
Differential Responses of Soil Respiration and Its Components to Nitrogen Application in Urban Forests


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Evaluation of Rs and Its Components
2.4. Sampling and Analysis of Soil
2.5. Statistical Analysis
3. Results
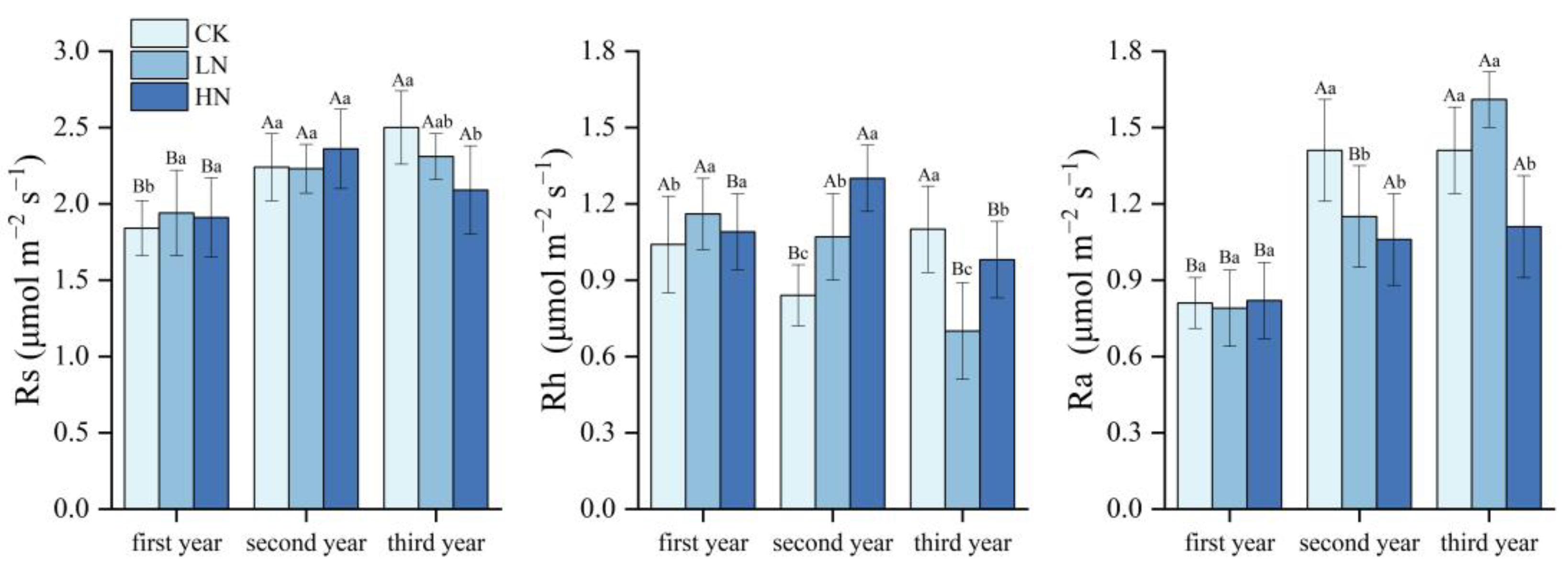
3.1. Effects of N Application on Rs and Its Components
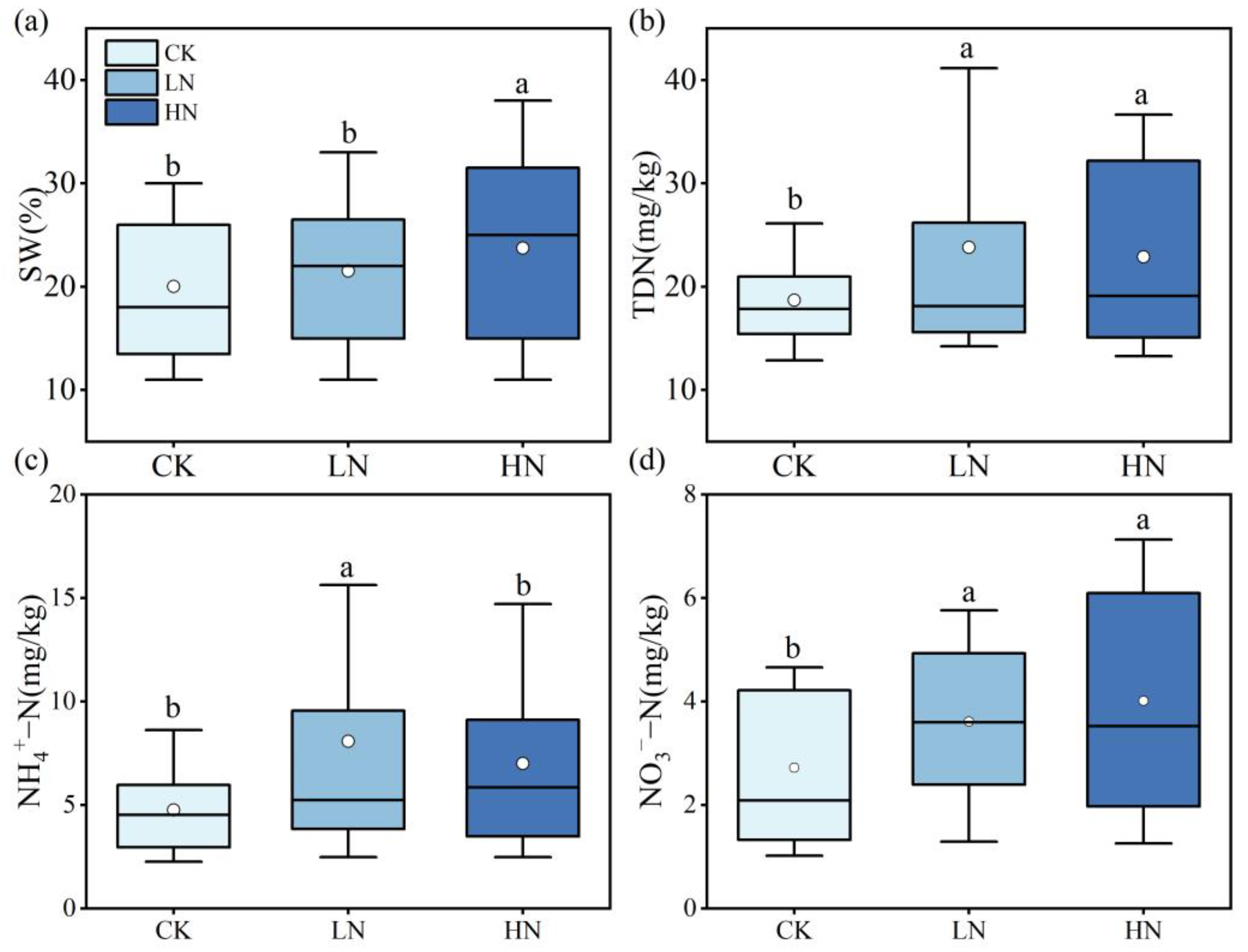
3.2. Changes in the Soil Factors after N Application
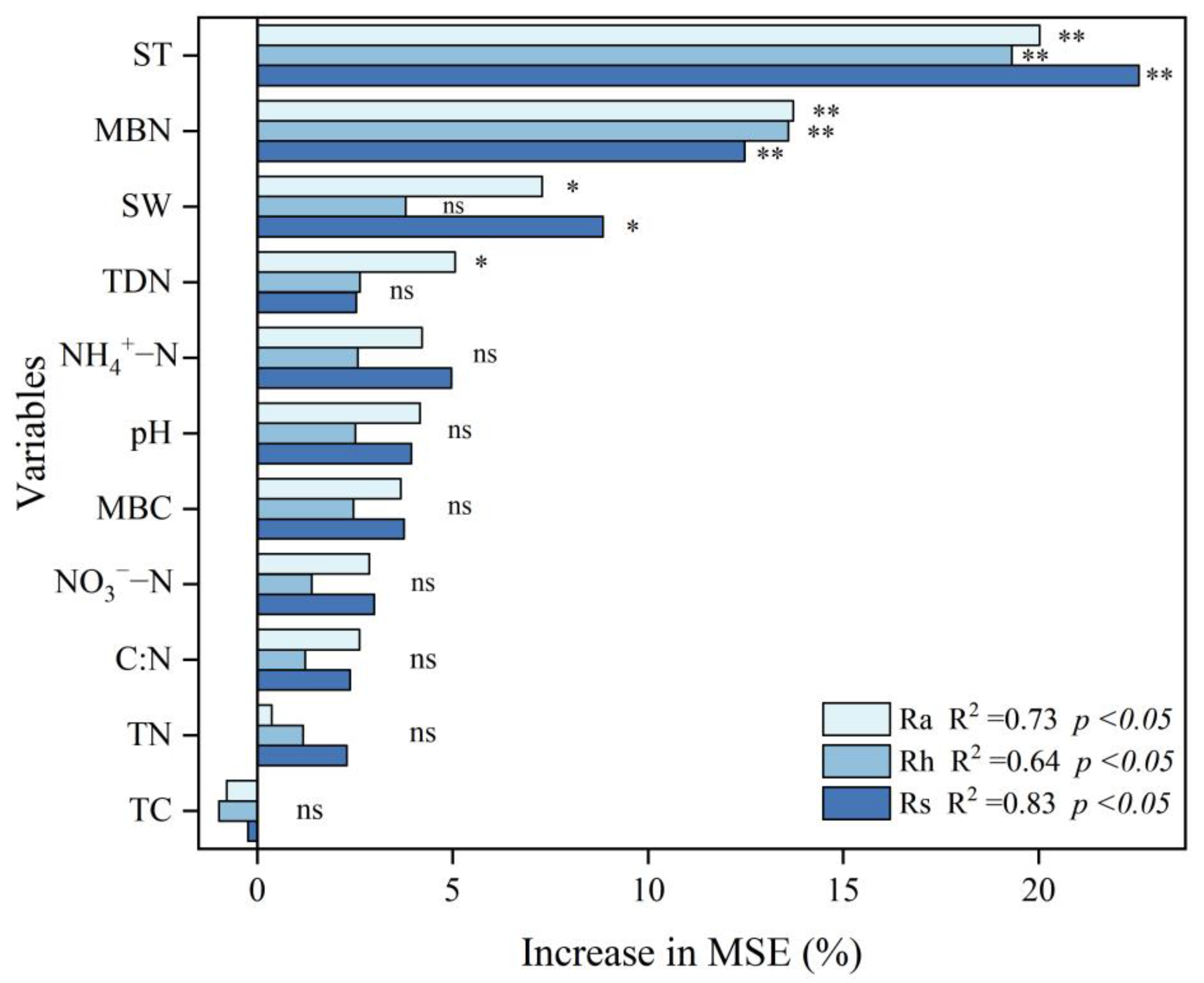
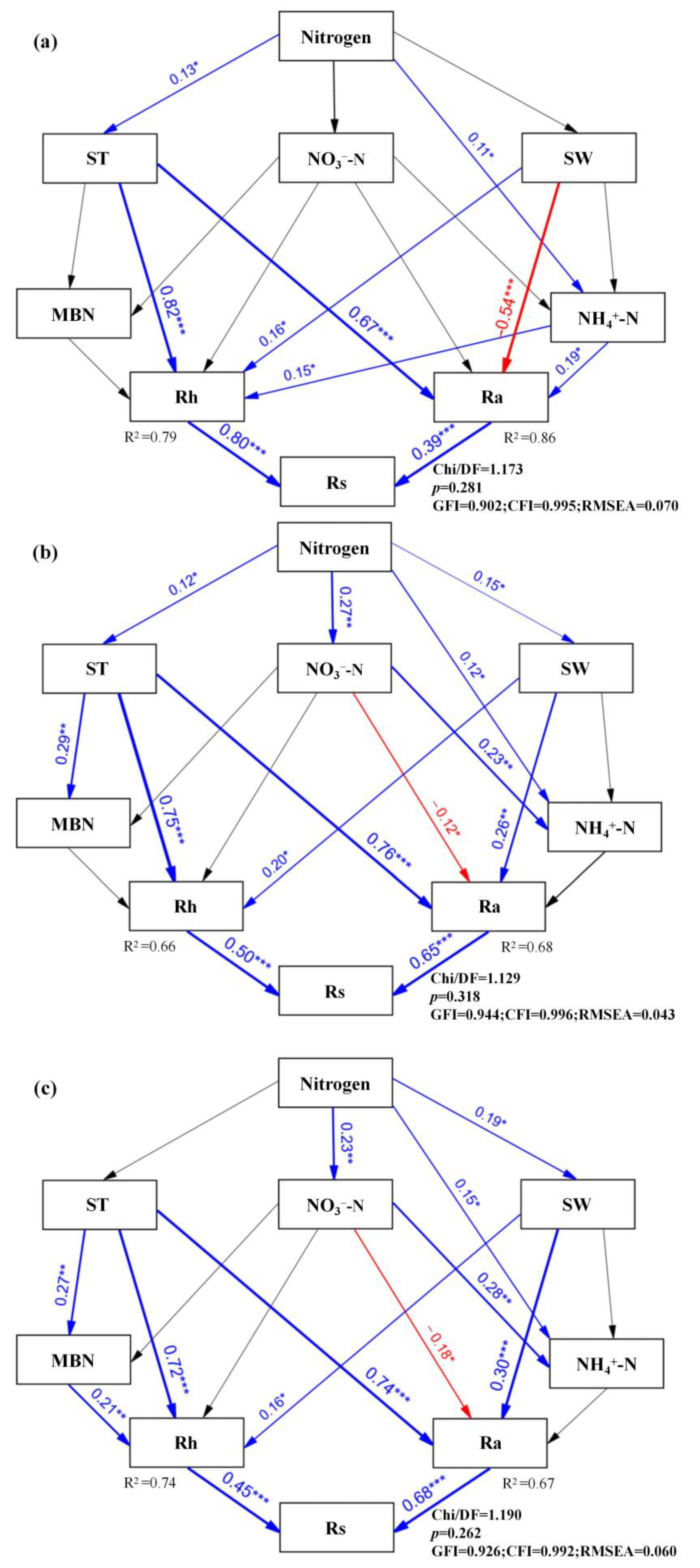
3.3. Factors Influencing Rs and Its Components
4. Discussion
4.1. Dynamics of Rs and Its Components
4.2. Factors Influencing Rs and Its Components
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the Nitrogen Cycle: Recent Trends, Questions, and Potential Solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Song, X.Z.; Chang, S.X.; Peng, C.H.; Xiao, W.F.; Zhang, J.B.; Xiang, W.H.; Li, Y.; Wang, W.F. Nitrogen depositions increase soil respiration and decrease temperature sensitivity in a Moso bamboo forest. Agric. For. Meteorol. 2019, 268, 48–54. [Google Scholar] [CrossRef]
- Luo, Q.P.; Gong, J.R.; Zhai, Z.W.; Pan, Y.; Liu, M.; Xu, S.; Wang, Y.H.; Yang, L.L.; Baoyin, T.-T. The responses of soil respiration to nitrogen addition in a temperate grassland in northern China. Sci. Total Environ. 2016, 569–570, 1466–1477. [Google Scholar] [CrossRef]
- Xiao, H.B.; Shi, Z.H.; Li, Z.W.; Wang, L.; Chen, J.; Wang, J. Responses of soil respiration and its temperature sensitivity to nitrogen addition: A meta-analysis in China. Appl. Soil Ecol. 2020, 18, 103484. [Google Scholar] [CrossRef]
- Liu, H.M.; Zhou, G.Y.; Bai, S.H.; Song, J.J.; Shang, Y.J.; He, M.; Wang, X.H.; Zheng, Z.M. Differential response of soil respiration to nitrogen and phosphorus addition in a highly phosphorus-limited subtropical forest, China. For. Ecol. Manag. 2019, 448, 499–508. [Google Scholar] [CrossRef]
- Zhou, L.Y.; Zhou, X.H.; Zhang, B.C.; Lu, M.; Luo, Y.Q.; Liu, L.L.; Li, B. Different responses of soil respiration and its components to nitrogen addition among biomes: A meta-analysis. Glob. Chang. Biol. 2014, 20, 2332–2343. [Google Scholar] [CrossRef]
- Zhou, L.Y.; Zhou, X.H.; Shao, J.J.; Nie, Y.Y.; He, Y.H.; Jiang, L.L.; Wu, Z.T.; Hosseini, B.S. Interactive effects of global change factors on soil respiration and its components: A meta-analysis. Glob. Chang. Biol. 2016, 22, 3157–3169. [Google Scholar] [CrossRef] [PubMed]
- Devaraju, N.; Bala, G.; Caldeira, K.; Nemani, R. A model based investigation of the relative importance of CO2-fertilization, climate warming, nitrogen deposition and land use change on the global terrestrial carbon uptake in the historical period. Clim. Dyn. 2016, 47, 173–190. [Google Scholar] [CrossRef]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.-A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G. Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- Liu, L.L.; Greaver, T.L. A global perspective on belowground carbon dynamics under nitrogen enrichment. Ecol. Lett. 2010, 13, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.Q.W.; Yan, W.M.; Shangguan, Z.Q. The effects of nitrogen enrichment on soil CO2 fluxes depending on temperature and soil properties. Glob. Ecol. Biogeogr. 2016, 25, 475–488. [Google Scholar] [CrossRef]
- Du, E.Z.; de Vries, W.; Han, W.X.; Liu, X.J.; Yan, Z.B.; Jiang, Y. Imbalanced phosphorus and nitrogen deposition in China’s forests. Atmos. Chem. Phys. 2016, 16, 8571–8579. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Zhao, B.; Yan, Z.C.; Yang, L.; Zhao, X.H.; Zhang, C.Y. Short-term response of soil respiration to simulated nitrogen deposition in a Pinus tabulaeformis plantation on Taiyue Mountain, China. Acta Ecol. Sin. 2018, 38, 8184–8193. [Google Scholar] [CrossRef]
- Li, C.B.; Peng, Y.F.; Nie, X.Q.; Yang, Y.H.; Yang, L.C.; Li, F.; Fang, K.; Xiao, Y.M.; Zhou, G.Y. Differential responses of heterotrophic and autotrophic respiration to nitrogen addition and precipitation changes in a Tibetan alpine steppe. Sci. Rep. 2018, 8, 16546. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Shi, Y.J.; Fang, H.Y.; Zhou, G.M.; Xu, X.J.; Zhou, Y.F.; Tao, J.X.; Ji, B.Y.; Xu, J.; Li, C.; et al. Vegetation carbon stocks driven by canopy density and forest age in subtropical forest ecosystems. Sci. Total Environ. 2018, 631–632, 619–626. [Google Scholar] [CrossRef]
- Yu, S.Q.; Chen, Y.Q.; Zhao, J.; Fu, S.L.; Li, Z.A.; Xia, H.P.; Zhou, L.X. Temperature sensitivity of total soil respiration and its heterotrophic and autotrophic components in six vegetation types of subtropical China. Sci. Total Environ. 2017, 607–608, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Li, F.M.; Pei, J.Y.; Ren, J.; Gong, Y.H.; Yuan, Z.Q.; Ke, W.B.; Zheng, Y.; Bai, X.K.; Ye, J.-S. Impacts of warming and nitrogen addition on soil autotrophic and heterotrophic respiration in a semi-arid environment. Agric. For. Meteorol. 2017, 248, 449–457. [Google Scholar] [CrossRef]
- Peng, Y.F.; Li, F.; Zhou, G.Y.; Fang, K.; Zhang, D.Y.; Li, C.B.; Yang, G.B.; Wang, G.Q.; Wang, J.; Mohammat, A.; et al. Nonlinear response of soil respiration to increasing nitrogen additions in a Tibetan alpine steppe. Environ. Res. Lett. 2017, 12, 024018. [Google Scholar] [CrossRef]
- Spohn, M.; Pötsch, E.M.; Eichorst, S.A.; Woebken, D.; Wanek, W.; Richter, A. Soil microbial carbon use efficiency and biomass turnover in a long-term fertilization experiment in a temperate grassland. Soil Biol. Biochem. 2016, 97, 168–175. [Google Scholar] [CrossRef]
- Ma, S.H.; Chen, G.P.; Tian, D.; Du, E.Z.; Xiao, W.; Jiang, L.; Zhou, Z.; Zhu, J.L.; He, H.B.; Zhu, B.; et al. Effects of seven-year nitrogen and phosphorus additions on soil microbial community structures and residues in a tropical forest in Hainan Island, China. Geoderma 2020, 361, 114034. [Google Scholar] [CrossRef]
- Peng, Y.F.; Guo, D.L.; Yang, Y.H. Global patterns of root dynamics under nitrogen enrichment. Glob. Ecol. Biogeogr. 2016, 26, 102–114. [Google Scholar] [CrossRef]
- Tu, L.H.; Dai, H.Z.; Hu, T.X.; Zhang, J.; Luo, S.H. Effect of simulated nitrogen deposition on soil respiration of poly-green hybrid bamboo forest in Yuping area, west China. Chin. J. Appl. Ecol. 2011, 22, 829–836. [Google Scholar] [CrossRef]
- Zhang, C.P.; Niu, D.C.; Hall, S.J.; Wen, H.Y.; Li, X.D.; Fu, H.; Wan, C.G.; Elser, J.J. Effects of simulated nitrogen deposition on soil respiration components and their temperature sensitivities in a semiarid grassland. Soil Biol. Biochem. 2014, 75, 113–123. [Google Scholar] [CrossRef]
- Li, X.D.; Zhang, C.Q.; Fu, H.; Guo, D.; Song, X.R.; Wan, C.Q.; Ren, J.Z. Grazing exclusion alters soil microbial respiration, root respiration and the soil carbon balance in grasslands of the Loess Plateau, northern China. Soil Sci. Plant Nutr. 2013, 59, 877–887. [Google Scholar] [CrossRef] [Green Version]
- Riggs, C.E.; Hobbie, S.E.; Bach, E.M.; Hofmockel, K.S.; Kazanski, C.E. Nitrogen addition changes grassland soil organic matter decomposition. Biogeochemistry 2015, 125, 203–219. [Google Scholar] [CrossRef]
- Zeng, W.J.; Zhang, J.Y.; Wang, W. Strong root respiration response to nitrogen and phosphorus addition in nitrogen-limited temperate forests. Sci. Total Environ. 2018, 642, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.K.; Li, Y.P.; Zhang, F.Y.; He, T.X. Short-term nitrogen fertilization decrease root and microbial respiration in a young Cunninghamia lanceolata plantation. J. Plant Ecol. 2015, 39, 1166–1175. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.K.; Zhang, W.D.; Sun, T.; Chen, L.C.; Pang, X.Y.; Wang, Y.P.; Xiao, F.M. N and P fertilization reduced soil autotrophic and heterotrophic respiration in a young Cunninghamia lanceolata forest. Agric. For. Meteorol. 2017, 232, 66–73. [Google Scholar] [CrossRef]
- Li, J.M.; Ding, W.X.; Cai, Z.C. Effects of nitrogen fertilization on soil respiration during maize growth season. Chin. J. Appl. Ecol. 2010, 21, 2025–2030. [Google Scholar] [CrossRef]
- Tomotsune, M.; Yoshitake, S.; Watanabe, S.; Hiroshi, K. Separation of root and heterotrophic respiration within soil respiration by trenching, root biomass regression, and root excising methods in a cool-temperate deciduous forest in Japan. Ecol. Res. 2013, 28, 259–269. [Google Scholar] [CrossRef]
- Fu, R.X.; Xu, X.N.; Yu, Y.C.; Zhang, Y.B.; Sun, Z.L.; Tao, X. Forest soil respiration response to increasing nitrogen deposition along an urban-rural gradient. Glob. Ecol. Conserv. 2021, 27, e01575. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Li, Y.; Ning, C.; Zheng, W.; Zhao, D.Y.; Li, Z.Q.; Yan, W.D. Litter Management as a Key Factor Relieves Soil Respiration Decay in an Urban-Adjacent Camphor Forest under a Short-Term Nitrogen Increment. Forests 2020, 11, 216. [Google Scholar] [CrossRef] [Green Version]
- Goncharova, O.; Matyshak, G.; Udovenko, M.; Semenyuk, O.; Epstein, H.; Bobrik, A. Temporal dynamics, drivers, and components of soil respiration in urban forest ecosystems. Catena 2020, 185, 104299. [Google Scholar] [CrossRef]
- Tian, P.; Liu, S.G.; Zhao, X.C.; Sun, Z.L.; Yao, X.; Niu, S.L.; Crowther, T.W.; Wang, Q.K. Past climate conditions predict the influence of nitrogen enrichment on the temperature sensitivity of soil respiration. Commun. Earth Environ. 2021, 2, 251. [Google Scholar] [CrossRef]
- Chen, Y.J.; Day, S.D.; Shrestha, R.K.; Strahm, B.D.; Wiseman, P.E. Influence of urban land development and soil rehabilitation on soil-atmosphere greenhouse gas fluxes. Geoderma 2014, 226–227, 348–353. [Google Scholar] [CrossRef]
- Du, E.Z.; Fenn, M.E.; De Vries, W.; Ok, Y.S. Atmospheric nitrogen deposition to global forests: Status, impacts and management options. Environ. Pollut. 2019, 250, 1044–1048. [Google Scholar] [CrossRef]
- Shchepeleva, A.S.; Vasenev, V.I.; Mazirov, I.M.; Vasenev, I.I.; Prokhorov, I.S.; Gosse, D.D. Changes of soil organic carbon stocks and CO2 emissions at the early stages of urban turf grasses’ development. Urban Ecosyst. 2017, 20, 309–321. [Google Scholar] [CrossRef]
- Dai, Y.Z.; Wang, H.L.; Chen, M.Y.; Wang, D.D.; Cao, X.Q.; Chu, B.Y.; Xu, X.N. Response of soil bacterial communities to nitrogen and phosphorus additions in an age-sequence of subtropical forests. iForest 2021, 14, 71–79. [Google Scholar] [CrossRef]
- Gao, Q.; Hasselquist, N.J.; Palmroth, S.; Zheng, Z.M.; You, W.H. Short-term response of soil respiration to nitrogen fertilization in a subtropical evergreen forest. Soil Biol. Biochem. 2014, 76, 297–300. [Google Scholar] [CrossRef]
- Liu, X.J.; Zhang, Y.; Han, W.X.; Tang, A.; Shen, J.L.; Cui, Z.L.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.W.; Xu, W.; Tang, A.H.; Lu, L.; Liu, X.J. Inorganic nitrogen wet deposition in eastern China Comparison of different land use-based monitoring sites in north and south regions. Chin. J. Appl. Ecol. 2016, 27, 3205–3212. [Google Scholar] [CrossRef]
- Johnson, D.; Geisinger, D.; Walker, R.; Newman, J.; Vose, J.; Elliot, K.; Ball, T. Soil pCO2, soil respiration, and root activity in CO2 fumigated and nitrogen-fertilized ponderosa pine. Plant Soil 1994, 165, 129–138. [Google Scholar] [CrossRef]
- Zhang, P.J.; Li, L.Q.; Pan, G.X.; Ren, J.C. Soil quality changes in land degradation as indicated by soil chemical, biochemical and microbiological properties in a karst area of southwest Guizhou, China. Environ. Geol. 2006, 51, 609–619. [Google Scholar] [CrossRef]
- Liu, S.; Wang, C. Spatio-temporal patterns of soil microbial biomass carbon and nitrogen in five temperate forest ecosystems. Acta. Ecol. Sin. 2010, 30, 3135–3143. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: https://www.R-project.org/ (accessed on 5 March 2021).
- Archer, E. Estimate Permutation p-Values for Importance Metrics, R Package Version 1.5.2; Robert Gentleman; MA, USA; 2013. Available online: https://cran.r-project.org/web/packages/rfPermute/index.html/ (accessed on 8 April 2022).
- Liu, H.; Zang, R.G.; Chen, H.Y.H. Effects of grazing on photosynthetic features and soil respiration of rangelands in the Tianshan Mountains of Northwest China. Sci. Rep. 2016, 6, 30087. [Google Scholar] [CrossRef] [Green Version]
- Bowden, R.D.; Davidson, E.D.; Savage, K.; Chris, A.; Paul, S. Chronic nitrogen additions reduce total soil respiration and microbial respiration intemperate forest soils at the Harvard Forest. For. Ecol. Manag. 2004, 196, 43–56. [Google Scholar] [CrossRef]
- Yan, T.; Qu, T.T.; Sun, Z.Z.; Dybzinski, R.; Chen, A.P.; Yao, X.C.; Zeng, H.; Piao, S.L. Negative effect of nitrogen addition on soil respiration dependent on stand age: Evidence from a 7-year field study of larch plantations in northern China. Agric. For. Meteorol. 2018, 262, 24–33. [Google Scholar] [CrossRef]
- Shao, R.X.; Deng, L.; Yang, Q.H.; Shangguan, Z.P. Nitrogen fertilization increase soil carbon dioxide efflux of winter wheat field: A case study in Northwest China. Soil Tillage Res. 2014, 143, 164–171. [Google Scholar] [CrossRef]
- Vicca, S.; Bahn, M.; Estiarte, M.; van Loon, E.E.; Vargas, R.; Alberti, G.; Janssens, I.A. Can current moisture responses predict soil CO2 efflux under altered precipitation regimes? A synthesis of manipulation experiments. Biogeosciences 2014, 11, 3307–3308. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.H.; Wan, S.Q. Water-and plant-mediated responses of soil respiration to topography, fire, and nitrogen fertilization in a semiarid grassland in northern China. Soil Biol. Biochem. 2008, 40, 679–687. [Google Scholar] [CrossRef]
- Chen, D.; Li, J.J.; Lan, Z.C.; Hu, S.J.; Bai, Y.F.; Niu, S.L. Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment. Funct. Ecol. 2016, 30, 658–669. [Google Scholar] [CrossRef]
- Wang, J.S.; Song, B.; Ma, F.F.; Tian, D.S.; Li, Y.; Yan, T.; Quan, Q.; Zhang, F.Y.; Li, Z.L.; Wang, B.X.; et al. Nitrogen addition reduces soil respiration but increases the relative contribution of heterotrophic component in an alpine meadow. Funct. Ecol. 2019, 33, 2239–2253. [Google Scholar] [CrossRef]
- Tu, L.H.; Hu, T.X.; Zhang, J.; Li, X.W.; Hu, H.L.; Liu, L.; Xiao, Y.L. Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem. Soil Biol. Biochem. 2013, 58, 255–264. [Google Scholar] [CrossRef]
- Baldos, A.P.; Corre, M.D.; Veldkamp, E. Response of N cycling to nutrient inputs in forest soils across a 1000–3000 m elevation gradient in the Ecuadorian Andes. Ecology 2015, 96, 749–761. [Google Scholar] [CrossRef]
- Ma, Y.C.; Zhu, B.; Sun, Z.Z.; Zhao, C.; Yang, Y.; Piao, S.L. The effects of simulated nitrogen deposition on extracellular enzyme activities of litter and soil among different-aged stands of larch. J. Plant Ecol. 2014, 7, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xu, L.; Zheng, S.; Chen, Z.; Cao, Y.Q.; Wen, X.F.; He, N.P. Temperature sensitivity of soil microbial respiration in soils with lower substrate availability is enhanced more by labile carbon input. Soil Biol. Biochem. 2021, 154, 108148. [Google Scholar] [CrossRef]
- Yang, L.; Wang, J.S.; Zhao, B.; Zhao, X.H. Effects of Long-Term Nitrogen Application on Soil Respiration and Its Components in Warm-Temperate Forest of Pinus tabulaeformis. Sci. Silvae Sin. 2021, 57, 1–11. [Google Scholar] [CrossRef]
- Song, X.Z.; Zhou, G.M.; Gu, H.H.; Qi, L.H. Management practices amplify the effects of N deposition on leaf litter decomposition of the Moso bamboo forest. Plant Soil 2015, 395, 391–400. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Baquerizo, M.; Eldridge, D.J.; Ochoa, V.; Gozalo, B.; Singh, B.K.; Maestre, F.T. Soil microbial communities drive the resistance of ecosystem multifunctionality to global change in drylands across the globe. Ecol. Lett. 2017, 20, 1295–1305. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.B.; Lv, J.H.; Li, Q.; Ying, Y.Q.; Peng, C.H.; Song, X.Z. Effects of nitrogen deposition and management practices on leaf litterfall and N and P return in a Moso bamboo forest. Biogeochemistry 2017, 134, 115–124. [Google Scholar] [CrossRef]
- Cui, J.; Wang, J.J.; Xu, J.; Xu, C.H.; Xu, X.N. Changes in soil bacterial communities in an evergreen broad-leaved forest in east China following 4 years of nitrogen addition. J. Soils Sediments 2017, 17, 2156–2164. [Google Scholar] [CrossRef]
- He, Z.; Xiong, P.F.; Jia, Z.; Zhou, J.J.; Niu, F.R.; Xu, B.C. Responses of soil respiration to rainfall depth and frequency in semiarid grassland communities. Ecohydrology 2021, 14, 2326. [Google Scholar] [CrossRef]
- Fernández-Alonso, M.; Díaz-Pinés, E.; Rubio, A. Drivers of soil respiration in response to nitrogen addition in a Mediterranean mountain forest. Biogeochemistry 2021, 155, 305–321. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.Y.; Zhang, C.C.; Wu, X.C.; Du, E.Z.; Wu, H.W.; Yang, X.F.; Wang, P.; Bai, Y.; Wu, Y.N.; et al. Responses of soil respiration to rainfall addition in a desert ecosystem: Linking physiological activities and rainfall pattern. Sci. Total Environ. 2019, 650, 3007–3016. [Google Scholar] [CrossRef]
- Peng, Q.; Dong, Y.S.; Qi, Y.C.; Xiao, S.S.; He, Y.T.; Ma, T. Effects of nitrogen fertilization on soil respiration in temperate grassland in Inner Mongolia, China. Environ. Earth Sci. 2011, 62, 1163–1171. [Google Scholar] [CrossRef]
- Boone, R.D.; Nadelhoffer, K.J.; Canary, J.D.; Kaye, J.P. Roots exert a strong influence on the temperature sensitivity of soil respiration. Nature 1998, 396, 570–572. [Google Scholar] [CrossRef]
- Xia, M.X.; Talhelm, A.F.; Pregitzer, K.S. Chronic nitrogen deposition influences the chemical dynamics of leaf litter and fine roots during decomposition. Soil Biol. Biochem. 2017, 112, 24–34. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Altitude (m) | Slope (degree) | Canopy Closure (%) | Tree Height (m) | DBH (cm) | Age (year) |
|---|---|---|---|---|---|---|
| Periurban forest | 120 | 12.26 | 0.65 | 19.7 ± 6.76 | 22.54 ± 11.97 | 120 |
| Outskirt forest | 158 | 17.51 | 0.69 | 14.9 ± 1.28 | 32.56 ± 8.08 | 135 |
| Year | Factor | Rs | Rh | Ra |
|---|---|---|---|---|
| First year | S | 117.14 *** | 69.08 *** | 48.02 *** |
| N | 2.62 * | 3.59 * | 0.36 | |
| S × N | 0.87 | 1.32 | 0.65 | |
| Second year | S | 25.31 *** | 23.57 *** | 6.79 *** |
| N | 2.43 | 13.61 *** | 7.53 *** | |
| S × N | 6.26 *** | 12.30 *** | 3.20** | |
| Third year | S | 11.90 *** | 63.44 *** | 19.72 *** |
| N | 2.88 * | 10.98 *** | 3.21 * | |
| S × N | 6.25 *** | 14.83 *** | 2.39 | |
| Whole study period | Y | 4.23 ** | 35.65 *** | 25.88 *** |
| N | 6.16 ** | 4.93 ** | 4.01 * | |
| Y × N | 1.42 | 2.98 ** | 6.00 *** |
| Variables | Year | N | Year × N |
|---|---|---|---|
| Soil physical properties | |||
| ST (°C) | 83.12 *** | 1.33 | 4.23 |
| SW (%) | 36.47 *** | 11.51 ** | 15.97 * |
| pH | 55.19 *** | 0.96 | 10.74 |
| Soil chemical properties | |||
| NH4+-N (mg/kg) | 14.15 *** | 16.00 ** | 9.38 |
| NO3−-N (mg/kg) | 34.18 *** | 26.63 *** | 10.74 |
| TDN (mg/kg) | 18.78 *** | 9.79 ** | 5.62 |
| TN (g/kg) | 60.84 *** | 4.04 | 1.46 |
| TC (g/kg) | 23.29 *** | 2.26 | 1.61 |
| C:N | 85.68 *** | 0.30 | 0.74 |
| Soil microbial biomass indexes | |||
| MBC (mg/kg) | 10.79 * | 0.08 | 2.84 |
| MBN (mg/kg) | 39.83 *** | 0.80 | 4.49 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, B.; Deng, Z.; Yu, Y.; Xu, X.; Bi, L.; Tao, X. Differential Responses of Soil Respiration and Its Components to Nitrogen Application in Urban Forests. Forests 2022, 13, 2064. https://doi.org/10.3390/f13122064
Pan B, Deng Z, Yu Y, Xu X, Bi L, Tao X. Differential Responses of Soil Respiration and Its Components to Nitrogen Application in Urban Forests. Forests. 2022; 13(12):2064. https://doi.org/10.3390/f13122064
Chicago/Turabian StylePan, Baoxu, Ziqing Deng, Yuanchun Yu, Xiaoniu Xu, Lele Bi, and Xiao Tao. 2022. "Differential Responses of Soil Respiration and Its Components to Nitrogen Application in Urban Forests" Forests 13, no. 12: 2064. https://doi.org/10.3390/f13122064