
Effect of Prescribed Burning on Tree Diversity, Biomass Stocks and Soil Organic Carbon Storage in Tropical Highland Forests

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
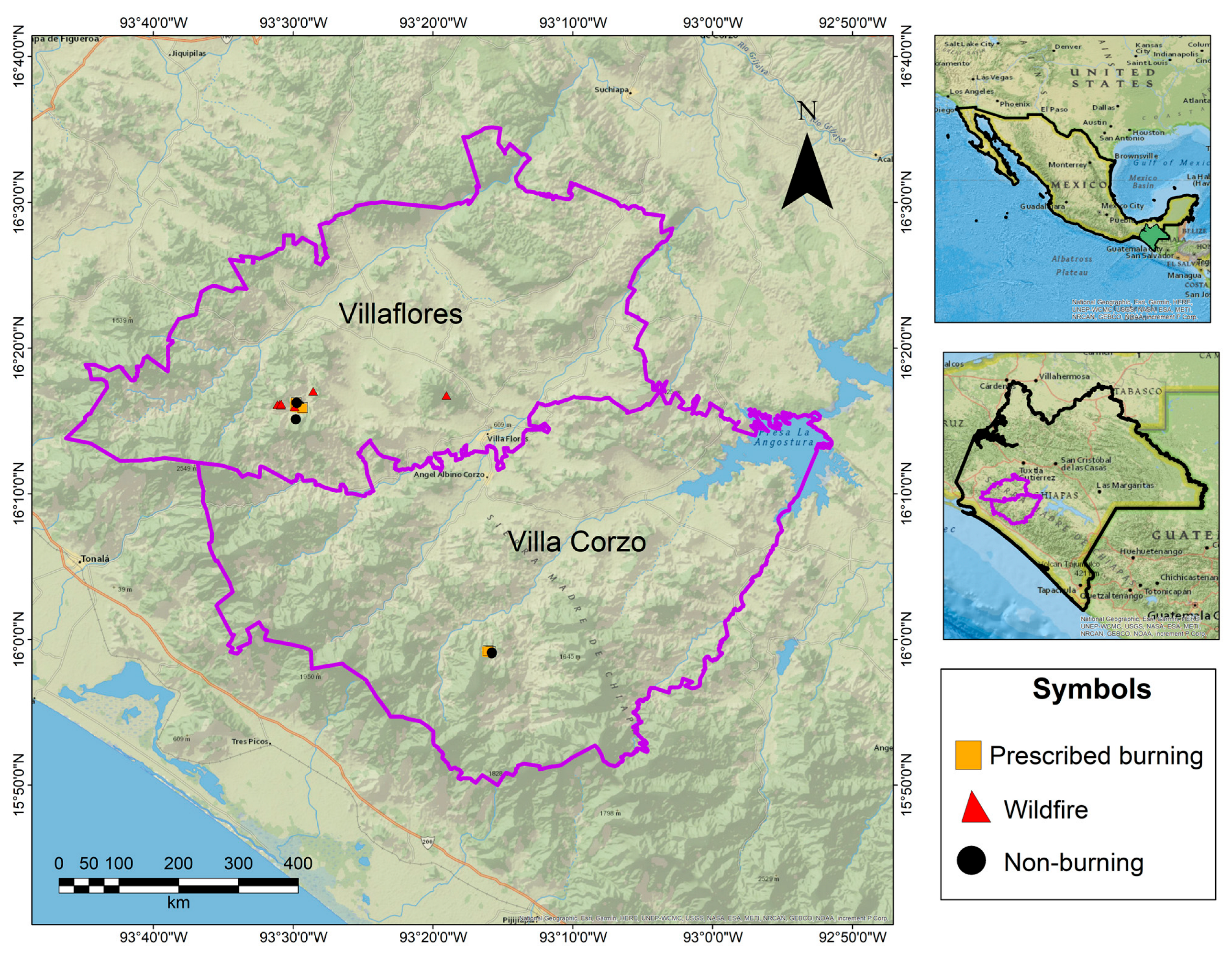
2.1. Study Sites
2.2. Sampling and Measurement of Trees
2.3. Diversity and Ecological Indices
2.4. Biomass Estimation
2.5. Soil Sampling and Analysis
2.6. Data Analysis
3. Results
3.1. Ecological Indices
3.2. Carbon Storage
3.3. Soil Properties
3.4. Relationship between Variables of Carbon Storage, Soil, and Plant Diversity
4. Discussions
4.1. Burning Effect on Ecological Indices
4.2. Burning Effects on C Storage
4.3. Burning Effects on Soil Properties
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pyles, M.V.; Magnago, L.F.S.; Maia, V.A.; Pinho, B.X.; Pitta, G.; de Gasper, A.L.; Vibrans, A.C.; Dos Santos, R.M.; van den Berg, E.; Lima, R.A. Human Impacts as the Main Driver of Tropical Forest Carbon. Sci. Adv. 2022, 8, eabl7968. [Google Scholar] [CrossRef] [PubMed]
- Aryal, D.R.; Corzo, R.R.; Cruz, A.L.; Sanabria, C.V.; Gómez, H.; Castro, F.G.H.; Ruiz, R.P.; Venegas, J.A.V.; de Coss, A.L.; Ruiz, D.M. Biomass Accumulation in Forests with High Pressure of Fuelwood Extraction in Chiapas, Mexico. Rev. Árvore 2018, 42, e420307. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Silva, S.; De Jong, B.H.; Huerta-Lwanga, E.; Mendoza-Vega, J.; Morales-Ruiz, D.E.; Aryal, D.R. Fine Root Biomass Stocks but Not the Production and Turnover Rates Vary with the Age of Tropical Successional Forests in Southern Mexico. Rhizosphere 2022, 21, 100474. [Google Scholar] [CrossRef]
- Mack, M.C.; Walker, X.J.; Johnstone, J.F.; Alexander, H.D.; Melvin, A.M.; Jean, M.; Miller, S.N. Carbon Loss from Boreal Forest Wildfires Offset by Increased Dominance of Deciduous Trees. Science 2021, 372, 280–283. [Google Scholar] [CrossRef]
- Lee, J.; Choung, Y. Key Factors Affecting the Initial Regeneration Following Forest Fires. Forests 2022, 13, 1859. [Google Scholar] [CrossRef]
- Canadell, J.G.; Meyer, C.; Cook, G.D.; Dowdy, A.; Briggs, P.R.; Knauer, J.; Pepler, A.; Haverd, V. Multi-Decadal Increase of Forest Burned Area in Australia Is Linked to Climate Change. Nat. Commun. 2021, 12, 6921. [Google Scholar] [CrossRef]
- Zheng, B.; Ciais, P.; Chevallier, F.; Chuvieco, E.; Chen, Y.; Yang, H. Increasing Forest Fire Emissions despite the Decline in Global Burned Area. Sci. Adv. 2021, 7, eabh2646. [Google Scholar] [CrossRef]
- Balch, J.K.; Abatzoglou, J.T.; Joseph, M.B.; Koontz, M.J.; Mahood, A.L.; McGlinchy, J.; Cattau, M.E.; Williams, A.P. Warming Weakens the Night-Time Barrier to Global Fire. Nature 2022, 602, 442–448. [Google Scholar] [CrossRef]
- van Wees, D.; van Der Werf, G.R.; Randerson, J.T.; Andela, N.; Chen, Y.; Morton, D.C. The Role of Fire in Global Forest Loss Dynamics. Glob. Chang. Biol. 2021, 27, 2377–2391. [Google Scholar] [CrossRef]
- Jones, M.W.; Abatzoglou, J.T.; Veraverbeke, S.; Andela, N.; Lasslop, G.; Forkel, M.; Smith, A.J.; Burton, C.; Betts, R.A.; van der Werf, G.R. Global and Regional Trends and Drivers of Fire under Climate Change. Rev. Geophys. 2022, 60, e2020RG000726. [Google Scholar] [CrossRef]
- Jones, M.W.; Santín, C.; van der Werf, G.R.; Doerr, S.H. Global Fire Emissions Buffered by the Production of Pyrogenic Carbon. Nat. Geosci. 2019, 12, 742–747. [Google Scholar] [CrossRef] [Green Version]
- Giglio, L.; Randerson, J.T.; Van Der Werf, G.R. Analysis of Daily, Monthly, and Annual Burned Area Using the Fourth-generation Global Fire Emissions Database (GFED4). J. Geophys. Res. Biogeosciences 2013, 118, 317–328. [Google Scholar] [CrossRef] [Green Version]
- FAO. Global Forest Resources Assessment 2020: Main Report; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2020. [Google Scholar]
- Liu, Z.; Ballantyne, A.P.; Cooper, L.A. Biophysical Feedback of Global Forest Fires on Surface Temperature. Nat. Commun. 2019, 10, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez Trejo, D.A.; Martínez Muñoz, P.; Lara, P.J.M. Efectos Del Fuego En El Arbolado de Un Bosque Tropical de Pino y En El de Una Selva Baja Caducifolia En Villaflores, Chiapas. Cienc. Florest. 2019, 29, 1033–1047. [Google Scholar] [CrossRef] [Green Version]
- Pompa-García, M.; Martínez-Rivas, J.A.; Valdez-Cepeda, R.D.; Aguirre-Salado, C.A.; Rodríguez-Trejo, D.A.; Miranda-Aragón, L.; Rodríguez-Flores, F.D.; Vega-Nieva, D.J. NDVI Values Suggest Immediate Responses to Fire in an Uneven-Aged Mixed Forest Stand. Forests 2022, 13, 1901. [Google Scholar] [CrossRef]
- Rodríguez-Trejo, D.A.; Martínez-Hernández, P.A.; Ortiz-Contla, H.; Chavarría-Sánchez, M.R.; Hernández-Santiago, F. The Present Status of Fire Ecology, Traditional Use of Fire, and Fire Management in Mexico and Central America. Fire Ecol. 2011, 7, 40–56. [Google Scholar] [CrossRef]
- CONAFOR. Incendios: Sistema Nacional de Información Forestal; Comision Nacional Forestal (CONAFOR): Zapopan, Jalisco, Mexico, 2020. [Google Scholar]
- Kelly, L.T.; Giljohann, K.M.; Duane, A.; Aquilué, N.; Archibald, S.; Batllori, E.; Bennett, A.F.; Buckland, S.T.; Canelles, Q.; Clarke, M.F. Fire and Biodiversity in the Anthropocene. Science 2020, 370, eabb0355. [Google Scholar] [CrossRef]
- Baptiste, A.J.; Macario, P.A.; Islebe, G.A.; Vargas-Larreta, B.; Pool, L.; Valdez-Hernández, M.; López-Martínez, J.O. Secondary Succession under Invasive Species (Pteridium Aquilinum) Conditions in a Seasonal Dry Tropical Forest in Southeastern Mexico. PeerJ 2019, 7, e6974. [Google Scholar] [CrossRef] [Green Version]
- Bataineh, M.; Portner, B.; Pelkki, M.; Ficklin, R. Prescribed Fire First-Order Effects on Oak and Maple Reproduction in Frequently Burned Upland Oak–Hickory Forests of the Arkansas Ozarks. Forests 2022, 13, 1865. [Google Scholar] [CrossRef]
- Bowman, D.M.; Williamson, G.J.; Price, O.F.; Ndalila, M.N.; Bradstock, R.A. Australian Forests, Megafires and the Risk of Dwindling Carbon Stocks. Plant Cell Environ. 2021, 44, 347–355. [Google Scholar] [CrossRef]
- Lasslop, G.; Hantson, S.; Harrison, S.P.; Bachelet, D.; Burton, C.; Forkel, M.; Forrest, M.; Li, F.; Melton, J.R.; Yue, C. Global Ecosystems and Fire: Multi-model Assessment of Fire-induced Tree-cover and Carbon Storage Reduction. Glob. Chang. Biol. 2020, 26, 5027–5041. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, A.F.; Caprio, A.C.; Georgiou, K.; Finnegan, C.; Hobbie, S.E.; Hatten, J.A.; Jackson, R.B. Low-intensity Frequent Fires in Coniferous Forests Transform Soil Organic Matter in Ways That May Offset Ecosystem Carbon Losses. Glob. Chang. Biol. 2021, 27, 3810–3823. [Google Scholar] [CrossRef] [PubMed]
- Alcañiz, M.; Outeiro, L.; Francos, M.; Úbeda, X. Effects of Prescribed Fires on Soil Properties: A Review. Sci. Total Environ. 2018, 613, 944–957. [Google Scholar] [CrossRef] [PubMed]
- Certini, G.; Moya, D.; Lucas-Borja, M.E.; Mastrolonardo, G. The Impact of Fire on Soil-Dwelling Biota: A Review. For. Ecol. Manag. 2021, 488, 118989. [Google Scholar] [CrossRef]
- Nave, L.E.; DeLyser, K.; Domke, G.M.; Holub, S.M.; Janowiak, M.K.; Kittler, B.; Ontl, T.A.; Sprague, E.; Sucre, E.B.; Walters, B.F. Disturbance and Management Effects on Forest Soil Organic Carbon Stocks in the Pacific Northwest. Ecol. Appl. 2022, 32, e2611. [Google Scholar] [CrossRef]
- Giuditta, E.; Marzaioli, R.; Esposito, A.; Ascoli, D.; Stinca, A.; Mazzoleni, S.; Rutigliano, F.A. Soil Microbial Diversity, Biomass, and Activity in Two Pine Plantations of Southern Italy Treated with Prescribed Burning. Forests 2019, 11, 19. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.V.; Aragão, L.E.; Barlow, J.; Espirito-Santo, F.; Young, P.J.; Anderson, L.O.; Berenguer, E.; Brasil, I.; Foster Brown, I.; Castro, B. Drought-Induced Amazonian Wildfires Instigate a Decadal-Scale Disruption of Forest Carbon Dynamics. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20180043. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Angerer, J.P.; Wu, X.B. The Impacts of Wildfires of Different Burn Severities on Vegetation Structure across the Western United States Rangelands. Sci. Total Environ. 2022, 845, 157214. [Google Scholar] [CrossRef]
- Rochimi, D.; Waring, K.M.; Sánchez Meador, A. Evaluating Early Post-Fire Tropical Lower Montane Forest Recovery in Indonesia. J. Trop. For. Sci. 2021, 33, 113–125. [Google Scholar] [CrossRef]
- Wright, J.; DeLaMater, D.; Simha, A.; Ury, E.; Ficken, C. Changes in Prescribed Fire Frequency Alter Ecosystem Carbon Dynamics. Ecosystems 2021, 24, 640–651. [Google Scholar] [CrossRef]
- Kolden, C.A. We’re Not Doing Enough Prescribed Fire in the Western United States to Mitigate Wildfire Risk. Fire 2019, 2, 30. [Google Scholar] [CrossRef] [Green Version]
- Greenler, S.M.; Swihart, R.K.; Saunders, M.R. Prescribed Fire Promotes Acorn Survival and Seedling Emergence from Simulated Caches. For. Ecol. Manag. 2020, 464, 118063. [Google Scholar] [CrossRef]
- Bassett, T.J.; Landis, D.A.; Brudvig, L.A. Effects of Experimental Prescribed Fire and Tree Thinning on Oak Savanna Understory Plant Communities and Ecosystem Structure. For. Ecol. Manag. 2020, 464, 118047. [Google Scholar] [CrossRef]
- Valor, T.; Casals, P.; Altieri, S.; González-Olabarria, J.R.; Piqué, M.; Battipaglia, G. Disentangling the Effects of Crown Scorch and Competition Release on the Physiological and Growth Response of Pinus Halepensis Mill. Using Δ13C and Δ18O Isotopes. For. Ecol. Manag. 2018, 424, 276–287. [Google Scholar] [CrossRef]
- Molina, J.; Ortega, M.; y Silva, F.R. Scorch Height and Volume Modeling in Prescribed Fires: Effects of Canopy Gaps in Pinus Pinaster Stands in Southern Europe. For. Ecol. Manag. 2022, 506, 119979. [Google Scholar] [CrossRef]
- Miller, R.K.; Field, C.B.; Mach, K.J. Barriers and Enablers for Prescribed Burns for Wildfire Management in California. Nat. Sustain. 2020, 3, 101–109. [Google Scholar] [CrossRef]
- Shephard, N.T.; Narine, L.; Peng, Y.; Maggard, A. Climate Smart Forestry in the Southern United States. Forests 2022, 13, 1460. [Google Scholar] [CrossRef]
- Pereira, P.; Bogunovic, I.; Zhao, W.; Barcelo, D. Short-Term Effect of Wildfires and Prescribed Fires on Ecosystem Services. Curr. Opin. Environ. Sci. Health 2021, 22, 100266. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, J.; Zhou, L.; Li, F.; Fu, S. Effects of Prescribed Burning on Carbon Accumulation in Two Paired Vegetation Sites in Subtropical China. For. Ecosyst. 2019, 6, 26. [Google Scholar] [CrossRef] [Green Version]
- Barefoot, C.R.; Willson, K.G.; Hart, J.L.; Schweitzer, C.J.; Dey, D.C. Effects of Thinning and Prescribed Fire Frequency on Ground Flora in Mixed Pinus-Hardwood Stands. For. Ecol. Manag. 2019, 432, 729–740. [Google Scholar] [CrossRef]
- Merino, A.; Jiménez, E.; Fernández, C.; Fontúrbel, M.T.; Campo, J.; Vega, J.A. Soil Organic Matter and Phosphorus Dynamics after Low Intensity Prescribed Burning in Forests and Shrubland. J. Environ. Manag. 2019, 234, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Moghli, A.; Santana, V.M.; Baeza, M.J.; Pastor, E.; Soliveres, S. Fire Recurrence and Time since Last Fire Interact to Determine the Supply of Multiple Ecosystem Services by Mediterranean Forests. Ecosystems 2022, 25, 1358–1370. [Google Scholar] [CrossRef]
- de Jesús Villar-Hernández, B.; Pérez-Elizalde, S.; Rodríguez-Trejo, D.A.; Pérez-Rodríguez, P. Análisis Espacio Temporal de La Ocurrencia de Incendios Forestales En El Estado Mexicano de Oaxaca. Rev. Mex. De Cienc. For. 2022, 13, 120–144. [Google Scholar] [CrossRef]
- Ruiz-Corzo, R.; Aryal, D.R.; Venegas-Sandoval, A.; Jerez-Ramírez, D.O.; Fernández-Zúñiga, K.S.; del Carmen Lopez-Cruz, S.; López-Herández, J.C.; Peña-Alvarez, B.; Velázquez-Sanabria, C.A. Dinámica Temporal de Combustibles Forestales y Efecto Del Incendio En Cerro Nambiyugua, Chiapas, México. Ecosistemas Y Recur. Agropecu. 2022, 9, e3253. [Google Scholar] [CrossRef]
- Aryal, D.R.; Morales-Ruiz, D.E.; López-Cruz, S.; Tondopó-Marroquín, C.N.; Lara-Nucamendi, A.; Jiménez-Trujillo, J.A.; Pérez-Sánchez, E.; Betanzos-Simon, J.E.; Casasola-Coto, F.; Martínez-Salinas, A. Silvopastoral Systems and Remnant Forests Enhance Carbon Storage in Livestock-Dominated Landscapes in Mexico. Sci. Rep. 2022, 12, 16769. [Google Scholar] [CrossRef]
- McLauchlan, K.K.; Higuera, P.E.; Miesel, J.; Rogers, B.M.; Schweitzer, J.; Shuman, J.K.; Tepley, A.J.; Varner, J.M.; Veblen, T.T.; Adalsteinsson, S.A. Fire as a Fundamental Ecological Process: Research Advances and Frontiers. J. Ecol. 2020, 108, 2047–2069. [Google Scholar] [CrossRef]
- Semarnat/Sagarpa Norma Oficial Mexicana Nom-015 Especificaciones Técnicas de Los Métodos de Uso Del Fuego En Los Terrenos Forestales y En Los Terrenos de Uso Agropecuario. 2009, pp. 1–77. Available online: https://www.gob.mx/profepa/documentos/norma-oficial-mexicana-nom-015-semarnat-sagarpa-2007 (accessed on 8 November 2022).
- Cottam, G.; Curtis, J.T. The Use of Distance Measures in Phytosociological Sampling. Ecology 1956, 37, 451–460. [Google Scholar] [CrossRef]
- Sorensen, T.A. A Method of Establishing Groups of Equal Amplitude in Plant Sociology Based on Similarity of Species Content and Its Application to Analyses of the Vegetation on Danish Commons. Biol. Skar. 1948, 5, 1–34. [Google Scholar]
- Vargas-Larreta, B.; López-Sánchez, C.A.; Corral-Rivas, J.J.; López-Martínez, J.O.; Aguirre-Calderón, C.G.; Álvarez-González, J.G. Allometric Equations for Estimating Biomass and Carbon Stocks in the Temperate Forests of North-Western Mexico. Forests 2017, 8, 269. [Google Scholar] [CrossRef] [Green Version]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C. Improved Allometric Models to Estimate the Aboveground Biomass of Tropical Trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Cairns, M.A.; Brown, S.; Helmer, E.H.; Baumgardner, G.A. Root Biomass Allocation in the World’s Upland Forests. Oecologia 1997, 111, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Van Wagner, C. Practical Aspects of the Line Intersect Method; Canadian Forestry Service: Victoria, BC, Canada, 1982; Volume 12, ISBN ISSN 0706-1854. [Google Scholar]
- Aryal, D.R.; De Jong, B.H.; Gaona, S.O.; Vega, J.M.; Olguín, L.E.; Cruz, S.L. Fine Wood Decomposition Rates Decline with the Sge of Tropical Successional Forests in Southern Mexico: Implications to Ecosystem Carbon Storage. Ecosystems 2022, 25, 661–677. [Google Scholar] [CrossRef]
- Heanes, D. Determination of Total Organic-C in Soils by an Improved Chromic Acid Digestion and Spectrophotometric Procedure. Commun. Soil Sci. Plant Anal. 1984, 15, 1191–1213. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I. An Examination of the Degtjareff Method for Determining Soil Organic Matter and a Proposed Modification of the Chromic Acid Titration Method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Morales Ruiz, D.E.; Aryal, D.R.; Pinto Ruiz, R.; Guevara Hernández, F.; Casanova Lugo, F.; Villanueva Lopez, G. Carbon Contents and Fine Root Production in Tropical Silvopastoral Systems. Land Degrad. Dev. 2021, 32, 738–756. [Google Scholar] [CrossRef]
- IBM Corporation IBM SPSS Statistics 25. 2017. Available online: https://www.ibm.com/mx-es/products/spss-statistics (accessed on 23 June 2021).
- Durigan, G.; Pilon, N.A.; Abreu, R.C.; Hoffmann, W.A.; Martins, M.; Fiorillo, B.F.; Antunes, A.Z.; Carmignotto, A.P.; Maravalhas, J.B.; Vieira, J. No Net Loss of Species Diversity after Prescribed Fires in the Brazilian Savanna. Front. For. Glob. Chang. 2020, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, M.J.; North, M.P.; Zald, H.S.; Hurteau, M.D. Changing Climate Reallocates the Carbon Debt of Frequent-fire Forests. Glob. Change Biol. 2020, 26, 6180–6189. [Google Scholar] [CrossRef]
- Brockway, D.G.; Lewis, C.E. Long-Term Effects of Dormant-Season Prescribed Fire on Plant Community Diversity, Structure and Productivity in a Longleaf Pine Wiregrass Ecosystem. For. Ecol. Manag. 1997, 96, 167–183. [Google Scholar] [CrossRef]
- Santos, R.O.D.; Souza, A.P.D.; Soares, R.N.; Rabelo, F.G.; Costa, S.V.D. Effect of Fire under the Soils on the Organization of Communities of Three Remnants of Amazonian Savannas. Acta Bot. Bras. 2020, 34, 505–521. [Google Scholar] [CrossRef]
- Aryal, D.R. Grazing Intensity in Grassland Ecosystems: Implications for Carbon Storage and Functional Properties. CABI Rev. 2022. [Google Scholar] [CrossRef]
- Morici, K.E.; Bailey, J.D. Long-Term Effects of Fuel Reduction Treatments on Surface Fuel Loading in the Blue Mountains of Oregon. Forests 2021, 12, 1306. [Google Scholar] [CrossRef]
- Girona-García, A.; Ortiz-Perpiñá, O.; Badía-Villas, D.; Martí-Dalmau, C. Effects of Prescribed Burning on Soil Organic C, Aggregate Stability and Water Repellency in a Subalpine Shrubland: Variations among Sieve Fractions and Depths. Catena 2018, 166, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Volkova, L.; Roxburgh, S.H.; Weston, C.J. Effects of Prescribed Fire Frequency on Wildfire Emissions and Carbon Sequestration in a Fire Adapted Ecosystem Using a Comprehensive Carbon Model. J. Environ. Manag. 2021, 290, 112673. [Google Scholar] [CrossRef] [PubMed]
- Fontúrbel, T.; Carrera, N.; Vega, J.A.; Fernández, C. The Effect of Repeated Prescribed Burning on Soil Properties: A Review. Forests 2021, 12, 767. [Google Scholar] [CrossRef]
- James, J.A.; Kern, C.C.; Miesel, J.R. Legacy Effects of Prescribed Fire Season and Frequency on Soil Properties in a Pinus Resinosa Forest in Northern Minnesota. For. Ecol. Manag. 2018, 415, 47–57. [Google Scholar] [CrossRef]
- Nichols, L.; Shinneman, D.J.; McIlroy, S.K.; de Graaff, M.-A. Fire Frequency Impacts Soil Properties and Processes in Sagebrush Steppe Ecosystems of the Columbia Basin. Appl. Soil Ecol. 2021, 165, 103967. [Google Scholar] [CrossRef]
- Hahn, G.E.; Coates, T.A.; Aust, W.M. Soil Chemistry Following Single-Entry, Dormant Season Prescribed Fires in the Ridge and Valley Province of Virginia, USA. Commun. Soil Sci. Plant Anal. 2021, 52, 2065–2073. [Google Scholar] [CrossRef]
- Pellegrini, A.F.; Harden, J.; Georgiou, K.; Hemes, K.S.; Malhotra, A.; Nolan, C.J.; Jackson, R.B. Fire Effects on the Persistence of Soil Organic Matter and Long-Term Carbon Storage. Nat. Geosci. 2022, 15, 5–13. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diversity and Structure | DBH Category | Mean (95% Confidence Interval) | ||
|---|---|---|---|---|
| Prescribed Burning | Non-Burning | Wildfire | ||
| Shannon (H) index | All trees | 1.26 (0.97–1.55) a | 1.14 (0.81–1.47) a | 0.36 (0.12–0.59) b |
| Simpson (D) index | All tress | 0.37 (0.22–0.52) a | 0.42 (0.28–0.56) a | 0.67 (0.37–0.97) a |
| Tree height (m) | <7.5 cm | 1.73 (0.73–4.10) | 5.95 (3.85–9.19) | 3.59 (1.51–8.51) |
| >7.5 cm | 13.50 (11.72–15.56) | 12.99 (11.25–15.00) | 11.76 (10.19–13.56) | |
| All trees | 13.00 (11.79–14.33) | 11.42 (9.63–13.56) | 10.73 (8.76–13.14) | |
| Crown diameter (m) | <7.5 cm | 1.76 (0.72–4.29) | 5.69 (3.31–9.81) | 1.58 (0.71–3.51) |
| >7.5 cm | 53.25 (25.31–112.05) | 37.89 (15.59–92.07) | 24.30 (11.66–50.62) | |
| All trees | 50.80 (23.13–111.59) | 31.52 (13.45–73.88) | 20.90 (9.83–44.42) | |
| Basal area (m2 ha−1) | <7.5 cm | 0.81 (0.57–1.16) | 0.21 (0.10–0.43) | 0.28 (0.10–0.76) |
| >7.5 cm | 12.22 (6.73–22.17) | 10.88 (7.68–15.43) | 9.77 (7.15–13.35) | |
| All trees | 12.34 (6.78–22.45) | 11.12 (7.81–15.84) | 9.97 (7.41–13.41) | |
| Sorensen’s (Jaccard’s) Similarity Indices between Burning Treatments | |||
|---|---|---|---|
| Prescribed Burning | Non-Burning | Wildfire | |
| Prescribed burning | 78 (64) | 59 (42) | |
| Non-burning | 55 (38) | ||
| Wildfire | |||
| Carbon Pools | Carbon Stocks (Mg ha−1) Mean (95% Confidence Interval) | ||
|---|---|---|---|
| Prescribed Burning | Non-Burning | Wildfire | |
| AGB (Mg C ha−1) | 41.96 (28.85–55.06) | 35.23 (22.12–48.33) | 26.14 (13.03–39.24) |
| BGB (Mg C ha−1) | 10.08 (7.10–13.05) | 8.60 (5.62–11.58) | 6.53 (3.56–9.51) |
| Total biomass (Mg C ha−1) | 52.04 (35.96–68.12) | 43.83 (27.75–59.91) | 32.67 (16.59–48.75) |
| Deadwood (Mg C ha−1) | 2.28 (1.06–3.51) | 1.20 (0.00–2.43) | 0.59 (0.00–1.82) |
| Litter (Mg C ha−1) | 3.59 (2.33–4.86) | 4.65 (3.38–5.92) | 2.68 (1.42–3.95) |
| SOC (Mg C ha−1) | 183.9 (166.1–201.7) | 167.9 (150.1–185.7) | 144.3 (126.5–162.1) |
| Variables | Component 1 (Soil and Diversity) | Component 2 (Carbon Store) | Component 3 (Litter Accumulation) | Component 4 (Deadwood) |
|---|---|---|---|---|
| SOC content % | 0.13 | 0.76 | 0.03 | 0.11 |
| Soil pH | 0.87 | 0.02 | −0.16 | 0.06 |
| Soil_moisture | 0.66 | −0.39 | 0.20 | −0.39 |
| Soil bulk density | −0.37 | 0.72 | 0.18 | −0.22 |
| Tree biomass | 0.27 | 0.71 | 0.07 | 0.19 |
| Litter mass | 0.25 | 0.13 | −0.77 | −0.13 |
| Deadwood mass | 0.13 | −0.12 | 0.01 | −0.96 |
| Fine root biomass | −0.15 | 0.56 | −0.45 | 0.06 |
| Tree diversity | 0.86 | 0.21 | −0.04 | −0.18 |
| Elevation | −0.41 | −0.43 | 0.30 | −0.03 |
| Slope of the terrain | −0.04 | −0.28 | −0.84 | 0.16 |
| Variance explained | 22.15% | 21.83% | 15.47% | 11.47% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Cruz, S.d.C.; Aryal, D.R.; Velázquez-Sanabria, C.A.; Guevara-Hernández, F.; Venegas-Sandoval, A.; Casanova-Lugo, F.; La O-Arias, M.A.; Venegas-Venegas, J.A.; Reyes-Sosa, M.B.; Pinto-Ruiz, R.; et al. Effect of Prescribed Burning on Tree Diversity, Biomass Stocks and Soil Organic Carbon Storage in Tropical Highland Forests. Forests 2022, 13, 2164. https://doi.org/10.3390/f13122164
López-Cruz SdC, Aryal DR, Velázquez-Sanabria CA, Guevara-Hernández F, Venegas-Sandoval A, Casanova-Lugo F, La O-Arias MA, Venegas-Venegas JA, Reyes-Sosa MB, Pinto-Ruiz R, et al. Effect of Prescribed Burning on Tree Diversity, Biomass Stocks and Soil Organic Carbon Storage in Tropical Highland Forests. Forests. 2022; 13(12):2164. https://doi.org/10.3390/f13122164
Chicago/Turabian StyleLópez-Cruz, Susana del Carmen, Deb Raj Aryal, Carlos Alberto Velázquez-Sanabria, Francisco Guevara-Hernández, Andrea Venegas-Sandoval, Fernando Casanova-Lugo, Manuel Alejandro La O-Arias, José Apolonio Venegas-Venegas, Mariela Beatriz Reyes-Sosa, René Pinto-Ruiz, and et al. 2022. "Effect of Prescribed Burning on Tree Diversity, Biomass Stocks and Soil Organic Carbon Storage in Tropical Highland Forests" Forests 13, no. 12: 2164. https://doi.org/10.3390/f13122164
APA StyleLópez-Cruz, S. d. C., Aryal, D. R., Velázquez-Sanabria, C. A., Guevara-Hernández, F., Venegas-Sandoval, A., Casanova-Lugo, F., La O-Arias, M. A., Venegas-Venegas, J. A., Reyes-Sosa, M. B., Pinto-Ruiz, R., Hernández-López, A., Medina-Jonapá, F. J., Ramírez-Diaz, R., López-Cruz, A., & Alcudia-Aguilar, A. (2022). Effect of Prescribed Burning on Tree Diversity, Biomass Stocks and Soil Organic Carbon Storage in Tropical Highland Forests. Forests, 13(12), 2164. https://doi.org/10.3390/f13122164