | 4 | 0.00 | 0.00 | 11.82 | 7.25 |
| Raunkiaer’s Life Forms | Number of Plant Taxa | Biological Spectra (%) of Study Area | Raunkiaer’s Normal Worldwide Spectra (%) | Deviation = Raunkiaer’s Normal Spectra–Biological Spectra (%) of Study Area | |
|---|---|---|---|---|---|
| PH * | 65 * | 19.82 | 46.00 | 26.18 | |
| mcPH | 17 | 5.18 | 0.00 ** | –5.18 | |
| mgPH | 18 | 5.49 | 3.00 | –2.49 | |
| msPH | 11 | 3.35 | 28.00 | 24.65 | |
| nnPH | 19 | 5.79 | 15.00 | 9.21 | |
| CH | 91 | 27.74 | 9.00 | –18.74 | |
| GE | 37 | 11.28 | 6.00 | –5.28 | |
| HC | 46 | 14.02 | 26.00 | 11.98 | |
| TH | 82 | 25.00 | 13.00 | –12.00 | |
| L | 6 | 1.83 | 0.00 ** | –1.83 | |
| Ep | 1 | 0.30 | 0.00 ** | −0.30 | |
| Total | 328 | 100.00 | 100.00 | - | |
| Raunkiaer’s Life Forms | Number of Plant Taxa | Percentage (%) |
|---|---|---|
| LP | 27 | 8.23 |
| MS | 80 | 24.39 |
| MI | 72 | 21.95 |
| MG | 10 | 3.05 |
| NP | 139 | 42.38 |
| Total | 328 | 100.00 |
| Plant Groups as APG IV (Voucher Number) | Habit | Life Span | Phenology Period | Habitat | Raunkiaer′s Life Forms | Raunkiaer′sSub-Types | LeafSpectra | Conservation Status (IUCN) | Specific Distribution | Native (N)/Alien (A) |
|---|---|---|---|---|---|---|---|---|---|---|
| Magnoliids | ||||||||||
| Laurales Juss. ex Bercht. & J.Presl | ||||||||||
| Lauraceae Juss. | ||||||||||
| Litsea glutinosa (Lour.) C.B.Rob. (54,761) | T | P | Sep–Oct | Sparse forests and thickets | PH | msPH | MI | LC | Himalayas | N |
| Monocots | ||||||||||
| Acorales Mart. | ||||||||||
| Acoraceae Mart. | ||||||||||
| Acorus calamus L. (56,255) | H | P | Jul–Sep | River Banks | GE | NP | LC | North America | A | |
| Alismatales R.Br. ex Bercht & J.Presl | ||||||||||
| Araceae Juss. | ||||||||||
| Arisaema flavum (Forssk.) Schott. (56,347) | H | P | Jun–Jul | Field margins | GE | MS | NE | Asia | N | |
| Arisaema jacquemontii Blume (57,631) | H | P | Apr–Jun | Open forests | GE | MS | LC | North America, Eurasia | A | |
| Dioscoreales Mart | ||||||||||
| DioscoreaceaeR.Br | ||||||||||
| Dioscorea bulbifera L. (57,667) | S | A | Jul–Sep | Mixed forests | L | NP | NE | Himalayas | N | |
| Liliales Perleb | ||||||||||
| Liliaceae Juss. | ||||||||||
| Fritillaria cirrhosa D.Don (56,291) | P | H | Aug–Oct | Alpine thickets and scrubs | HC | LP | NE | Asia | N | |
| Asparagales Link | ||||||||||
| Orchidaceae Juss. | ||||||||||
| Calanthe tricarinata Lindl. (57,667) | H | P | Apr–Jun | Grassy slopes | GE | NP | NE | Himalayas | N | |
| Dactylorhiza hatagirea (D.Don) Soo (56,210) | H | P | Jul–Sep | Alpine scrubs and thickets | GE | NP | NE | Indian subcontinent | N | |
| Epipactis helleborine (L.) Crantz (57,602) | H | P | Jul–Sep | Moist shady places | GE | MI | LC | Asia and Africa | A | |
| Iridaceae Juss. | ||||||||||
| Iris hookeriana Foster (55,708) | H | P | Apr–Jun | Grassy slopes | GE | NP | NE | Himalayas | N | |
| Iris kashmiriana Baker (55,721) | H | P | Jun–Jul | Grassy areas, graveyards | GE | NP | NE | Himalayas | N | |
| Amaryllidaceae J.St.Hil | ||||||||||
| Allium humile Kunth (56,229) | H | P | Jun–Jul | Alpine thickets and rocky crevices | GE | MI | NE | Himalayas | N | |
| Allium stracheyi Baker (56,228) | H | P | Jun–Jul | Alpine thickets and rocky crevices | GE | MI | NE | Himalayas | N | |
| Asparagaceae Juss. | ||||||||||
| Asparagus filicinus Buch.–Ham. ex D.Don (54,548) | H | S | May–Jul | Moist shady places | PH | nnPH | MI | DD | Himalayas | N |
| Maianthemum purpureum (Wall.) La Frankie (56,231) | H | P | Jun–Jul | Forest Thickets and slopes | CH | MS | NE | Himalayas | N | |
| Polygonatum cirrhifolium (Wall.) Royle (56,230) | H | P | Jun–Aug | Forest Thickets and margins | GE | LP | NE | Himalayas | N | |
| Polygonatum verticillatum (L.) All. (56,224) | H | P | Jun–Aug | Forest Thickets and margins | GE | LP | NE | Eurasia | A | |
| Poales Small | ||||||||||
| Cyperaceae Juss. | ||||||||||
| Carex nivalis Boott (55,767) | P | H | May–Jun | Hill slopes | HC | MI | NE | Eurasia | A | |
| Cyperus niveus Retz. (56,348) | H | A | Sep–Oct | Stream sides | TH | MI | NE | Asia | N | |
| Cyperus rotundus L. (56,408) | H | A | May–Jun | Grasslands | TH | MI | LC | Africa and Europe | A | |
| Poaceae Barnhart | ||||||||||
| Alopecurus arundinaceus Poir. (56,319) | H | P | Jul–Sep | Grasslands and slopes | HC | NP | LC | Eurasia | A | |
| Arthraxon lanceolatus (Roxb.) Hochst. (56,336) | H | A | Jul–Sep | Rocky surfaces | TH | NP | LC | Eurasia | A | |
| Bothriochloa ischaemum (L.) Keng (56,337) | H | A | May–Jun | Grasslands | TH | MI | NE | Indian subcontinent | N | |
| Capillipedium parviflorum (R.Br.) Stapf (57,651) | H | P | Jul–Sep | Mountain slopes | TH | NP | NE | Europe | A | |
| Cenchrus flaccidus (Griseb.) Morrone (56,421) | H | A | May–Jun | Waste areas | HC | MS | LC | South Africa | A | |
| Chrysopogon fulvus (Spreng.) Chiov. (56,317) | H | A | Jun–Jul | Moist shady places | TH | MS | NE | Asia | N | |
| Chrysopogon gryllus (L.) Trin. (56,339) | H | P | Aug–Sep | Moist shady places | HC | MS | NE | Eurasia | A | |
| Cynodon dactylon (L.) Pers. (56,260) | H | P | Sep–Dec | Road sides open areas | HC | NP | NE | Africa | A | |
| Echinochloa crus–galli (L.) P.Beauv (56,403) | H | P | May–Jun | Forest margins and road sides | HC | NP | LC | South Africa | A | |
| Elymus semicostatus (Steud.) Melderis (56,432) | H | P | Jul–Aug | Mountain slopes | HC | NP | NE | Indian subcontinent | N | |
| Isachne himalaica Hook.f. (65,322) | H | P | May–Jun | Moist, swampy areas | HC | NP | NE | Himalayas | N | |
| Melinis minutiflora P.Beauv. (56,323) | H | P | Jul–Aug | Field margins and road sides | HC | MS | NE | Africa | A | |
| Miscanthus nepalensis (Trin.) Hack. (56,326) | H | P | Aug–Sep | Mountain slopes and valleys | HC | MS | NE | Himalayas | N | |
| Panicum virgatum L. (56,329) | H | P | Jul–Aug | Field sides | TH | NP | LC | NorthAmerica | A | |
| Poa annua L. (56,304) | H | A | Apr–Jun | Moist shady places | TH | MS | LC | SouthAmerica | A | |
| Polypogon fugax Nees ex. Steud. (56,328) | H | A | Jul–Aug | Moist shady places in forests | TH | NP | NE | SouthAmerica | A | |
| Saccharum fillifolium Steud. (56,309) | H | P | Apr–Jun | Grasslands | TH | NP | NE | Himalayas | N | |
| Saccharum spontaneum L. (56,318) | H | P | Jul–Aug | Hill slopes | HC | NP | LC | Indian subcontinent | N | |
| Setaria italica (L.) P.Beauv. (56,330) | H | P | Jul–Aug | Slopes and Waste lands | TH | MS | NE | Africa | A | |
| Setaria viridis (L.) P.Beauv. (56,331) | H | P | May–Jun | Roadsides and field margins | TH | LP | NE | Asia | N | |
| II. Eudicots | ||||||||||
| Ranunculales Juss. ex Bercht. & J. Presl | ||||||||||
| Papaveraceae Juss. | ||||||||||
| Corydalis diphylla Wall. (57,665) | H | P | Apr–Jun | Steam sides, shady places | GE | NP | NE | Himalayas | N | |
| Corydalis vaginans Royle (57,665) | H | P | Apr–Jun | Stream sides, rock crevices | GE | NP | NE | Himalayas | N | |
| Corydalis govaniana Wall. (54,515) | H | P | Apr–Jun | Forest understories, moist places | GE | NP | NE | Himalayas | N | |
| Fumaria indica (Hausskn.) Pugsley (56,374) | H | P | Mar–Jun | Field margins and thickets | TH | LP | NE | Himalayas | N | |
| Papaver guilelmi–waldemarii (Klotzsch) Christenh. & Byng (55,715) | H | P | Jul–Sep | Rocky crevices | CH | MS | NE | Himalayas | N | |
| Berberidaceae Juss. | ||||||||||
| Berberis aristata DC. (56,315) | S | P | Apr–Jun | Open waste lands | PH | mcPH | NP | LC | Himalayas | N |
| Berberis jaeschkeana C.K.Schneid (57,637) | S | P | Apr–Jun | Forest Thickets, Wastelands | PH | mcPH | NP | NE | Himalayas | N |
| Berberis lycium Royle (54,312) | S | P | Apr–Jun | Forest margins, open waste lands | PH | mcPH | NP | LC | Himalayas | N |
| Podophyllum hexandrum Royle (56,285) | H | P | Jun–Jul | Alpine region open areas | HC | MI | NE | Asia | N | |
| Ranunculaceae Juss. | ||||||||||
| Aconitum heterophyllum Wall. ex Royle (55,769) | H | P | Jul–Sep | Alpine thickets and scrubs | CH | NP | EN | Indian subcontinent | N | |
| Aconitum lethale Griff (55,751) | H | P | Jul–Sep | Alpine thickets and scrubs | CH | NP | NE | Indian subcontinent | N | |
| Actaea spicata L. (56,297) | H | P | Jul–Aug | Moist shady places in forests | CH | MS | LC | Europe | A | |
| Anemonastrum obtusilobum (D.Don) Mosyakin (57,645) | H | P | Apr–June | Forest margins and alpine meadows | CH | MS | NE | Himalayas | N | |
| Anemonastrum polyanthes (D.Don) Holub (55,732) | H | P | Jun–Jul | Steep slopes near glaciers | CH | MS | NE | Himalayas | N | |
| Anemonstarum tetrasepalum (Royle) Holub (54,513) | H | P | Jun–Jul | Steep slopes, alpine scrubs | CH | MS | NE | Himalayas | N | |
| Aquilegia pubiflora Wall. ex Royle (54,766) | H | P | Apr–Jun | Forests, shady places | HC | NP | NE | Himalayas | N | |
| Caltha alba Cambess (55,771) | H | P | Jul–Sep. | Stream sides | CH | NP | NE | Himalayas | N | |
| Caltha palustris L. (54,528) | H | P | Jul–Sep | Stream sides | CH | MS | LC | Himalayas | N | |
| Clematis barbellata Edgew. (57,626) | S | P | Apr–Jun | Forests, Grassy slopes | L | NP | NE | Himalayas | N | |
| Delphinium denudatum Wall. ex Hook.f. & Thomson (54,789) | H | P | May–Jun | Hill slides, forest margins | TH | MS | NE | Himalayas | N | |
| Delphinum cashmerianum Royle (54,508) | H | P | Jul–Sep | Alpine scrubs and thickets | TH | MS | NE | Indian subcontinent | N | |
| Hepatica falconeri (Thomson) Steward (54,362) | H | P | Apr–Jun | Shady places in forests | CH | MS | NE | Indian subcontinent | N | |
| Ranunculus distans D.Don (54,380) | H | P | Jul–Oct | Stream sides, slopedareas | GE | NP | NE | Asia | N | |
| Ranunculus bulbosus L. (54,787) | H | P | Apr–Jun | Disturbed woods, waste lands | GE | NP | NE | Himalayas | N | |
| Ranunculus hirtellus Royle (56,218) | H | P | Apr–Jul | Forest margins and alpine meado3ews | CH | MS | NE | Himalayas | N | |
| Thalictrum foliolosum DC. (56,217) | S | P | Jun–Aug | Alpine thickets and scrubs | CH | MI | NE | Indian subcontinent | N | |
| Thalictrum virgatum Hook.f. & Thomson (56,314) | S | P | Jun–Aug | Forrest margins | CH | MI | NE | Himalayas | N | |
| Proteales Juss. ex Bercht. & J.Presl | ||||||||||
| Platanaceae T.Lestib | ||||||||||
| Platanus orientalis L. (57,677) | T | P | Apr–May | Open areas village sides | PH | mgPH | MS | VU | Europe | A |
| III. CORE EUDICOTS | ||||||||||
| SUPER–ROSIDS | ||||||||||
| Saxifragales Bercht. & J. Presl | ||||||||||
| Hamamelidaceae R.Br. | ||||||||||
| Parrotiopsis jacquemontiana (Decne.) Rehder (7632) | S | P | Apr–Jun | Dense forests | PH | msPH | NP | LC | Central America | A |
| Saxifragaceae Juss. | ||||||||||
| Bergenia ciliata (Haw.) Sternb.(54,359) | H | P | Apr–Jun | Rocky crevices | CH | NP | LC | SouthAmerica | A | |
| Saxifraga cernua L. (54,542) | H | P | Aug–Sep | Stream sides in alpine regions | CH | MI | NE | North America | A | |
| Saxifraga parnassifolia D.Don (55,724) | H | P | Aug–Sep | Stream sides in alpine regions | CH | NP | NE | Eurasia | A | |
| Saxifraga sibirica L. (55,472) | H | P | Aug–Sep | Stream sides in alpine regions | CH | NP | LC | Himalayas | N | |
| Crassulaceae J.St.–Hil. | ||||||||||
| Hylotelephiu mewersii (Ledeb.) H.Ohba (55,740) | H | P | Jul–Sep | Forests, rock crevices | CH | NP | NE | Afghanistan to Siberia | A | |
| Rhodiola bupleuroides (Wall.ex Hook.f. & Thomson) S.H.Fu. (55,797) | H | P | Jul–Aug | Forest Thickets, Rocky slopes | CH | LP | NE | Himalayas | N | |
| Rhodiola himalensis (D.Don) S.H.Fu (56,204) | H | P | Jul–Sep | Rocky crevicesin alpine areas | CH | LP | NE | Himalayas | N | |
| Rhodiola wallichiana (Hook.) S.H.Fu (55,797) | H | P | Jul–Aug | Forest thickets and Rocky scrubs | CH | LP | NE | Himalayas | N | |
| Rosularia adenotricha (Wall. ex Edgew) C.–A.Jansson (57,666) | H | P | May–Jun | Rocky crevices | TH | MI | NE | Himalayas | N | |
| Sedum orades (Decne.) Raym.–Hamet (55,723) | H | P | Jul–Sep | Rocky crevices | TH | MI | NE | Himalayas | N | |
| Rosids | ||||||||||
| Fabales Bromhead | ||||||||||
| Fabaceae Lindl. | ||||||||||
| Indigofera heterantha Wall. ex Brandis (57,625) | S | P | Apr–Jun | Forest margins waste lands | PH | mcPH | MS | LC | Himalayas | N |
| Lotus corniculatus L. (RRLH57635) | H | P | Apr–Jun | Alpine meadows | CH | MS | LC | Asia | N | |
| Medicago lupulina L. (56,295) | H | P | Jul–Sep | Woodland marginsriverbanks | TH | NP | LC | Europe | A | |
| Oxytropis lapponica (Wahlenb.) J.Gay (56,406) | H | P | Jun–Aug | Alpine meadows | GE | MI | NE | Eurasia | A | |
| Phyllodium elegans (Lour.) Desv. (57,633) | S | P | Jul–Sep | Field margins, waste lands | PH | nnPH | MI | NE | Eurasia and North America | A |
| Robinia pseudoacacia L. (57,638) | T | P | Apr–Jun | Waste lands, road sides | PH | mgPH | MS | LC | Europe | A |
| Thermopsis barbata Benth. (54,531) | H | P | Jul–Aug | Alpine high areas | CH | MS | NE | Eurasia | A | |
| Trifolium pratense L. (57,641) | H | P | Apr–May | Grasslands meadows | HC | NP | LC | Himalayas | N | |
| Rosales Bercht. & J.Presl | ||||||||||
| Rosaceae Juss. | ||||||||||
| Agrimonia pilosa Ledeb. (57,622) | H | P | Jul–Sep | Inside forests | TH | MS | NE | Himalayas | N | |
| Cotoneaster micrphyllus Wall. ex Lindl. (54,527) | S | P | May–Jun | Alpine rich regions | PH | mcPH | NP | NE | Himalayas | N |
| Crataegus songarica K. Koch (54,737) | T | P | Apr–Jun | Field margins | PH | mgPH | NP | LC | Himalayas | N |
| Filipendula vestita (Wall. ex G.Don) Maxim. (55,738) | H | P | May–Aug | Alpine meadows and scrubs | CH | NP | NE | Himalayas | N | |
| Fragaria nubicola (Lindl. ex Hook.f.) Lacaita (55,768) | H | P | May–Aug | Meadows, forest valleys | TH | MS | NE | Himalayas | N | |
| Geumelatum Wall. ex G.Don (55,707) | H | P | Apr–Jun | Inside forests | CH | NP | NE | Indo–Malayan region | N | |
| Geum urbanum L. (54,725) | H | P | Jul–Sep | Forest disturbed areas | CH | NP | LC | Himalayas | N | |
| Potentilla argyrophylla Wall. ex Lehm. (55,786) | H | P | May–Jun | Forest thickets and meadows | CH | LP | NE | Himalayas | N | |
| Potentilla atrosanguinea G.Lodd. (54,538) | H | P | May–Jun | Forest thickets and meadows | CH | NP | NE | Europe and America | ||
| Potentilla indica (Andrews) Th.Wolf (54,340) | H | P | Jun–Aug | Forest scrubs and margins | CH | NP | NE | Himalayas | N | |
| Potentilla nepalensis Hook. (54,759) | H | P | Jul–Aug | Forest margins and thickets | CH | LP | NE | Asia and North America | A | |
| Potentilla reptans L. (54,355) | H | P | Apr–Jun | Forest margins and thickets | CH | NP | NE | Egypt and North Africa | A | |
| Prinsepia utilis Royle (54,702) | H | P | Apr–Jun | Waste lands, forest slopes | PH | nnPH | MI | NE | Himalayas | N |
| Rosa moschata Herrm. (54,304) | S | P | May–Jun | Forest margins | PH | nnPH | MS | NE | Himalayas | N |
| Rosa webbiana Wall. ex Royle (54,742) | S | P | May–Jun | Forrest slopes and margins | L | NP | NE | Himalayas | N | |
| Rubus ellipticus Sm. (54,734) | S | P | Apr–May | River sides, sparse forests | PH | nnPH | MS | LC | Himalayas | N |
| Rubus idaeus L. (57,642) | S | P | Apr–Jun | River sides and forest margins | PH | nnPH | NP | LC | Asia | N |
| Rubus niveus Thunb. (57,623) | S | P | May–Jul | River sides and forest | PH | nnPH | NP | NE | Himalayas | N |
| Sibbaldia cuneata Edgew. (55,735) | H | P | Jul–Sep | Alpine scrubs and meadows | CH | NP | NE | Eurasia and Africa | A | |
| Sorbaria tomentosa (Lindl.) Rehder (54,783) | S | P | Jul–Aug | River sides, forest margins | PH | mcPH | MI | NE | Himalayas | N |
| Spiraea canescens D.Don (57,630) | S | P | Jul–Aug | Forest margins, waste lands | PH | mcPH | NP | LC | Himalayas | N |
| Elaeagnaceae Juss | ||||||||||
| Elaeagnus umbellata Thunb. (54,747) | S | P | Apr–Jun | Forest thickets and margins | L | NP | LC | Himalayas | N | |
| Ulmaceae Mirb. | ||||||||||
| Ulmus wallichiana Planch. (57,606) | T | P | Mar–Apr | Temperate forests, village sides | PH | mgPH | MS | UV | Indian subcontinent | N |
| Cannabaceae Mart. | ||||||||||
| Cannabis sativa L. (56,223) | H | P | Aug–Sep | Waste lands, field margins | CH | NP | NE | Asia | N | |
| Celtis australis L. (54,719) | T | P | Mar–Apr | Roadsides, field margins | PH | mgPH | NP | LC | Europe | A |
| Trema orientale (L.) Blume (56,236) | T | P | Jul–Sep | Dry scrubs and open slopes | PH | mgPH | NP | LC | Asia | N |
| Moraceae Gaudich. | ||||||||||
| Ficus hispida L.f. (54,723) | T | P | Jun–Jul | Forest margins | PH | msPH | MS | LC | Himalayas | N |
| Ficus palmata Forssk. (56,205) | T | P | Aug–Nov | Forest slopes | PH | msPH | MS | NE | Himalayas | N |
| Morus alba L. (56,377) | T | P | Apr–May | Waste lands and village sides | PH | msPH | MS | LC | Himalayas | N |
| Urticaceae Juss. | ||||||||||
| Girardinia diversifolia (Link) Friis (54,728) | H | A | Oct–Nov | Disturbed areas, | CH | MS | NE | Himalayas | N | |
| Pilea umbrosa Wedd. ex Blume (54,326) | H | P | Jul–Aug | Shady, moist places | CH | NP | NE | Indian subcontinent | N | |
| Urtica dioica L. (54,501) | H | P | Jun–Sep | Waste lands | HC | MS | NE | Indian Subcontinent | N | |
| Fagales Engl. | ||||||||||
| Fagaceae Dumort. | ||||||||||
| Quercus floribunda Lindl. ex A.Camus (56,305) | T | P | Aug–Oct | Wastelands, forest margins | PH | mgPH | MI | LC | Himalayas | N |
| Quercus ilex L. (54,376) | T | P | Apr–May | Field margins, Waste lands | PH | mgPH | MI | LC | Europe | A |
| Quercus semecarpifolia Sm. (56,235) | T | P | Aug–Oct | Sparse forests and field margins | PH | mgPH | MI | LC | Indian subcontinent | N |
| Juglandaceae DC. ex Prleb. | ||||||||||
| Juglans regia L. (54,336) | T | P | Apr–Sep | Field margins | PH | msPH | MI | LC | Western Himalayas | N |
| Betulaceae Gray | ||||||||||
| Alnus nepalensis D.Don (56,292) | T | P | Jul–Sep | Sparse forests and river banks | PH | mcPH | MI | LC | Himalayas | N |
| Alnus nitida (Spach) Endl. (56,383) | T | P | Sep–Oct | PH | mcPH | MI | LC | Himalayas | N | |
| Betula utilis D.Don (56,289) | T | P | May–Jun | PH | msPH | MI | LC | Himalayas | N | |
| Corylus colurna L. (56,290) | T | P | Mar–Apr | PH | msPH | MS | LC | Asia | N | |
| Cucurbitales Juss. ex Bercht. & J.Presl | ||||||||||
| Cucurbitaceae Juss. | ||||||||||
| Solena heterophylla Lour. (54,790) | S | P | Apr–Jun | Mixed forest thickets | L | MI | NE | Himalayas | N | |
| Datiscaceae Dumort. | ||||||||||
| Datisca cannabina L. (56,374) | S | P | May–Aug | Forest margins and river sides | PH | nnPH | MS | NE | Europe and Himalayas | NMA |
| Celastrales Link | ||||||||||
| Celasteraceae R.Br. | ||||||||||
| Parnassia nubicola Wall. ex Royle (55,709) | H | P | Jul–Sep | Aline meadows and scrubs | CH | MS | NE | Himalayas | N | |
| Oxalidales Juss. ex Bercht. & J.Presl | ||||||||||
| Oxalidaceae R.Br. | ||||||||||
| Oxalis acetosella L. (54,328) | H | P | Jul–Sep | Forest shady places | TH | MS | NE | Himalayas | N | |
| Malpighiales Juss. ex Bercht. & J. Presl | ||||||||||
| Hypericaceae Juss. | ||||||||||
| Hypericum elodeoides Choisy (54,740) | H | P | Jul–Sep | Damp areas in forests | PH | nnPH | NP | NE | Himalayas | N |
| Hypericum hookerianum Wight & Arn. (54,723) | S | P | Apr–Jun | Forest margins | PH | nnPH | NP | NE | North America | A |
| Hypericum perforatum L. (57,662) | S | P | Jul–Sep | Forest scrubs, open areas | PH | nnPH | NP | LC | Himalayas | N |
| Violaceae Batsch | ||||||||||
| Viola canescens Wall. (54,376) | H | P | Mar–Apr | Shady, moist places in forests | TH | NP | NE | Eurasia | A | |
| Salicaceae Mirb. | ||||||||||
| Populus ciliata Wall. ex Royle (56,248) | T | P | May–Jun | Road sides and waste lands | PH | mgPH | MS | LC | Himalayas | N |
| Salix daltoniana Andersson (54,711) | T | P | May–Jun | Forest thickets and slopes | PH | msPH | NP | NE | Europe | A |
| Euphorbiaceae Juss. | ||||||||||
| Euphorbia obovata Decne. (57,620) | H | P | Jul–Sep | Slopy grasslands | TH | NP | EN | Himalayas | N | |
| Euphorbia prolifera Buch.–Ham. ex D.Don. (57,620) | H | P | Jun–Aug | Grasslands and forest margins | TH | NP | NE | Himalayas | N | |
| Euphorbia wallichii Hook.f. (56,394) | H | P | Jun–Aug | Grasslands and forest margins | TH | NP | NE | Himalayas | N | |
| Gerianales Juss. ex Bercht. & J.Presl | ||||||||||
| Geraniaceae Juss. | ||||||||||
| Geranium mascatense Boiss. (54,748) | H | P | Jul–Sep | Forest thickets and scrubs | TH | NP | NE | Indian subcontinent | N | |
| Geranium wallichianum D.Don ex Sweet (54,334) | H | P | Jul–Sep | Forest thickets and scrubs | TH | NP | LC | IndianSubcontinent | N | |
| Melianthaceae Horan. | ||||||||||
| Trillium govanianum Wall. ex D.Don (55,710) | H | P | May–Jun | Forest thickets and scrubs | GE | MS | EN | Himalayas | N | |
| Myrtales Juss. ex Bercht. & J.Presl | ||||||||||
| Onagraceae Juss. | ||||||||||
| Oenothera biennis L. (55,789) | H | P | Jul–Sep | Open and disturbed areas | TH | MS | NE | Himalayas | N | |
| Oenothera rosea L’Hér. ex Aiton (57,663) | H | P | Jul–Sep | Field margins, open areas | TH | NP | NE | Europe | A | |
| Sapindales Juss. ex Bercht. & J.Presl | ||||||||||
| Sapindaceae Juss. | ||||||||||
| Acer caesium Wall. ex Brandis (54,717) | T | P | Apr–Jun | Mixed coniferous forests | PH | mgPH | MG | LC | Himalayas | N |
| Aesculus indica (Wall.ex Cambess) Hook. (57,672) | T | P | Apr–Jun | Mixed coniferous forests | PH | mgPH | MG | LC | Himalayas | N |
| Rutaceae Juss. | ||||||||||
| Skimmia laureola (DC.) Decne. (54,526) | S | P | Apr–May | Mixed coniferous forests | PH | mcPH | MG | NE | Himalayas | N |
| Malvales Juss. ex Bercht. & J.Presl | ||||||||||
| Malvaceae Juss. | ||||||||||
| Hibiscus syriacus L. (57,673) | S | P | Jul–Sep | Hill slides along stream sides | PH | mcPH | NP | NE | Himalayas | N |
| Malva cachemiriana (Cambess.) Alef. (56,241) | H | P | Jul–Sep | Field margins and Waste lands | TH | MS | NE | Himalayas | N | |
| Malva neglecta Wallr. (57,650) | H | P | Jul–Sep | Disturbed areas | TH | LP | LC | Asia | N | |
| Brassicales Bromhead | ||||||||||
| Brassicaceae Burnett | ||||||||||
| Arabis alpina L. (54,307) | H | A | May–Jul | Road sides, slopes | CH | NP | NE | Eurasia | A | |
| Arabis amplexicaulis Edgew. (54,369) | H | A | Jun–Jul | Forest and field margins | TH | NP | NE | Eurasia | A | |
| Barbarea intermedia Boreau (56,221) | H | P | Jul–Aug | Open areas and forest margins | TH | MI | NE | Europe | A | |
| Capsella–bursa pastoris (L.) MediK. (56,354) | H | P | Mar–Apr | Field margins and open lands | TH | MI | LC | Eurasia and Africa | A | |
| Cardamine impatiens L. (56,226) | H | A | May–Jul | Field margins and Wastelands | HC | MI | NE | Eurasia | A | |
| Crucihimalaya stricta (Cambess.) Al Shebaz, O′Kane & R.A.Price (57,601) | H | A | Apr–Jun | Forest margins, grassy slopes | TH | NP | NE | Indian subcontinent | N | |
| Draba nuda (Bél.) Al–Shehbaz & M.Koch (54,324) | H | A | Apr–Jun | Stream sides, moist places | TH | NP | NE | Himalayas | N | |
| Lepidium virginicum L. (57,610) | H | A | Jul–Sep | Forest margins and filed edges | TH | NP | NE | NorthAmerica | A | |
| Nasturtium officinale W.T.Aiton (55,798) | H | P | Apr–Sep | Moist, damp areas along streams | TH | NP | LC | Asia and Africa | N | |
| Rorippa islandica (Oeder) Borbás (54,356) | H | A | Apr–Jun | Shady crevices, river sides | TH | NP | LC | Himalayas | N | |
| Super Asterids | ||||||||||
| Caryophyllales Juss. ex Bercht. & J.Presl | ||||||||||
| Polygonaceae Juss. | ||||||||||
| Bistorta affinis (D.Don) Greene (55,718) | H | P | Jul–Sep | Alpine scrubs and scrubs | CH | LP | NE | Himalayas | N | |
| Bistorta amplexicaulis (D.Don) Greene (57,643) | H | P | Jul–Sep | Mountain scrubs, field margins | CH | LP | NE | Africa and SouthAmerica | A | |
| Bistorta vivipara (L.) Delarbre (54,512) | H | P | Jul–Aug | Mountain scrubs | CH | LP | NE | Himalayas | N | |
| Fagopyrum cymosum (Trevir.) Meisn. (55,762) | H | P | Jul–Sep | Field margins and open areas | CH | LP | NE | Himalayas | N | |
| Fagopyrum esculentum Moench (67,644) | H | P | Jul–Sep | Field margins, Waste lands | CH | MS | NE | Indian subcontinent | N | |
| Koenigia rumicifolia (Royle ex Bab.) T.M.Schust. & Reveal (55,734) | H | P | Jul–Sep | Alpine thickets and scrubs | TH | MS | NE | Himalayas | N | |
| Koenigia alpina (All.) T.M. Schust & Reveal (54,766) | H | P | Jul–Aug | Mountain slopes, field margins | TH | NP | NE | Himalayas | N | |
| Oxyria digyna (L.) Hill (56,384) | H | P | May–Aug | Alpine scrubs and thickets | CH | MI | NE | Europe and America | A | |
| Persicaria glabra (Willd.) (56,358) | H | P | Sep–Nov | Moist shady places in forests | CH | MI | LC | Eurasia | A | |
| Persicaria lapathifolia (L.) Delarbre (56,281) | H | A | Aug–Nov | Moist shady places along roadsides | CH | MI | LC | Eurasia | A | |
| Rheum webbianum Royle (55,777) | H | P | Aug–Sep | Alpine scrubs, meadows | GE | MG | NE | Himalayas | N | |
| Rumex dentatus L. (56,357) | H | P | Jun–Jul | Mountain slopes and moist valleys | GE | MS | LC | Asia | N | |
| Rumex hastatus D.Don (54,703) | H | P | Apr–Jun | Dry mountain slopes | CH | MI | NE | Indian subcontinent | N | |
| Rumex nepalensis Spreng. (56,273) | H | P | Jun–Jul | Field margins and waste lands | HC | MS | NE | Asia and Africa | N | |
| Caryophyllaceae Juss. | ||||||||||
| Acanthophyllum cerastioides (D.Don) Madhani & Zarre (54,512) | H | P | Jul–Sep | Rocky slopes in alpine meadows | CH | NP | NE | Himalayas | N | |
| Cerastiumdavuricum Fisch. ex Spreng. (54,314) | H | P | Apr–Jun | Stream sides, damp areas | TH | NP | NE | Europe and NorthAmerica | A | |
| Cerastiumglomeratum Thuill. (54,343) | H | P | May–Jun | Moist damp areas along streams | TH | NP | NE | Europe | A | |
| Silene indica Roxb. ex Otth (55,878) | H | P | Jul–Aug | Mountain scrubs and forest margins | TH | NP | NE | Europe and NorthAmerica | A | |
| Silene vulgaris (Moench) Garcke (57,616) | H | P | Jul–Aug | Mountain scrubs | TH | NP | LC | Europe | A | |
| Amaranthaceae Juss. | ||||||||||
| Achyranthyes aspera L. (54,755) | H | P | Jul–Aug | Forest slopes, waste lands | CH | NP | NE | South America | A | |
| Amaranthus caudatus L. (576,04) | H | A | Jul–Sep | Field margins | TH | MI | NE | Eurasia | ||
| Amaranthus viridis L. (563,95) | H | A | Jun–Aug | Field margins | TH | MI | NE | America | A | |
| Blitum virgatum L. (557,83) | H | A | Jul–Sep | Forest margins | TH | MI | NE | Himalayas | N | |
| Chenopodium album L. (55,770) | H | A | Jul–Sep | Field margins, waste lands | TH | MI | NE | Himalayas | N | |
| Dysphania ambrosioides (L.) Mosyakin & Clemants (56,360) | H | A | Jul–Sep | Field margins and Waste lands | TH | MI | NE | America | A | |
| Phytolaccaceae R.Br. | ||||||||||
| Phytolacca acinosa Roxb (57,619) | H | P | Jul–Aug | Forest understories | GE | MG | NE | America | A | |
| Asterids | ||||||||||
| Ericales Bercht. & J.Presl | ||||||||||
| Balsaminaceae A.Rich. | ||||||||||
| Impatiens balfourii Hook.f. (54,762) | H | P | Jul–Aug | Stream sides, field margins | CH | LP | NE | Australia | A | |
| Impatiens brachycentra Kar. & Kir. (55,785) | H | A | Aug–Sep | Forest margins, along streams | CH | MS | NE | Eurasia and Africa | A | |
| Impatiens edgeworthii Hook.f. (55,782) | H | A | Jul–Sep | Inside forests, moist areas | CH | MS | NE | Eurasia and Africa | A | |
| Polemoniaceae Juss. | ||||||||||
| Polemonium caeruleum L. (55,761) | H | P | Apr–Jun | Moist areas in forests | CH | MS | NE | Asia | N | |
| Primulaceae Batsch ex Borkh. | ||||||||||
| Androsace rotundifolia Hardw. (54,789) | H | P | Apr–Jun | Grassy slopes in the forests | HC | NP | NE | Himalayas | N | |
| Androsace sarmentosa Wall. (56,215) | H | P | Jun–Jul | Rocky surfaces in high alpine regions | HC | NP | NE | Himalayas | N | |
| Primula denticulata Sm. (57,689) | H | P | Apr–Jun | Alpine thickets and moist scrubs | GE | MS | NE | Himalayas | N | |
| Primula matthioli (L.) V.A. Richt. (56,219) | H | P | Jun–Jul | Alpine thickets and moist scrubs | GE | MS | NE | Europe | A | |
| Primula rosea Royle (56,225) | H | P | Jun–Jul | Alpine thickets and moist scrubs | GE | MI | NE | Himalayas | N | |
| Ericaceae Juss. | ||||||||||
| Rhododendron anthopogon D.Don (55,741) | S | P | Apr–Jun | High mountain scrubs | PH | mcPH | NP | NE | Eurasia | A |
| Rhododendron arboretum (54,394) | T | P | Apr–Jun | Mixed temperate forests | PH | msPH | MS | LC | Himalayas | N |
| Rhododendron campanulatum D.Don (55,725) | S | P | May–Jun | High alpine scrubs | PH | mcPH | NP | NE | Eurasia | A |
| Gentianales Juss. ex Bercht. & J.Presl | ||||||||||
| Rubiaceae Juss. | ||||||||||
| Rubia cordifolia L. (57,636) | H | P | Jul–Sep | Sparse forest | HC | NP | NE | Himalayas | N | |
| Gentianaceae Juss. | ||||||||||
| Gentiana carinata (D.Don ex G.Don) Royle ex D.Don (56,208) | H | A | Jul–Aug | Alpine meadows and thickets | HC | NP | NE | Himalayas | N | |
| Gentiana argentea (Royle ex D.Don) Royle ex D.Don (54,332) | H | A | Mar–Apr | Waste lands, grassy slopes | HC | NP | NE | Europe and North America | A | |
| Jaeschkea oligosperma Knobl. (55,701) | H | A | Jul–Sep | Grassy slopes, grasslands | TH | MS | NE | Himalayas | N | |
| Swertia chirayita (Roxb.) H.Karst. (55,796) | H | P | Aug–Sep | Grassy slopes, grasslands | TH | NP | NE | Himalayas | N | |
| Apocynaceae Juss. | ||||||||||
| Vincetoxicum hirundinaria Medik. (57,669) | H | P | May–Jun | Thickets, forest margins | CH | MS | NE | Himalayas | N | |
| Boraginales Juss. ex Bercht. & J.Presl | ||||||||||
| Boraginaceae Juss. | ||||||||||
| Arnebia benthamii (Wall. ex G.Don) I.M.Jhonst. (56,240) | H | P | Jun–Jul | Mountain slopes, rock cervices | CH | MI | NE | Himalayas | N | |
| Cynoglossum lanceolatum Forssk. (55,779) | H | P | Jun–Aug | Forest margins and thickets | TH | NP | NE | South Africa | A | |
| Cynoglossum wallichii G.Don (54,792) | H | P | May–Aug | Hill slides, meadows | TH | NP | NE | NorthAmerica | A | |
| Hackelia macrophylla (Brand) I.M.Jhonst. (563,362) | H | P | Jun–Jul | Alpine thickets and scrubs | TH | Mi | NE | Himalayas | N | |
| Hackelia uncinata (Royle ex Benth.) C.E.C Fisch. (56,238) | H | P | Jun–Jul | Alpine thickets and scrubs | TH | MI | NE | Asia | N | |
| Lindelofia longiflora (DC.) Baill. (57,615) | H | P | Apr–Jun | Grasslands and slopes | CH | MI | NE | Asia | N | |
| Myosotis scorpioides L. (54,760) | H | P | May–Jun | Forest margins slopes | HC | MS | NE | Indian subcontinet | N | |
| Trichodesma indicum (L.) Sm. (54,319) | H | P | Mar–Apr | Shady and moist places | TH | NP | NE | Eurasia | A | |
| Solanales Juss. ex Bercht. & J.Presl | ||||||||||
| Convolvulaceae Juss. | ||||||||||
| Ipomoea purpurea (L.) Roth (57,612) | H | P | Jul–Sep | Fields, waste lands | TH | MS | NE | Himalayas | N | |
| Solanaceae Juss. | ||||||||||
| Datura stramonium L. (57,639) | H | P | Jul–Sep | Wastelands, road side | HC | MG | NE | Japan and South Korea | A | |
| Lamiales Bromhead | ||||||||||
| Oleaceae Hoffmanns. | ||||||||||
| Chrysojasminum humile (L.) Banfi (56,247) | S | P | Jul–Sep | Riversides | PH | mcPH | MI | NE | Indian subcontinent | N |
| Fraxinus excelsior L. (54,725) | T | P | Apr–Jun | Field margins, roadsides | PH | mgPH | NP | NT | Himalayas | N |
| Jasminum grandiflorum L. (54,767) | S | P | Jul–Sep | Forest margins, slopy areas | L | MI | NE | Eurasia | A | |
| Syringa emodi Wall. ex Royle (56,284) | S | P | Sep–Oct | Alpine scrubs and thickets | PH | nnPH | MS | NE | Himalayas | N |
| Plantaginaceae Juss. | ||||||||||
| Digitalis lanata Ehrh. (56,288) | H | A | Jun–Aug | Open areas and scrubs | TH | NP | LC | Europe and America | A | |
| Digitalis purpurea L. (57,607) | H | A | May–Jun | Forest margins | TH | NP | LC | Himalayas | N | |
| Lagotis cashmeriana (Royle ex Benth.) Rupr. (55,753) | H | P | Jul–Aug | Alpine thickets and scrubs | HC | NP | NE | Asia | N | |
| Lagotis kunawurensis (Royle ex Benth.) Rupr. (56,222) | H | P | Jul–Aug | Alpine scrubs and meadows | HC | NP | NE | Himalayas | N | |
| Picrorhiza kurroa Royle ex Benth. (55,758) | H | P | Jul–Sep | Alpine scrubs and thickets | CH | NP | EN | Himalayas | N | |
| Plantago himalaica Pilg. (56,284) | H | P | Jul–Aug | Mountain slopes and margins | HC | MI | NE | Indian subcontinent | N | |
| Plantago lanceolata L. (56,261) | H | P | Jul–Aug | Forest thickets and edges | HC | MS | LC | Eurasia | A | |
| Plantago major L. (54,794) | H | P | Jul–Aug | Forest margins, slopes | HC | MS | LC | Himalayas | N | |
| Wulfeniopsis amherstiana (Benth.) (54,741) | H | P | Jul–Aug | Rock crevices and shady areas | CH | NP | NE | Himalayas | N | |
| Scrophulariaceae Juss. | ||||||||||
| Verbascum thapsus L. (57,608) | H | P | Jun–Jul | Mountain slopes | CH | NP | LC | NorthAmerica | A | |
| Acanthaceae Juss. | ||||||||||
| Strobilanthes attenuata (Wall. ex Nees) Jacq. ex Nees (5578) | H | P | Jun–Oct | Forest thickets | HC | LP | NE | Himalayas | N | |
| Lamiaceae Martinov | ||||||||||
| Ajuga integrifolia Buch.–Ham. ex D.Don (54,331) | H | P | May–Jun | Grassy slopes | TH | MS | NE | Himalayas | N | |
| Elsholtzia ciliata (Thunb.) Hyl. (54,779) | H | P | Jul–Oct | Waste lands | HC | MI | NE | India and Australia | N | |
| Elsholtzia fruticosa (D.Don) Rehder (57,681) | H | S | Aug–Oct | Wastelands and forest margins | PH | nnPH | MI | NE | Himalayas | N |
| Isodon coesta (Buch.–Ham. ex D.Don) Kudo (54,553) | S | P | Sep–Oct | Woodland thickets | PH | nnPH | MI | NE | Himalayas | N |
| Isodon japonicus (Burm.f.) H.Hara (54,552) | S | P | Sep–Oct | Woodland thickets | PH | nnPH | MI | NE | Himalayas | N |
| Isodon rugosus (Wall. ex Benth.) Codd (57,629) | S | P | Jul–Oct | Wastelands and forest scrubs | PH | nnPH | MI | NE | Himalayas | N |
| Lamium album L. (54,551) | H | P | Jul–Sep | Forest margins Shady places | TH | MS | LC | Asia | N | |
| Lamium amplexicaule L. (54,303) | H | A | Mar–Apr | Forest margins | TH | MS | NE | Asia | N | |
| Leucas ciliata Benth. (54,780) | H | A | Jul–Oct | Roadsides, stream sides | TH | NP | NE | Indiansubcontinent | N | |
| Mentha arvensis L. (56,354) | H | P | Jul–Sep | Field margins and wastelands | GE | NP | LC | Eurasia | A | |
| Mentha longifolia (L.) L. (56,363) | H | P | Jul–Sep | Field margins and wastelands | GE | NP | LC | Eurasia | A | |
| Nepeta clarkei Hook.f. (54,731) | H | P | May–Aug | Wastelands, field margins | GE | NP | NE | Himalayas | N | |
| Nepeta laevigata (D.Don) Hand.–Mazz. (55,763) | H | P | May–Aug | Road sides, field margins | GE | NP | NE | Himalayas | N | |
| Nepeta lamiopsis Benth. ex Hook.f. (54,766) | H | P | Apr–Jun | Forest margins and thickets | HC | MS | NE | Himalayas | N | |
| Origanum vulgare L. (55,745) | H | P | Jul–Sep | Forest thickets | CH | NP | NE | Eurasia | A | |
| Pimpinella diversifolia DC (56,394) | H | P | Aug–Sep | Mountain scrubs and field margins | CH | MS | NE | |||
| Prunella vulgaris L. (57,634) | H | P | Jul–Aug | Waste lands | CH | NP | NE | Himalayas | N | |
| Salvia hians Royle ex Benth. (55,749) | H | P | Jul–Sep | Alpine thickets near stream sides | CH | NP | NE | Himalayas | N | |
| Salvia nubicola Wall. ex Sweet (57,668) | H | P | Jul–Sep | Grassy slopes | TH | NP | NE | Asia | N | |
| Thymus linearis Benth. (57,611) | H | P | Jul–Aug | Rocky slopes | HC | MI | NE | Eurasia | A | |
| Thymus serpyllum L. (57,671) | H | P | May–Jun | Rocky slopes | HC | MI | LC | Himalayas | N | |
| Mazaceae Reveal | ||||||||||
| Mazus pumilus (Burm.f.) Steenis (54,777) | H | A | Jul–Sep | Grasslands and thickets | CH | NP | NE | Africa and Asia | A | |
| Mazus surculosus D.Don (RRLH57685) | H | P | Jun–Sep | Forest edges and thickets | CH | NP | NE | South America | A | |
| Orobanchaceae Vent. | ||||||||||
| Pedicularis multiflora Pennell (54,507) | H | A | Jul–Sep | Alpine meadows, slopy areas | GE | NP | NE | Himalayas | N | |
| Pedicularis pectinata Wall. ex Benth. (54,540) | H | A | Jul–Sep | Alpine meadows, slopy areas | GE | MS | NE | Eurasia and America | A | |
| Pedicularis pyramidata Royle ex Benth. (54,796) | H | A | Jul–Sep | Alpine meadows, slopy areas | GE | NP | NE | Himalayas | N | |
| Asterales Link | ||||||||||
| Campanulaceae Juss. | ||||||||||
| Campanula latifolia L. (57,670) | H | P | Jul–Sep | Field margins, grassy areas | CH | LP | NE | Himalayas | N | |
| Campanula pallida Wall. (54,776) | H | P | Jul–Sep | Forest margins and scrubs | CH | MS | NE | SouthAmerica | A | |
| Codonopsis ovate Benth. (55,728) | H | P | Jul–Sep | Alpine thickets and scrubs | CH | MI | NE | NorthAmerica | A | |
| Cyananthus lobatus Wall. ex Benth. (55,736) | H | P | Jul–Sep | Alpine meadows and thickets | CH | NP | NE | Himalayas | N | |
| Asteraceae Bercht. & J.Presl | ||||||||||
| Achillea millefolium L. (55,759) | H | P | Jul–Sep | Grassy slopes and field margins | HC | MS | LC | Himalayas | N | |
| Anaphalis margaritacea(L.) Benth. & Hook.f. (54,799) | H | P | Jul–Sep | Shady areas in forests | CH | NP | NE | Himalayas | N | |
| Anaphalis nepalensis (Spreng.) Hand–Mazz. (54,539) | H | P | Jul–Sep | Forest margins, river banks | CH | NP | NE | Eurasia | A | |
| Anaphalis nubigena DC. (54,341) | H | P | Jul–Sep | Forest margins, river banks | CH | NP | NE | Himalayas | N | |
| Anaphalis triplinervis Sims ex C.B.Clarke (55,792) | H | P | Jul–Sep | Forest margins, river banks | CH | NP | NE | Himalayas | N | |
| Aster albescens (DC.) Wall. ex Hand.–Mazz. (54,546) | H | P | Jun–Aug | Alpine scrubs and thickets | TH | MI | NE | Asia | N | |
| Aster flaccidus Bunge (56,293) | H | A | Jun–Jul | High alpine regions | TH | MI | NE | Himalayas | N | |
| Aster falconeri (C.B.Clarke) Hutch. (56,293) | H | A | Jun–Jul | High alpine regions | TH | MI | NE | Eurasia | A | |
| Aster himalaicus C.B.Clarke (55,730) | H | P | Jul–Sep | High grasslands | TH | MI | NE | Europe | A | |
| Artemisia maritima L. (54,330) | H | P | Jul–Sep | Rocky areas, wasteland | CH | NP | NE | Europe | A | |
| Artemisia roxburghiana Besser (54,544) | H | P | Jul–Sep | Forest slopes, waste lands | CH | NP | NE | Himalayas | N | |
| Artemisia vestita Wall. ex Besser (55,799) | H | P | Jul–Sep | Rocky slopes, grasslands | CH | NP | NE | Eurasia Africa | A | |
| Carduus edelbergii Rech. f. (55,704) | H | P | Jul–Sep | Alpine thickets | TH | NP | NE | Indian subcontinent | N | |
| Cirsium arvense (L.) Scoop (57,612) | H | P | Apr–Jun | Field margins, waste lands | GE | NP | NE | Europe | A | |
| Cirsium falconeri (Hook. f.) Petr. (54,543) | H | P | Apr–Jun | Forest scrubs, wastelands | GE | NP | NE | Himalayas | N | |
| Dolomiaea macrocephala DC. ex Royle (55,706) | H | P | Jul–Sep | Alpine scrubs and thickets | CH | NP | NE | Himalayas | N | |
| Erigeron annuus (L.) Desf. (54,718) | H | A | Jul–Sep | Grassy slopes and forest margins | TH | MI | NE | SouthAmerica | A | |
| Erigeron bonariensis L. (57,647) | H | A | Jul–Sep | Field margins, grassy slopes | TH | MI | NE | Africa | A | |
| Erigeron multiradiatus (Lindl. ex DC.) Benth. & Hook.f. (55,754) | H | P | Jul–Sep | Forest Thickets and scrubs | TH | MI | NE | Eurasia | A | |
| Galinsoga parviflora Cav. (57,648) | H | A | Jul–Sep | Stream sides | TH | NP | NE | Africa | A | |
| Heliopsis helianthoides (L.) Sweet (55,780) | S | P | Jul–Sep | Field margins, road sides | PH | nnPH | NP | NE | Himalayas | N |
| Hippolytia longifolia (Rech.f.) C.Shih (55,703) | H | P | Jul–Sep | Forest thickets, grassy slopes | CH | MS | NE | Himalayas | N | |
| Inula grandiflora Willd. (55,729) | H | P | Jul–Sep | Moist shady places in forests | CH | MS | NE | Eurasia | A | |
| Inula royleana DC. (55,722) | H | P | Jul–Sep | Steam sides in high areas | CH | MG | NE | Eurasia | A | |
| Jacobaea nudicaulis (Buch.–Ham ex D.Don) B.Nord (57,640) | H | P | Aug–Sep | Grassy slopes and thickets | TH | NP | NE | Asia and North America | N | |
| Jurinea albescens (DC.) N.Garcia, Herrando & Susanna (55,714) | H | P | Jun–Sep | Mountain scrubs and thickets | HC | LP | NE | Himalayas | N | |
| Lactuca dolichophylla Kitam. (54,503) | H | P | June–Sep | Field margins, road sides | TH | LP | NE | Indian subcontinent | N | |
| Ligularia fischeri (Ledeb) Turcz. (54,528) | H | P | Apr–Jun | Stream sides | HC | MG | NE | Indian subcontinent | N | |
| Ligularia jacquemontiana (Decne.) M.A.Rau (55,726) | H | P | Apr–Jun | Stream sides | HC | MG | NE | Himalayas | N | |
| Pseudog naphalium affine (D.Don) Anderb. (54,791) | H | P | May–Jun | Wastelands and forest margins | TH | MI | NE | NorthAmerica | A | |
| Saussurea candolleana (DC.) Sch.Bip. (55,717) | H | P | Aug–Sep | Forest thickets, mountain scrubs | CH | MI | NE | Himalayas | N | |
| Saussurea roylei (DC.) Sch.Bip. (55,781) | H | P | Aug–Sep | Forest margins, alpine scrubs | CH | MS | NE | Himalayas | N | |
| Taraxacum officinale F.Wigg. (54,350) | H | P | Apr–Jun | Grasslands, forests, road sides | TH | NP | LC | NorthAmerica | A | |
| Tussilago farfara L. (54,785) | H | P | Mar–Apr | Forest understories | TH | MG | LC | America | A | |
| Dipsacales Juss. ex Bercht. & J.Presl | ||||||||||
| Adoxaceae E.Mey. | ||||||||||
| Viburnum grandiflorum Wall. ex DC (54,377) | S | P | Apr–Jun | Inside forests | PH | mcPH | NP | NE | Himalayas | N |
| Caprifoliaceae Juss. | ||||||||||
| Dipsacus inermis Wall. (55,776) | H | P | Jul–Sep | Filed margins, waste lands | CH | MI | NE | Asia | N | |
| Leycesteria formosa Wall. (54,715) | S | P | Apr–Jun | River sides | PH | nnPH | NP | NE | Europe | A |
| Lonicera obovata Royle ex Hook.f.& Thomson (54,542) | S | P | May–Jul | Alpine scrubs and thickets | PH | nnPH | MI | NE | Himalayas | N |
| Morina longifolia Wall. ex DC. (55,705) | H | P | Jul–Aug | Alpine scrubs ana slopes | CH | NP | NE | Himalayas | N | |
| Valeriana jatamansi Jones ex Roxb. (54,310) | H | P | Mar–Apr | Forest thickets and margins | GE | MI | NE | Himalayas | N | |
| Valeriana pyrolifolia Decne (57,688) | H | P | Mar–Apr | Forest thickets and margins | GE | MI | NE | Himalayas | N | |
| Apiales Nakai | ||||||||||
| Apiaceae Lindl. | ||||||||||
| Angelica glauca Edgew (54,331) | H | P | Jun–Aug | Alpine thickets and scrubs | TH | MS | EN | Indian subcontinent | N | |
| Bupleurum longicaule Wall. ex DC. (55,719) | H | P | Jul–Sep | Forest thickets grassy slopes | CH | MS | NE | Indian Subcontinent | N | |
| Carum carvi L. (55,778) | H | P | Jul–Sep | Bushy alpine meadows | TH | NP | LC | Eurasia | A | |
| Chaerophyllum reflexum Aitch. (54,797) | H | P | Jul–Sep | Open forests | CH | MS | NE | Himalayas | N | |
| Chaerophyllum villosum Wall. ex DC. (56,355) | H | P | Jul–Sep | Field margins and forest margins | CH | MS | NE | Indian subcontinent | N | |
| Cortia depressa (D.Don) C.Norman (55,731) | H | P | Jul–Sep | Alpine meadows | TH | NP | NE | Himalayas | N | |
| Heracleum candicans Wall. ex DC. (55,711) | H | P | Jul–Sep | Alpine thickets and rocky slopes | HC | MS | NE | Himalayas | N | |
| Hymenidium brunonis (DC.) Lindl. (55,739) | H | P | Jul–Sep | Sparse forests, forest margins | CH | NP | NE | Himalayas | N | |
| Hymenolaena candollei DC. (55,773) | H | P | Jul–Sep | Forest thickets and rocky scrubs | CH | MI | NE | Himalayas | N | |
| Tetrataenium canescens (Lindl.) Manden. (55,756) | H | P | Aug–Sep | Forest margins and scrubs | CH | MS | NE | Africa and Europe | A | |
| Araliaceae Juss | ||||||||||
| Hedera nepalensis K.Koch (54,365) | S | P | Oct–Apr | Forest thickets | PH | mcPH | NP | NE | Himalayas | N |
| Gymnosperms | ||||||||||
| Pinales Gorozh. | ||||||||||
| Pinaceae Spreng ex E.Rudolphi | ||||||||||
| Abies pindrow (Royle ex D.Don) Royle (54,367) | T | P | Mar–Apr | Alpine regions | PH | mgPH | LP | LC | Himalayas | N |
| Cedrus deodara (Roxb. ex D.Don) G.Don (57,645) | T | P | May–Jun | Coniferous forests | PH | mgPH | LP | LC | Himalayas | N |
| Picea smithiana (Wall) Boiss (54,305) | T | P | May–Jun | Coniferous forests | PH | mgPH | LP | LC | Himalayas | N |
| Pinus wallichiana A.B.Jacks. (57,675) | T | P | Mar–Apr | Coniferous forests | PH | mgPH | LP | LC | Himalayas | N |
| Cupressaceae | ||||||||||
| Juniperus communis L. (56,251). | T | P | Sep–Oct | Alpine regions | PH | msPH | LP | LC | Himalayas | N |
| Juniperus polycarpos K.Koch (55,737) | S | P | Sep–Oct | Alpine regions | PH | mcPH | LP | LC | Himalayas | N |
| Taxaceae | ||||||||||
| Taxus wallichiana Zucc. (56,225) | T | P | Aug–Dec | Coniferous forests | PH | mgPH | LP | EN | Himalayas | N |
| LYCOPHYTES AND FERNS | ||||||||||
| Polypodiales Link | ||||||||||
| Aspleniaceae Newman | ||||||||||
| Asplenium dalhousieae Hook. (54,334) | H | P | Oct–Nov | Moist shady places along rocks | HC | MS | NE | Himalayas | N | |
| Asplenium trichomanes L. (54,317) | H | P | Oct–Nov | Moist rocky surfaces | HC | MS | NE | Himalayas | N | |
| Pteridaceae E.D.M Kirchn | ||||||||||
| Adiantum venustum D.Don (54,367) | H | A | Jan–Apr | Stream sides | HC | MI | NE | Himalayas | N | |
| Onychium japonicum (Thumb.) Kunze (54,371) | H | P | Oct–Dec | Moist and damp areas | CH | MS | NE | Himalayas | N | |
| Pteris biaurita L. (54,370) | H | P | Mar–Apr | Forest margins, rocky slopes | HC | NP | NE | Himalayas | N | |
| Pteris cretica L. (54,379) | H | P | Mar–Apr | Forest margins | HC | NP | NE | Himalayas | N | |
| Polypodiaceae J.Presl & C.Presl | ||||||||||
| Polystichum luctuosum (Kuntze) T.Moore (54,335) | H | P | Mar–Apr | Moist shady places | HC | MI | NE | Himalayas | N | |
| Equisetales DC. ex Bercht. & J.Presl | ||||||||||
| Eqisetaceae Michx. ex DC. | ||||||||||
| Equisetum arvense L. (54,322) | H | A | Jan–Apr | Field margins | GE | MI | LC | Indian subcontinent | N | |
| Selaginellales Prantl | ||||||||||
| Selaginellaceae Willk | ||||||||||
| Selaginella eurynota A.Braun (55,774) | H | A | Mar–Apr | Moist shady places along streams | Ep | NP | NE | Himalayas | N |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surmal, O.; Singh, B.; Musarella, C.M. Understanding Species Diversity, Phenology and Environmental Implications of Different Life Forms in Coniferous Forests: A Case Study from Bhallesa Hills of Pir Panjal Mountain, Western Himalaya, India. Forests 2022, 13, 2050. https://doi.org/10.3390/f13122050
Surmal O, Singh B, Musarella CM. Understanding Species Diversity, Phenology and Environmental Implications of Different Life Forms in Coniferous Forests: A Case Study from Bhallesa Hills of Pir Panjal Mountain, Western Himalaya, India. Forests. 2022; 13(12):2050. https://doi.org/10.3390/f13122050
Chicago/Turabian StyleSurmal, Opender, Bikarma Singh, and Carmelo Maria Musarella. 2022. "Understanding Species Diversity, Phenology and Environmental Implications of Different Life Forms in Coniferous Forests: A Case Study from Bhallesa Hills of Pir Panjal Mountain, Western Himalaya, India" Forests 13, no. 12: 2050. https://doi.org/10.3390/f13122050
APA StyleSurmal, O., Singh, B., & Musarella, C. M. (2022). Understanding Species Diversity, Phenology and Environmental Implications of Different Life Forms in Coniferous Forests: A Case Study from Bhallesa Hills of Pir Panjal Mountain, Western Himalaya, India. Forests, 13(12), 2050. https://doi.org/10.3390/f13122050