
Identification and Tissue-Specific Expression Analysis of CYP720B Subfamily Genes in Slash Pine and Loblolly Pine



Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of CYP720B in Slash Pine and Loblolly Pine
2.2. Sequence Analysis of CYP720Bs
2.3. Plant Materials and Growth Conditions
2.4. RNA Isolation and Gene Expression Analyses
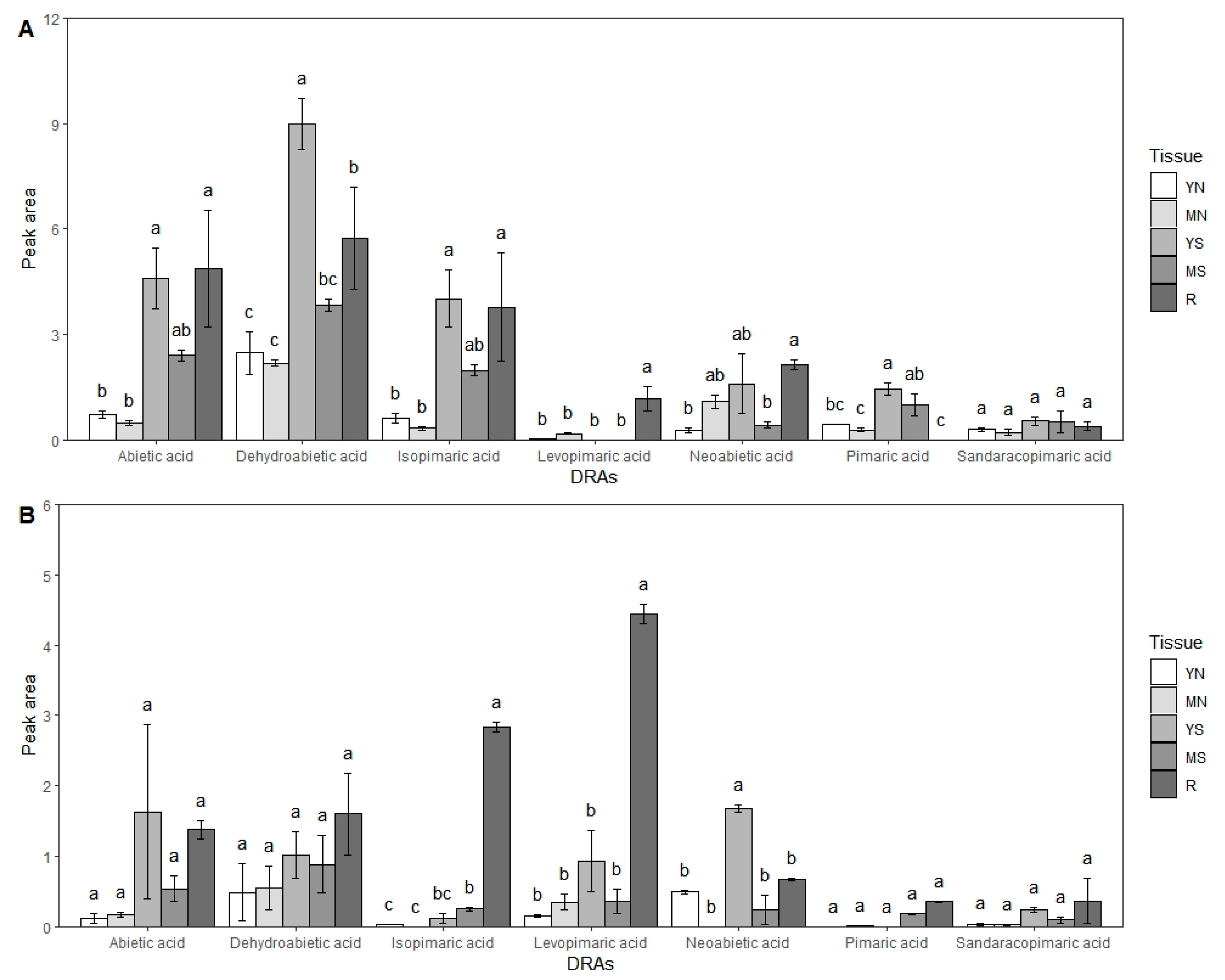
2.5. Extraction of DRAs
3. Results
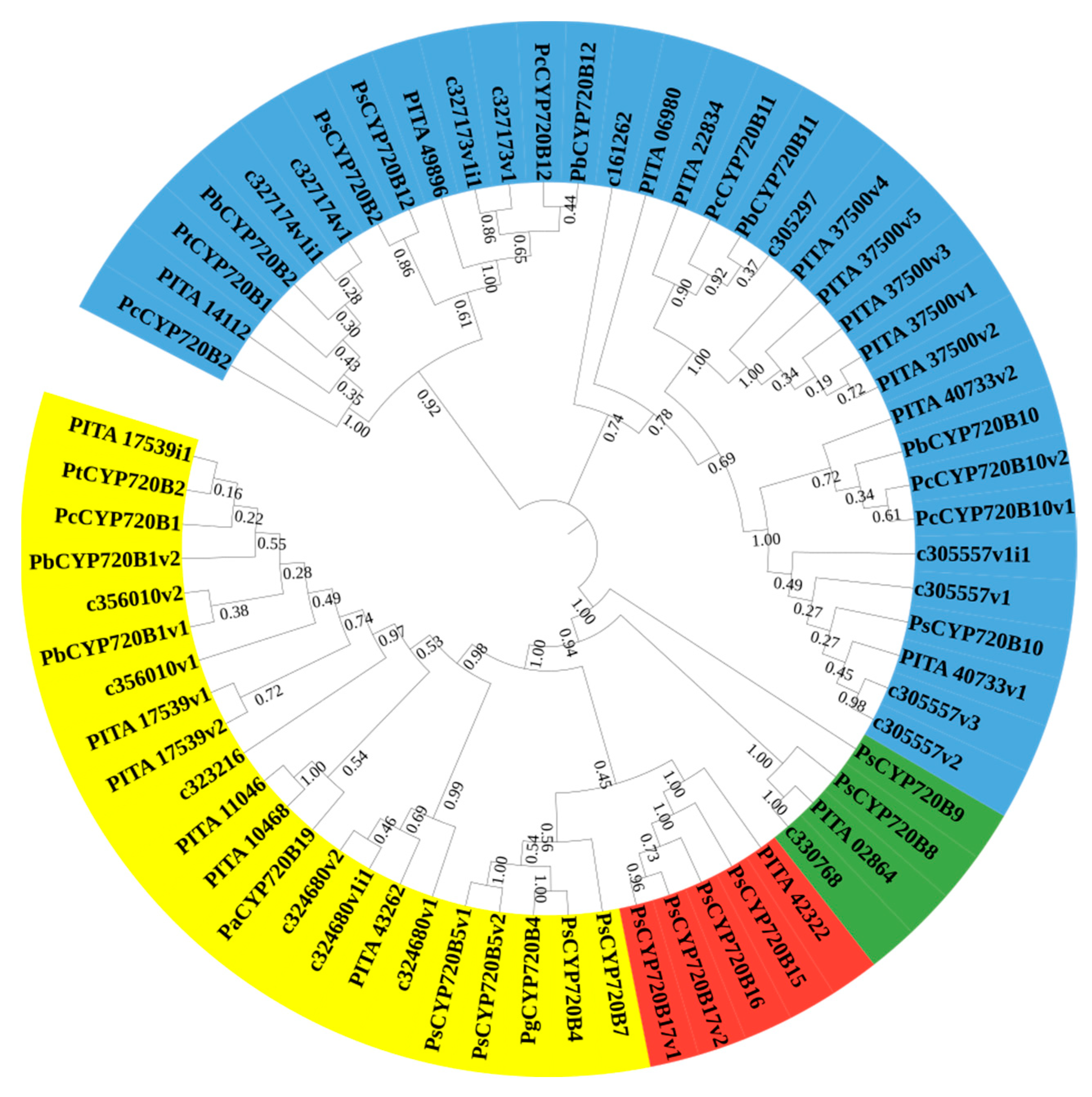
3.1. Identification and Phylogenetic Analysis of CYP720B in Slash Pine and Loblolly Pine
3.2. Multiple Sequence Alignment, Phylogenetic Analysis, and Classification of CYP720B
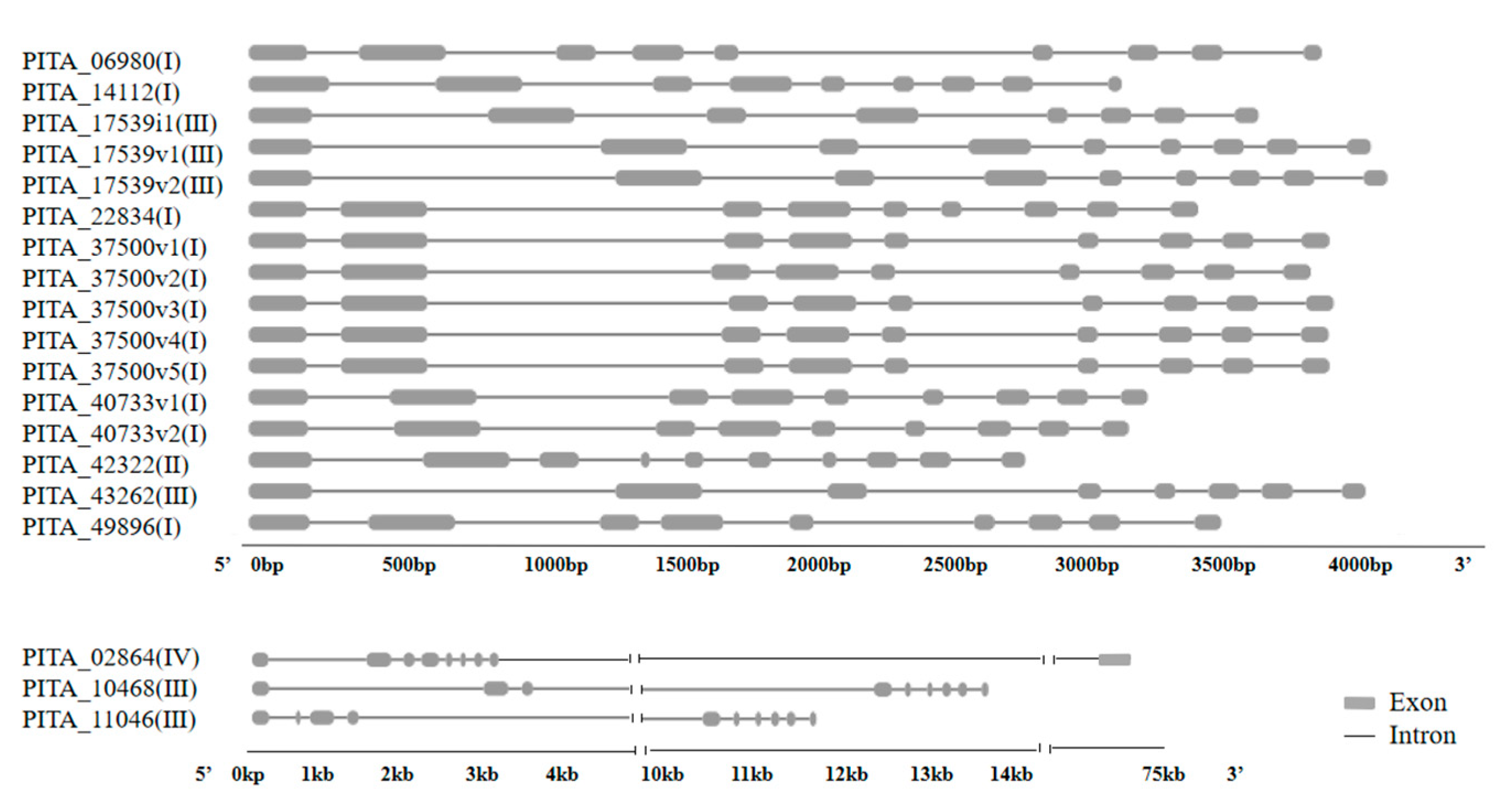
3.3. Gene Structure and Motif Composition of CYP720B
3.4. Upstream Cis-Acting Elements of Loblolly Pine CYP720B
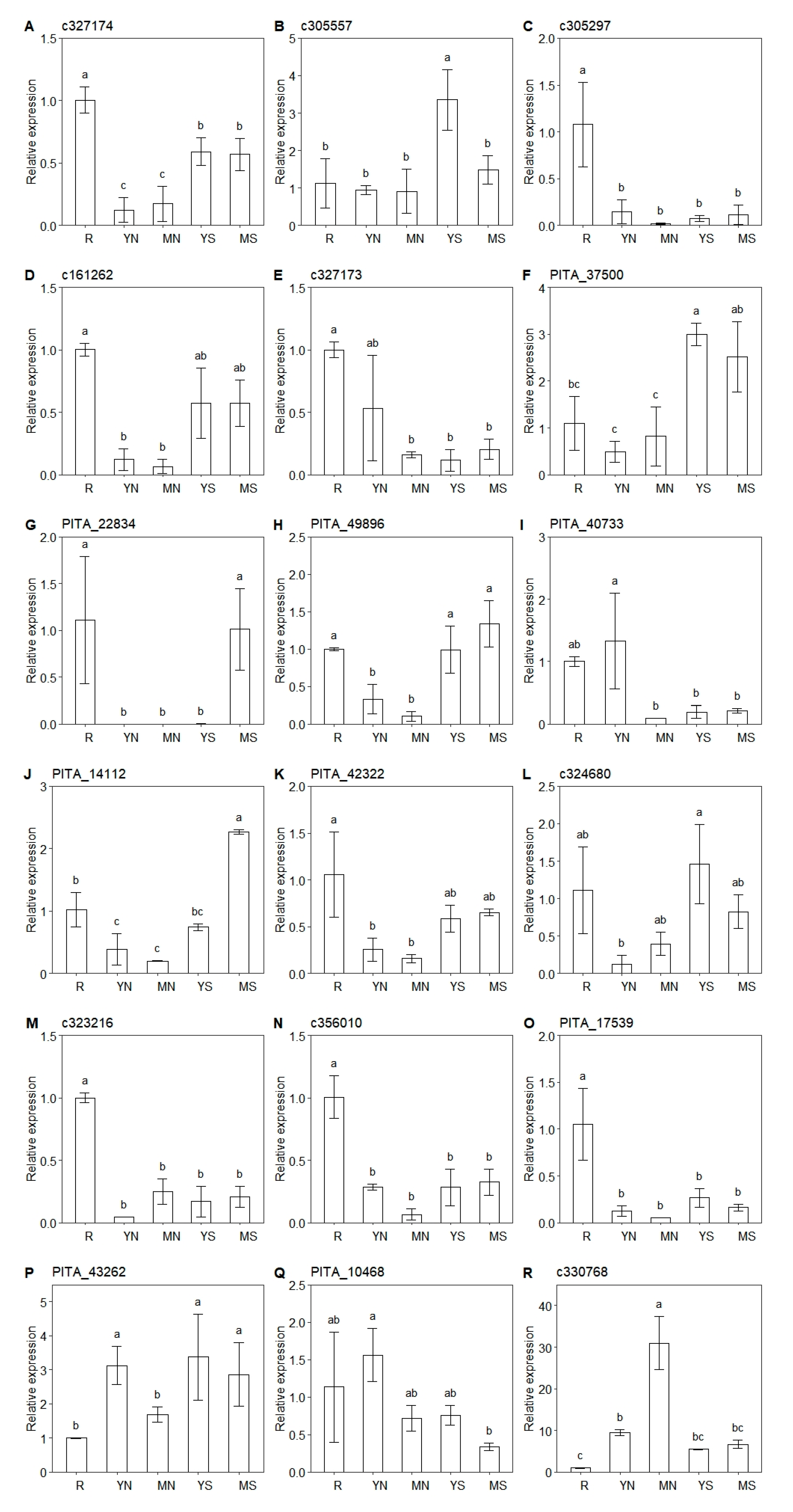
3.5. Revealing the Difference in the Expression of CYP720Bs in Different Tissues
4. Discussion
4.1. Characterization of CYP720B
4.2. Function of CYP720B
4.3. Expression Specificity of CYP720B
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Homologous Protein | Accession Number | Species | Homologous Protein | Accession Number | Species |
|---|---|---|---|---|---|
| PsCYP720B12 | HM245397 | Picea sitchensis | PbCYP720B1v1 | KJ845665 | Pinus banksiana |
| PsCYP720B15 | HM245398 | Picea sitchensis | PbCYP720B1v2 | KJ845666 | Pinus banksiana |
| PsCYP720B16 | HM245399 | Picea sitchensis | PbCYP720B2 | KJ845667 | Pinus banksiana |
| PsCYP720B17v1 | HM245400 | Picea sitchensis | PbCYP720B10 | KJ845668 | Pinus banksiana |
| PsCYP720B17v2 | HM245401 | Picea sitchensis | PbCYP720B11 | KJ845669 | Pinus banksiana |
| PsCYP720B2 | HM245402 | Picea sitchensis | PbCYP720B12 | KJ845670 | Pinus banksiana |
| PsCYP720B4 | HM245403 | Picea sitchensis | PcCYP720B1 | KJ845671 | Pinus contorta |
| PsCYP720B5v1 | HM245404 | Picea sitchensis | PcCYP720B2 | KJ845672 | Pinus contorta |
| PsCYP720B5v2 | HM245405 | Picea sitchensis | PcCYP720B10v1 | KJ845673 | Pinus contorta |
| PsCYP720B7 | HM245406 | Picea sitchensis | PcCYP720B10v2 | KJ845674 | Pinus contorta |
| PsCYP720B8 | HM245407 | Picea sitchensis | PcCYP720B11 | KJ845675 | Pinus contorta |
| PsCYP720B10 | HM245408 | Picea sitchensis | PcCYP720B12 | KJ845676 | Pinus contorta |
| PsCYP720B9 | HM245410 | Picea sitchensis | PaCYP720B19 | KJ624415 | Pinus armandi |
| PtCYP720B2 | Q50EK5 | Pinus taeda | PgCYP720B4 | FJ609175 | Pinus glauca |
| PtCYP720B1 | Q50EK6 | Pinus taeda |
| Gene_ID | F′(to 3′) | R′(to 5′) |
|---|---|---|
| PITA_02864 | ATCCAGAGTTGAGTGCACCA | AGAACGAGGGAAGGCTCTTT |
| PITA_06980 | GGCAGGCTGTTTCAATCCAA | AACGTTCTGTATGCCCTCCA |
| PITA_10468 | ATTATCGCTCCATGACCAGC | AGAACCTCCCCTCGTTTTGT |
| PITA_11046 | TTATTCGGAAGCCCAGCAGT | AGCCTCTCAAATCCCAGCAA |
| PITA_40733 | ATGGCTGGTGTTCTTCGTCT | TTGGCGTGGTTGAGAAATGG |
| PITA_42322 | AGCCTCTCAAACCTCAGCAA | AGCAGATCCCCAGTTCAACA |
| PITA_49896 | ACAAACATGTCCTGCAGCAC | AGCCTCTCGAACCTCAACAA |
| PITA_37500 | TTCTGCGCTCACATTTGACC | ATGAAGAAAGAGGGCCAGCT |
| PITA_14112 | ACCTCTCGAACCTCAGCAAA | TTGTGTCCGTGGATCCAGAA |
| PITA_22834 | TGTCCTCCATGAAGTGCACA | GGCAGGCTGTTTCAATCCAA |
| PITA_17539 | TGTCGTCGATGAATTGCCTG | TTTACAGGTGGTGGAATGCC |
| PITA_43262 | ACCGCTATTCCATGGAGCTT | TCAGCAGATCCCCAGTTCAA |
| c356010 | TTACAGGTGGTGGAATGCC | TGTCGTCGATGAATTGCCTG |
| c323216 | CTAATCGAGAGGTACATCTGCC | TTGAACTGGGGATCTGCTGA |
| c327174 | AGTGCTGGGCTTCTTATTGC | CGTGGTTGAGCAGTGGAATT |
| c161262 | TTCGTCTGTTTCGTTCTGGC | CTTGAATGAATCGGCGTGGT |
| c327173 | AAGCATCCACAAGTTGTCCG | GTCGTGCTCAGCCTTCAATT |
| c305297 | CCATTTCTCAACCACGCCAA | ATTGAACCAGCTTGCCTTCG |
| c305557 | AAACACATCGTCTGGCCAAC | AACGCCATGGGTCAAACTTG |
| c330768 | AGAACGAGGGAAGGCTCTTT | ATCCAGAGTTGAGTGCACCA |
| c324680 | AAGCTCCATGGAATAGCGGT | GAGCAGTTGTTTGGCCATCA |
References
- Keeling, C.I.; Bohlmann, J. Diterpene Resin Acids in Conifers. Phytochemistry 2006, 67, 2415–2423. [Google Scholar] [CrossRef] [PubMed]
- Stepp, J.R. Plant Resins: Chemistry, Evolution, Ecology, Ethnobotany. Econ. Bot. 2003, 57, 419–420. [Google Scholar] [CrossRef]
- Keeling, C.I.; Weisshaar, S.; Lin, R.P.C.; Bohlmann, J. Functional Plasticity of Paralogous Diterpene Synthases Involved in Conifer Defense. Proc. Natl. Acad. Sci. USA 2008, 105, 1085–1090. [Google Scholar] [CrossRef] [Green Version]
- Zerbe, P.; Chiang, A.; Yuen, M.; Hamberger, B.; Hamberger, B.; Draper, J.A.; Britton, R.; Bohlmann, J. Bifunctional Cis-abienol Synthase from Abies Balsamea Discovered by Transcriptome Sequencing and Its Implications for Diterpenoid Fragrance Production. J. Biol. Chem. 2012, 287, 12121–12131. [Google Scholar] [CrossRef] [Green Version]
- Hall, D.E.; Zerbe, P.; Jancsik, S.; Quesada, A.L.; Dullat, H.; Madilao, L.L.; Yuen, M.; Bohlmann, J. Evolution of Conifer Diterpene Synthases: Diterpene Resin Acid Biosynthesis in Lodgepole Pine and Jack Pine Involves Monofunctional and Bifunctional Diterpene Synthases. Plant Physiol. 2013, 161, 600–616. [Google Scholar] [CrossRef] [Green Version]
- Hartmut, K.L. The 1-deoxy-D-xylulose-5-phosphate Pathway of Isoprenoid Biosynthesis in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 47–65. [Google Scholar] [CrossRef]
- Geisler, K.; Jensen, N.B.; Yuen, M.M.S.; Madilao, L.; Bohlmann, J. Modularity of Conifer Diterpene Resin Acid Biosynthesis: P450 Enzymes of Different CYP720B Clades Use Alternative Substrates and Converge on the Same Products. Plant Physiol. 2016, 171, 152–164. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, C.; Lu, W. Heterologous Production of Levopimaric Acid in Saccharomyces Cerevisiae. Microb. Cell Fact 2018, 17, 1–10. [Google Scholar] [CrossRef]
- Celedon, J.M.; Bohlmann, J. Oleoresin Defenses in Conifers: Chemical Diversity, Terpene Synthases and Limitations of Oleoresin Defense under Climate Change. New Phytol. 2019, 224, 1444–1463. [Google Scholar] [CrossRef] [Green Version]
- Aparajita, B.; Björn, H. P450s Controlling Metabolic Bifurcations in Plant Terpene Specialized Metabolism. Phytochem. Rev. 2018, 17, 80–111. [Google Scholar] [CrossRef] [Green Version]
- Semiz, A.; Sen, A. Cloning and Expression of a CYP720B Orthologue Involved in the Biosynthesis of Diterpene Resin Acids in Pinus brutia. Mol. Biol. Rep. 2015, 42, 737–744. [Google Scholar] [CrossRef]
- Warren, R.L.; Keeling, C.I.; Yuen, M.M.; Raymond, A.; Taylor, G.A.; Vandervalk, B.P.; Mohamadi, H.; Paulino, D.; Chiu, R.; Jackman, S.D.; et al. Improved White spruce (Picea glauca) Genome Assemblies and Annotation of Large Gene Families of Conifer Terpenoid and Phenolic Defense Metabolism. Plant J. 2015, 83, 189–212. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Z.Y.; Lu, Q.; Zhang, X.Y. A Review of Induced Defenses in Conifers. J. Anhui Agri. Sci. 2014, 42, 810–813. [Google Scholar] [CrossRef]
- Zeneli, G.; Krokene, P.; Christiansen, E.; Krekling, T.; Gershenzon, J. Methyl Jasmonate Treatment of Mature Norway spruce (Picea abies) Trees Increases the Accumulation of Terpenoid Resin Components and Protects Against Infection by Ceratocystis polonica, a Bark Beetle-associated Fungus. Tree Physiol. 2006, 26, 977–988. [Google Scholar] [CrossRef]
- Ro, D.K.; Arimura, G.; Lau, S.Y.; Piers, E.; Bohlmann, J. Loblolly pine Abietadienol/Abietadienal Oxidase PtAO (CYP720B1) is a Multifunctional, Multisubstrate Cytochrome P450 Monooxygenase. Proc. Natl. Acad. Sci. USA 2005, 102, 8060–8065. [Google Scholar] [CrossRef] [Green Version]
- Hamberger, B.; Ohnishi, T.; Hamberger, B.; Séguin, A.; Bohlmann, J. Evolution of Diterpene Metabolism: Sitka Spruce CYP720B4 Catalyzes Multiple Oxidations in Resin Acid Biosynthesis of Conifer Defense against Insects. Plant Physiol. 2011, 157, 1677–1695. [Google Scholar] [CrossRef] [Green Version]
- Pham, T.; Chen, H.; Dai, L.; Vu, T.Q.T. Isolation a P450 Gene in Pinus armandi and Its Expression after Inoculation of Leptographium qinlingensis and Treatment with Methyl Jasmonate. Russ. J. Plant Physiol. 2016, 63, 118–125. [Google Scholar] [CrossRef]
- Chen, Y.L.; Kang, L.H.; Malajczuk, N.; Dell, B. Selecting Ectomycorrhizal Fungi for Inoculating Plantations in South China: Effect of Scleroderma on Colonization and Growth of Exotic Eucalyptus globulus, E. urophylla, Pinus elliottii, and P. radiata. Mycorrhiza 2006, 16, 251–259. [Google Scholar] [CrossRef]
- Rodrigues, K.C.S.; Fett-Neto, A.G. Oleoresin Yield of Pinus elliottii in a Subtropical Climate: Seasonal Variation and Effect of Auxin and Salicylic Acid-Based Stimulant Paste. Ind. Crop. Prod. 2009, 30, 316–320. [Google Scholar] [CrossRef]
- Min, Y.; Ting, J.; Leiming, D.; Lu, Z.; Chunhui, L.; Siyu, L.; Meng, L. Resin Yield in Pinus elliottii Engelm is Related to the Resin Flow Rate, Resin Components and Resin Duct Characteristics at Three Locations in Southern China. Ind. Crop. Prod. 2020, 160, 113141. [Google Scholar] [CrossRef]
- Westbrook, J.W.; Walker, A.R.; Neves, L.G.; Munoz, P.; Resende, M.F., Jr.; Neale, D.B.; Wegrzyn, J.L.; Huber, D.A.; Kirst, M.; Davis, J.M.; et al. Discovering Candidate Genes that Regulate Resin Canal Number in Pinus taeda Stems by Integrating Genetic Analysis across Environments, Ages, and Populations. New Phytol. 2015, 205, 627–641. [Google Scholar] [CrossRef] [Green Version]
- Diao, S.; Ding, X.; Luan, Q.; Jiang, J. A Complete Transcriptional Landscape Analysis of Pinus elliottii Engelm. Using Third-Generation Sequencing and Comparative Analysis in the Pinus Phylogeny. Forests 2019, 10, 942. [Google Scholar] [CrossRef] [Green Version]
- Zimin, A.V.; Stevens, K.A.; Crepeau, M.W.; Puiu, D.; Wegrzyn, J.L.; Yorke, J.A.; Langley, C.H.; Neale, D.B.; Salzberg, S.L. An Improved Assembly of the Loblolly pine Mega-genome Using Long-read Single-molecule Sequencing. Gigascience 2017, 6, 1–4. [Google Scholar] [CrossRef]
- De, L.; Fernanda, D.C.; Füller, T.N.; Silva, R.; Kerber, M.R.; Lima, M.S.; Fett, J.P.; Fett-Neto, A.G. Reference Genes for qPCR Analysis in Resin-Tapped Adult Slash Pine As a Tool to Address the Molecular Basis of Commercial Resinosis. Front. Plant Sci. 2016, 7, 849. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Wang, H.; Sathyan, P.; Stasolla, C.; Loopstra, C.A. Real-time RT–PCR Analysis of Loblolly pine (Pinus taeda) Arabinogalactan-protein and Arabinogalactan-protein-like Genes. Physiol. Plantarum. 2005, 124, 91–106. [Google Scholar] [CrossRef]
- Zhao, S.; Fernald, R.D. Comprehensive Algorithm for Quantitative Real-time Polymerase Chain Reaction. J. Comput. Biol. 2005, 12, 1047–1064. [Google Scholar] [CrossRef] [PubMed]
- Rieu, I.; Powers, S.J. Real-Time Quantitative RT–PCR: Design, Calculations, and Statistics. Plant Cell 2009, 21, 1031–1033. [Google Scholar] [CrossRef] [Green Version]
- Robert, J.A.; Madilao, L.L.; White, R.; Yanchuk, A.; King, J.; Bohlmann, J. Terpenoid Metabolite Profiling in Sitka spruce Identifies Association of Dehydroabietic acid, (+)-3-carene, and Terpinolene with Resistance Against White pine weevil. Botany 2010, 88, 810–820. [Google Scholar] [CrossRef]
- Kalb, V.F.; Loper, J.C. Proteins from Eight Eukaryotic Cytochrome P-450 Families Share a Segmented Region of Sequence Similarity. Proc. Nat. Acad. Sci. USA 1988, 85, 1721–1725. [Google Scholar] [CrossRef] [Green Version]
- Choe, S. The DWF4 Gene of Arabidopsis Encodes a Cytochrome P450 That Mediates Multiple 22alpha-hydroxylation Steps in Brassinosteroid Biosynthesis. Plant Cell 1998, 10, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, L.; Zhang, X.; Xiwu, G. Molecular Cloning and Recombinant Expression of Cytochrome P450 CYP6B6 from Helicoverpa armigera in Escherichia coli. Mol. Biol. Rep. 2013, 40, 1211–1217. [Google Scholar] [CrossRef]
- Rupasinghe, S.; Schuler, M.A.; Kagawa, N.; Yuan, H.; Lei, L.; Zhao, B.; Kelly, S.L.; Waterman, M.R.; Lamb, D.C. The Cytochrome P450 Gene Family CYP157 Does Not Contain EXXR in the K-helix Reducing the Absolute Conserved P450 Residues to a Single Cysteine. FEBS Lett. 2006, 580, 6338–6342. [Google Scholar] [CrossRef] [Green Version]
- Neale, D.B.; Wegrzyn, J.L.; Stevens, K.A.; Zimin, A.V.; Puiu, D.; Crepeau, M.W.; Cardeno, C.; Koriabine, M.; Holtz-Morris, A.E.; Liechty, J.D.; et al. Decoding the massive genome of loblolly pine using haploid dna and novel assembly strategies. Genome Biol. 2014, 15, R59. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.Q.; Jian, H.J.; Yang, B.; Lu, K.; Zhang, A.X.; Liu, P. Genome-wide analysis and expression profiling of the grf gene family in oilseed rape (Brassica napus L.). Gene 2017, 620, 36–45. [Google Scholar] [CrossRef]
- Bathe, U.; Tissier, A. Cytochrome P450 Enzymes: A Driving Force of Plant Diterpene Diversity. Phytochemistry 2019, 161, 149–162. [Google Scholar] [CrossRef]
- Godard, K.; Byun-Mckay, A.; Levasseur, C.; Plant, A.; Séguin, A.; Bohlmann, J. Testing of a Heterologous, Wound- and Insect-inducible Promoter for Functional Genomics Studies in Conifer Defense. Plant Cell Rep. 2007, 26, 2083–2090. [Google Scholar] [CrossRef]
- Forman, V.; Callari, R.; Folly, C.; Heider, H.; Hamberger, B. Production of Putative Diterpene Carboxylic Acid Intermediates of Triptolide in Yeast. Molecules 2017, 22, 981. [Google Scholar] [CrossRef] [Green Version]
- Enrica, A.; Stefano, C.; Bartolomeo, S.; Rita, P.A.; Maurizio, B.; Francesco, M.; Peter, B.C.; Agostino, S.; Mario, C. Diterpene Resin Acids and Olefins in Calabrian Pine (Pinus nigra subsp. laricio (Poiret) Maire) Oleoresin: GC-MS Profiling of Major Diterpenoids in Different Plant Organs, Molecular Identification and Expression Analysis of Diterpene Synthase Genes. Plants 2021, 10, 2391. [Google Scholar] [CrossRef]
- Simón, B.F.D.; Aranda, I.; López-Hinojosa, M.; Miguel, L.; Cervera, M.T. Scion-rootstock Interaction and Drought Systemic Effect Modulate the Organ-specific Terpene Profiles in Grafted Pinus pinaster Ait. Environ. Exp. Bot. 2021, 186, 104437. [Google Scholar] [CrossRef]
- Huang, A.C.; Osbourn, A. Plant Terpenes that Mediate Below-ground Interactions: Prospects for Bioengineering Terpenoids for Plant Protection. Pest Manag. Sci. 2019, 75, 2368–2377. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, M.M.; Wang, Q.; Webster, F.X.; Kiemle, D.; Hong, Y.J.; Tantillo, D.J.; Coates, R.M.; Wray, A.T.; Askew, W.; O’Donnell, C.; et al. Formation of the Unusual Semivolatile Diterpene Rhizathalene by the Arabidopsis Class I Terpene Synthase TPS08 in the Root Stele Is Involved in Defense against Belowground Herbivory. Plant Cell 2013, 25, 1108–1125. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Galhano, R.; Wiemann, P.; Bueno, E.; Tiernan, M.; Wu, W.; Chung, I.M.; Gershenzon, J.; Tudzynski, B.; Sesma, A.; et al. Genetic evidence for natural product-mediated plant–plant allelopathy in rice (Oryza sativa). New Phytol. 2012, 193, 570–575. [Google Scholar] [CrossRef] [Green Version]


| Species | Gene ID | Protein Length (aas) | Molecular Weight (MolWt) | Isoelectric Point (pI) | Gene ID | Protein Length (aas) | Molecular Weight (MolWt) | Isoelectric Point (pI) |
|---|---|---|---|---|---|---|---|---|
| Pinus elliottii | c330768 | 479 | 55,417.44 | 9.5 | c323216 | 477 | 55,019.62 | 8.23 |
| c161262 | 484 | 56,135.81 | 8.74 | c305297 | 486 | 56,193.09 | 9.48 | |
| c327173 | 487 | 56,246.2 | 9.57 | c327174 | 487 | 56,250.19 | 9.36 | |
| c327173i1 | 470 | 54,233.74 | 9.58 | c327174i1 | 447 | 51,369.63 | 9.53 | |
| c324680v1 | 456 | 52,894.31 | 9.08 | c356010v1 | 470 | 54,039.35 | 8.63 | |
| c324680v2i2 | 402 | 46,469.52 | 7.93 | c356010v2 | 481 | 55,656.16 | 8.11 | |
| c324680i1 | 464 | 53,577.96 | 8.46 | c305557v1 | 486 | 56,469.38 | 9.29 | |
| c305557v2 | 486 | 56,499.45 | 9.15 | c305557v3 | 486 | 56,489.41 | 9.15 | |
| c305557i1 | 447 | 51,912.93 | 8.47 | |||||
| Pinus taeda | PITA_49896 | 489 | 56,291.24 | 9.6 | PITA_14112 | 497 | 57,223.98 | 8.56 |
| PITA_42322 | 431 | 49,609.36 | 9.64 | PITA_06980 | 457 | 52,413.8 | 9.7 | |
| PITA_11046 | 497 | 56,972.88 | 8.19 | PITA_02864 | 511 | 58,378.87 | 9.04 | |
| PITA_10468 | 478 | 54,899.42 | 7.89 | PITA_43262 | 403 | 46,434.27 | 7.45 | |
| PITA_22834 | 487 | 55,925.77 | 9.36 | PITA_17539v1 | 482 | 55,576.02 | 7.88 | |
| PITA_40733v1 | 487 | 56,629.66 | 9.29 | PITA_17539i1 | 453 | 52,353.47 | 8.17 | |
| PITA_40733v2 | 487 | 56,495.54 | 9.48 | PITA_17539v2 | 482 | 55,646.18 | 8.07 | |
| PITA_37500v1 | 487 | 56,041.91 | 9.51 | PITA_37500v4 | 487 | 56,085.96 | 9.51 | |
| PITA_37500v2 | 487 | 56,075.92 | 9.51 | PITA_37500v5 | 487 | 56,013.85 | 9.51 | |
| PITA_37500v3 | 487 | 56,055.93 | 9.51 |
| Clade | Gene ID | (P/I)PGSx(G/P)xP | AGx(D/E)T | ExxR | PxRx | FxxGxxxCxG |
|---|---|---|---|---|---|---|
| Clade I | c161262 | PPGSRGWP | AGHET | ETLR | PWRW | FGGGARLCPG |
| c305297 | PPGSRGWP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| c305557v1 | PPGSSGWP | AGHET | ETHR | PWRW | FGGGLRLCPG | |
| c305557i1 | PPGSSGWP | AGHET | ETHR | PWRW | FGGGLRLCPG | |
| c305557v2 | PPGSSGWP | AGHET | ETHR | PWRW | FGGGLRLCPG | |
| c305557v3 | PPGSSGWP | AGHET | ETHR | PWRW | FGGGLRLCPG | |
| c327173 | PPGSTGLP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| c327173i1 | PPGSTGLP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| c327174 | PPGSTGWP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| c327174i1 | PPGSTGWP | AGHET | --LR | PWRW | FGGGARLCPG | |
| PITA_06980 | PPGSTGWP | AGHET | ETLR | PWRW | FGGGPRLCPG | |
| PITA_14112 | PPGSTGWP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| PITA_22834 | PAGSRGWP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| PITA_49896 | PPGSTGLP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| PITA_37500v1 | PPGSRGWP | AGHET | ETLR | PWRW | FGSGARLCPG | |
| PITA_37500v2 | PPGSRGWP | AGHET | ETLR | PWRW | FGSGARFCPG | |
| PITA_37500v3 | PPGSRGWP | AGHET | ETLR | PWRW | FGSGARLCPG | |
| PITA_37500v4 | PPGSRGWP | AGHET | ETLR | PWRW | FGSGARLCPG | |
| PITA_37500v5 | PPGSRGWP | AGHET | ETLR | PWRW | FGSGARLCPG | |
| PITA_40733v1 | PPGSSGWP | AGHET | ETHR | PWRW | FGGGLRLCPG | |
| PITA_40733v2 | PPGSSGWP | AGHET | ETHR | PWRW | FGGGLRLCPG | |
| Clade III | c323216 | PPGSTGWP | AGHET | ETLR | PWRW | FGGGARLCPG |
| c324680v1 | PPGSTGWP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| c324680i1 | PPGSTGWP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| c324680v2i2 | PPGSTGWP | ----- | ETLR | PWRW | FGGGARLCPG | |
| c356010v1 | PPGSTGWP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| c356010v2 | PPGSTGWP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| PITA_10468 | PPGSTGWP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| PITA_11046 | PPGSTGWP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| PITA_43262 | PPGSTGWP | ----- | ETLR | PWRW | FGEGARLCPG | |
| PITA_17539v1 | PTGSTGWP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| PITA_17539i1 | PPGSTGWP | AGHET | EILR | PWRW | FGGGARLCPG | |
| PITA_17539v2 | PTGSTGWP | AGHET | ETLR | PWRW | FGGGARLCPG | |
| Clade II | PITA_42322 | PPGSTGWP | ----T | ---- | PSRW | FGGGARLCPG |
| Clade IV | c330768 | PPGSTGWP | AG-QT | ETLR | PWRW | FGAGARLCPG |
| PITA_02864 | PPGSTGWP | AG-QT | ETLR | PWRW | FGAGARLCPG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Ding, X.; Luan, Q.; Jiang, J.; Diao, S. Identification and Tissue-Specific Expression Analysis of CYP720B Subfamily Genes in Slash Pine and Loblolly Pine. Forests 2022, 13, 283. https://doi.org/10.3390/f13020283
Zhang Y, Ding X, Luan Q, Jiang J, Diao S. Identification and Tissue-Specific Expression Analysis of CYP720B Subfamily Genes in Slash Pine and Loblolly Pine. Forests. 2022; 13(2):283. https://doi.org/10.3390/f13020283
Chicago/Turabian StyleZhang, Yini, Xianyin Ding, Qifu Luan, Jingmin Jiang, and Shu Diao. 2022. "Identification and Tissue-Specific Expression Analysis of CYP720B Subfamily Genes in Slash Pine and Loblolly Pine" Forests 13, no. 2: 283. https://doi.org/10.3390/f13020283
APA StyleZhang, Y., Ding, X., Luan, Q., Jiang, J., & Diao, S. (2022). Identification and Tissue-Specific Expression Analysis of CYP720B Subfamily Genes in Slash Pine and Loblolly Pine. Forests, 13(2), 283. https://doi.org/10.3390/f13020283