
Mobile Messenger RNAs in Grafts of Salix matsudana Are Associated with Plant Rooting

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Grafting
2.2. Extraction of Total RNA
2.3. RNA Library Construction and Sequencing
2.4. Identification of the Differentially Expressed Genes (DEGs) and Function Analysis
2.5. Determination of the Transfer RNAs
2.6. Estimation of RNA Transfer Ratio
3. Results
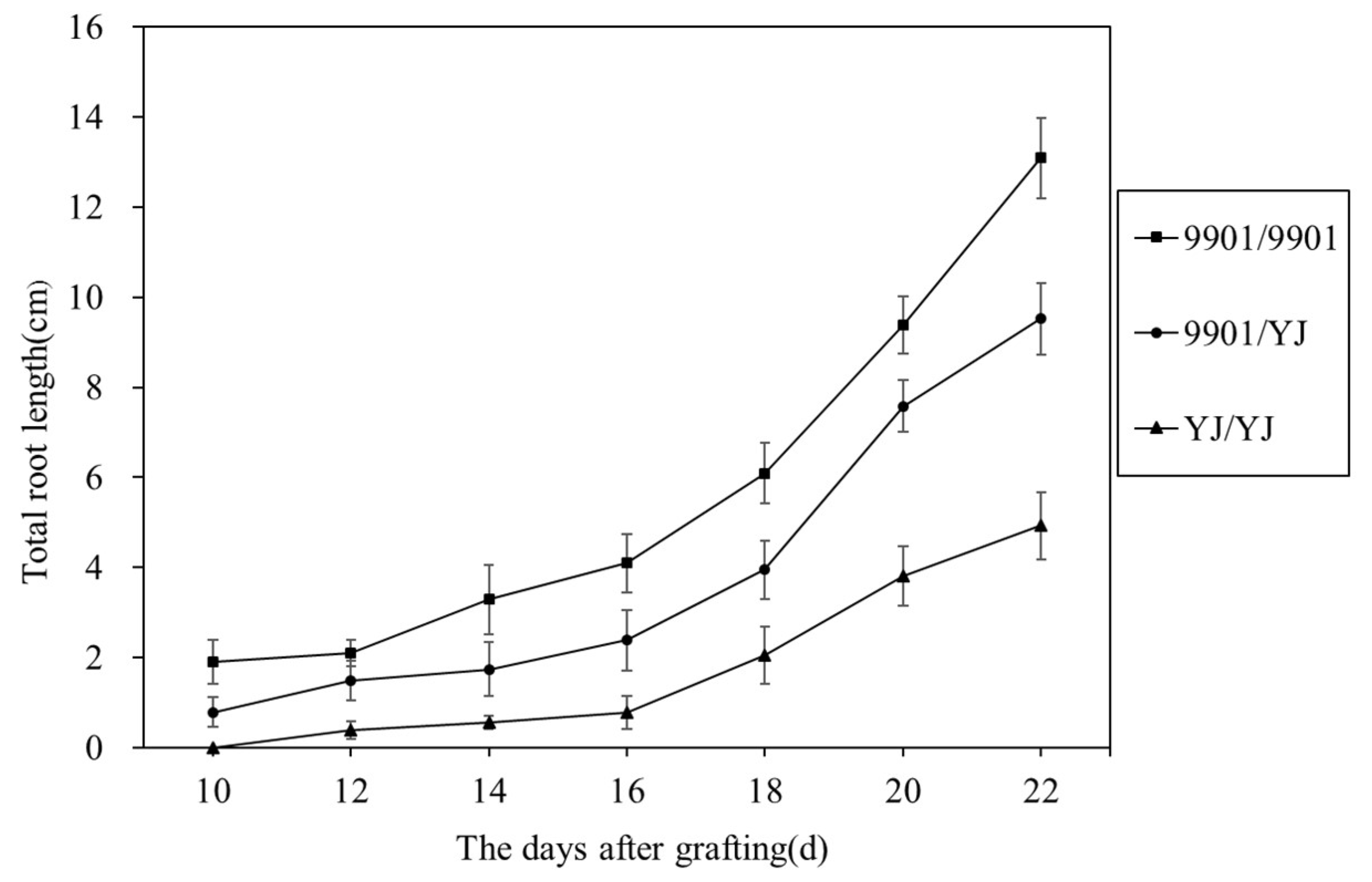
3.1. 9901 Scions Enhanced the Rooting of YJ Rootstocks in 9901/YJ Grafts
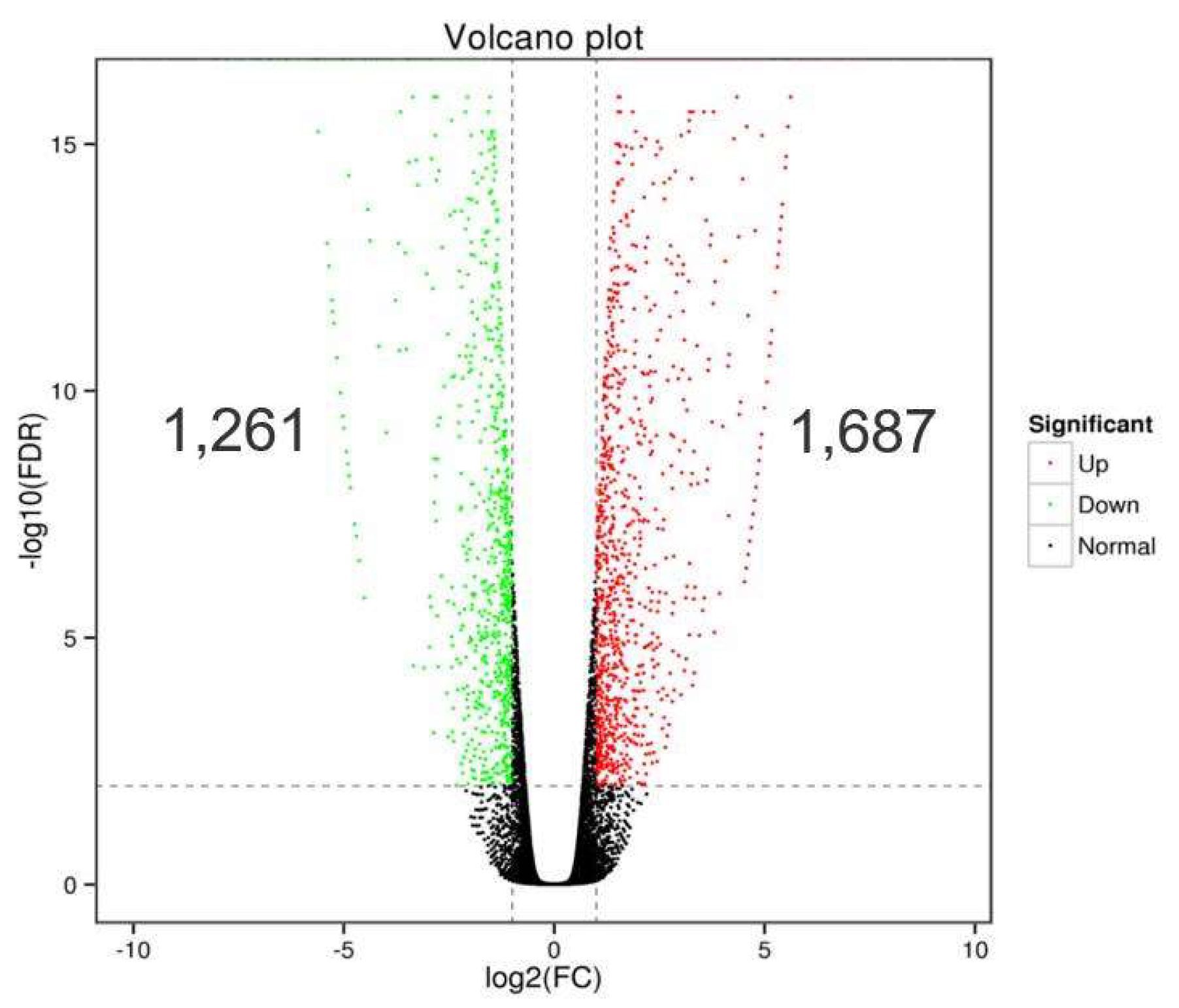
3.2. DEGs in Homologous and Heterologous Willow Grafts
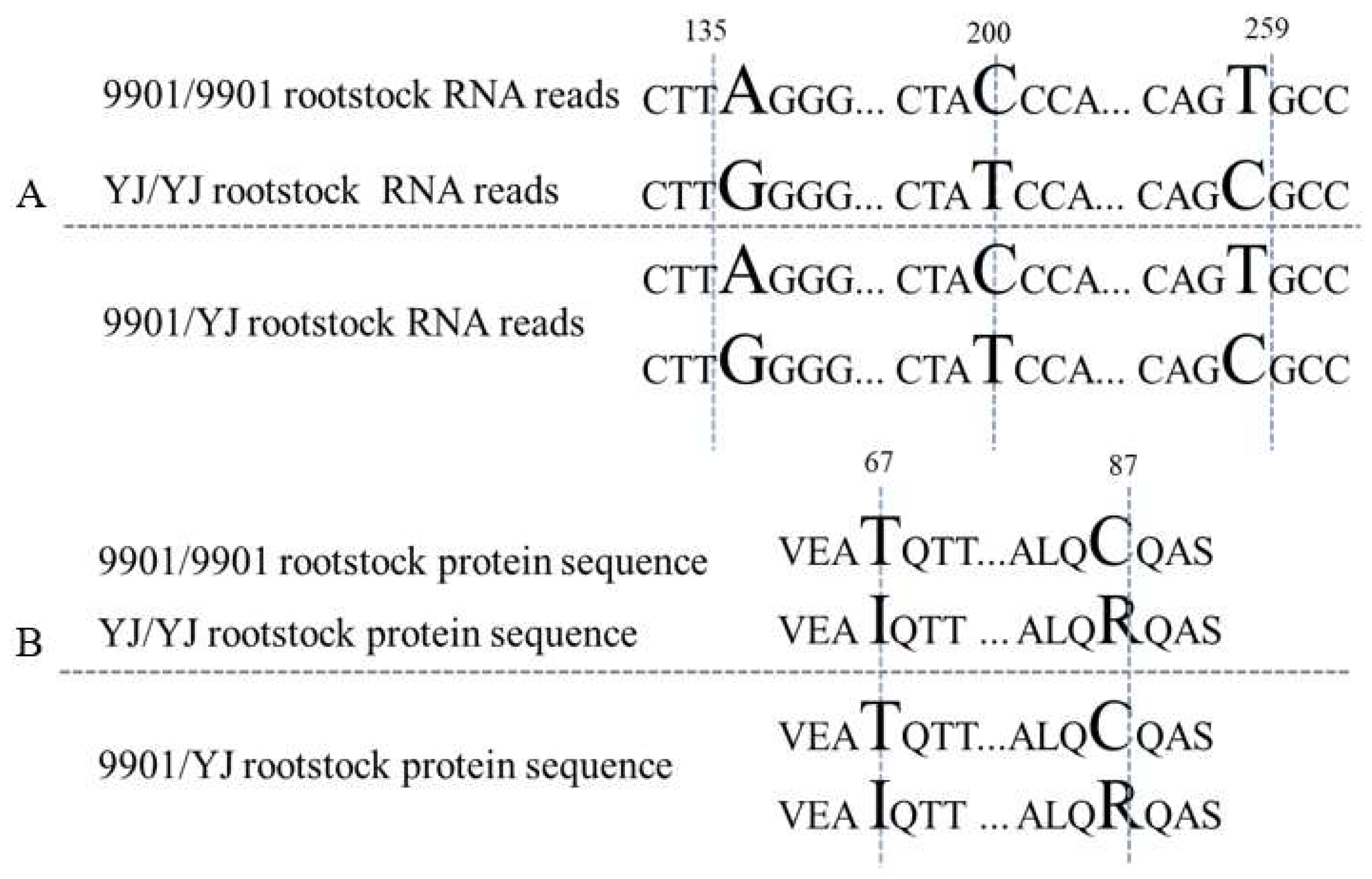
3.3. SNP Analysis Revealed 692 Shoot-to-Root Mobile mRNAs in 9901/YJ Grafts
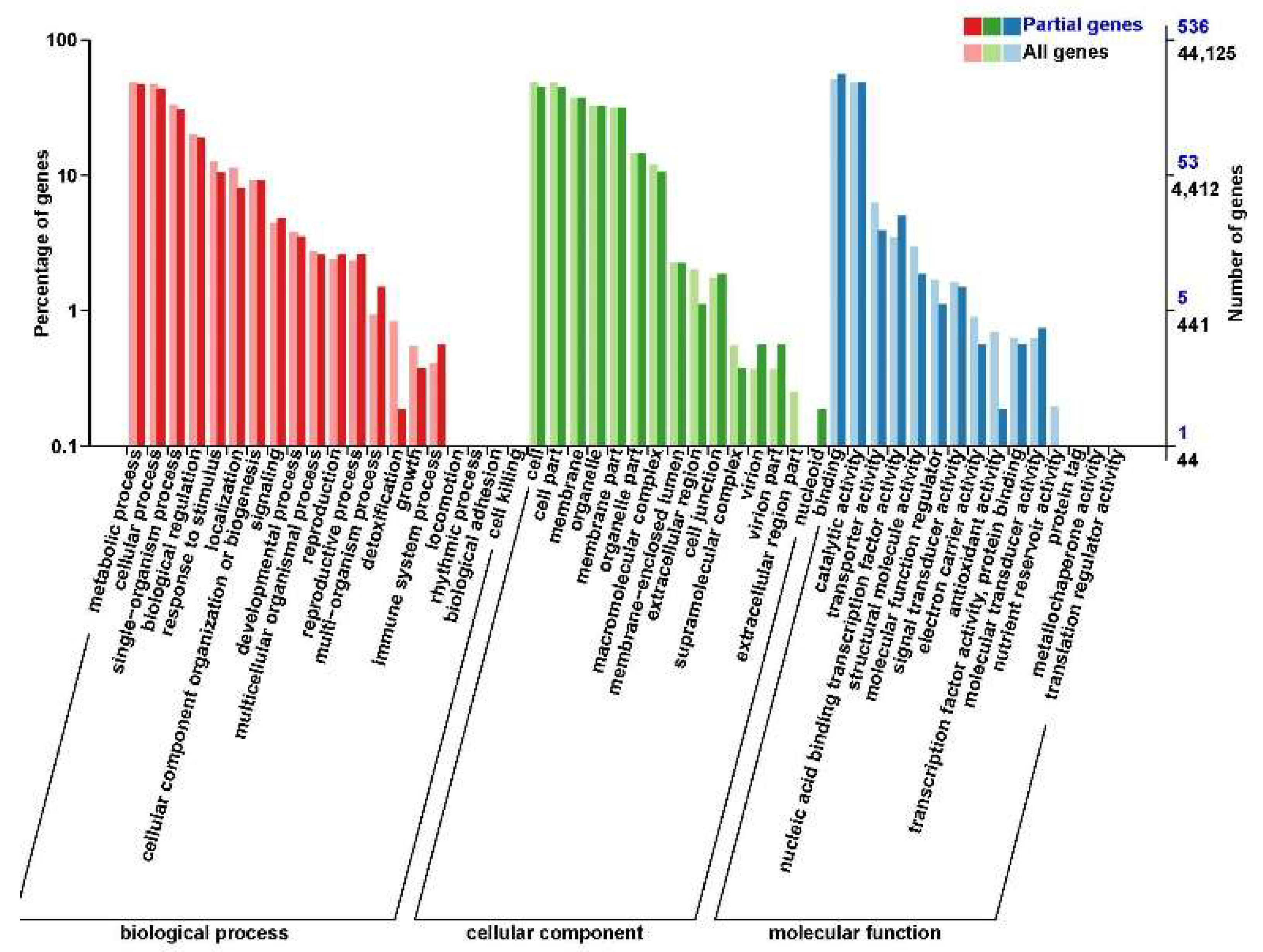
3.4. Characterization of the Mobile mRNAs in Grafted Willows
3.5. The Mobile mRNAs and Pathways Involved in Roots Formation
4. Discussion
4.1. The Number of Mobile mRNAs Varied Vastly in Different Grafts
4.2. The Mobile mRNAs Efficiently Affect Plant Performance
4.3. The Mobile mRNAs in Grafts Are of Some Sequence Characteristics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Darikova, J.A.; Savva, Y.V.; Vaganov, E.A.; Grachev, A.M.; Kuznetsova, G.V. Grafts of woody plants and the problem of incompatibility between scion and rootstock. J. Sib. Fed. Univ. Biol. 2011, 4, 54–63. [Google Scholar]
- Haroldsen, V.M.; Szczerba, M.W.; Aktas, H.; Lopez-Baltazar, J.; Odias, M.J.; Chi-Ham, C.L.; Labavitch, J.M.; Bennett, A.B.; Powell, A.L.T. Mobility of transgenic nucleic acids and proteins within grafted rootstocks for agricultural improvement. Front. Plant Sci. 2012, 3, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Canio, W.; Kessler, S.; Sinha, N. Developmental changes due to long-distance movement of a homeobox fusion mRNA in tomato. Science 2001, 293, 287–289. [Google Scholar] [CrossRef] [PubMed]
- Haywood, V.; Yu, T.; Huang, N.; Lucas, W.J. Phloem long-distance trafficking of gibberellic acid-insensitive RNA regulates leaf development. Plant J. 2005, 42, 49–68. [Google Scholar] [CrossRef] [PubMed]
- Gaion, L.A.A.; Braz, L.T.; Carvalho, R.F. Grafting in vegetable crops: A great technique for agriculture. Int. J. Veg. Sci. 2018, 24, 85–102. [Google Scholar] [CrossRef]
- Sanchez-Rodríguez, E.; Rubio-Wilhelmi, M.M.; Blasco, B.; Leyva, R.; Romero, L.; Ruiz, J.M. Antioxidant response resides in the shoot in reciprocal grafts of drought-tolerant and drought-sensitive cultivars in tomato under water stress. Plant Sci. 2012, 188–189, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, J.; Huang, L.; Ding, X.; Shi, K.; Yu, J. Grafting of Cucumis sativus onto Cucurbita ficifolia leads to improved plant growth, increased light utilization and reduced accumulation of reactive oxygen species in chilled plants. J. Plant Res. 2009, 122, 529–540. [Google Scholar] [CrossRef]
- Savvas, D.; Colla, G.; Rouphael, Y.; Dietmar, S. Amelioration of heavy metal and nutrient stress in fruit vegetables by grafting. Sci. Hortic. 2010, 127, 156–161. [Google Scholar] [CrossRef]
- Banerjee, A.K.; Chatterjee, M.; Yu, Y.; Suh, S.; Miller, W.A.; Hannapel, D.J. Dynamics of a mobile RNA of potato involved in a long-distance signaling pathway. Plant Cell. 2006, 18, 3443–3457. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Mao, L.; Jittayasothorn, Y.; Kang, Y.; Jiao, C.; Fei, Z.; Zhong, G. Messenger RNA exchange between scions and rootstocks in grafted grapevines. BMC Plant Biol. 2015, 15, 251. [Google Scholar] [CrossRef] [Green Version]
- Thieme, C.J.; Rojas-Triana, M.; Stecyk, E.; Schudoma, C.; Zhang, W.; Yang, L.; Miñambres, M.; Walther, D.; Schulze, W.X.; Paz-Ares, J.; et al. Endogenous Arabidopsis messenger RNAs transported to distant tissues. Nat. Plants 2015, 1, 15025. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zheng, Y.; Ham, B.; Chen, J.; Yoshida, A.; Kochian, L.V.; Fei, Z.; Lucas, W.J. Vascular-mediated signalling involved in early phosphate stress response in plants. Nat. Plants 2016, 2, 16033. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Zheng, Y.; Huang, J.; Zhou, X.; Li, R.; Zhao, M.; Wang, S.; Huang, Z.; Lan, H.; Turgeon, R.; et al. Elucidation of the mechanisms of long-distance mRNA movement in a Nicotiana benthamiana/tomato heterograft system. Plant Physiol. 2018, 177, 745–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, 14. [Google Scholar] [CrossRef] [Green Version]
- Gotz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talon, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3425. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Sukumar, P.; Maloney, G.S.; Muday, G.K. Localized induction of the ATP-binding cassette B19 auxin transporter enhances adventitious root formation in Arabidopsis. Plant Physiol. 2013, 162, 1392–1405. [Google Scholar] [CrossRef]
- Cheng, X.; Jiang, H.; Zhang, J.; Qian, Y.; Zhu, S.; Cheng, B. Overexpression of type-A rice response regulators, OsRR3 and OsRR5, results in lower sensitivity to cytokinins. Genet. Mol. Res. 2010, 9, 348–359. [Google Scholar] [CrossRef]
- Zhao, Y.; Cheng, S.; Song, Y.; Huang, Y.; Zhou, S.; Liu, X.; Zhou, D. The interaction between rice ERF3 and WOX11 promotes crown root development by regulating gene expression involved in cytokinin signaling. Plant Cell 2015, 27, 2469–2483. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kang, J.; Cho, D.; Park, J.H.; Kim, S.Y. ABF2, an ABRE-binding bZIP factor, is an essential component of glucose signaling and its overexpression affects multiple stress tolerance. Plant J. 2004, 40, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Lozano, M.; Dutta, S.K.; Natarajan, P.; Tomason, Y.R.; Lopez, C.; Katam, R.; Levi, A.; Nimmakayala, P.; Reddy, U.K. Transcriptome changes in reciprocal grafts involving watermelon and bottle gourd reveal molecular mechanisms involved in increase of the fruit size, rind toughness and soluble solids. Plant Mol. Biol. 2020, 102, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Notaguchi, M.; Higashiyama, T.; Suzuki, T. Identification of mRNAs that move over long distances using an RNA-seq analysis of Arabidopsis/Nicotiana benthamiana heterografts. Plant Cell Physiol. 2015, 56, 311–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Liu, C.; Li, Z.; Ran, Q.; Xie, G.; Wang, B.; Fang, S.; Chu, J.; Zhang, J. ZmbZIP4 contributes to stress resistance in maize by regulating ABA synthesis and root development. Plant Physiol. 2018, 178, 753–770. [Google Scholar] [CrossRef] [Green Version]
- Kanehira, A.; Yamada, K.; Iwaya, T.; Tsuwamoto, R.; Kasai, A.; Mikio, N.; Harada, T. Apple phloem cells contain some mRNAs transported over long distances. Tree Genet. Genom. 2010, 6, 635–642. [Google Scholar] [CrossRef]
- Omid, A.; Keilin, T.; Glass, A.; Leshkowitz, D.; Wolf, S. Characterization of phloem-sap transcription profile in melon plants. J. Exp. Bot. 2007, 58, 3645–3656. [Google Scholar] [CrossRef] [Green Version]
- Hellmann, H.; Estelle, M. Plant development: Regulation by protein degradation. Science 2002, 297, 793–797. [Google Scholar] [CrossRef] [Green Version]
- Maraschin, F.S.; Memelink, J.; Offringa, R. Auxin-induced, SCFTIR1-mediated poly-ubiquitination marks AUX/IAA proteins for degradation. Plant J. 2009, 59, 100–109. [Google Scholar] [CrossRef]
- Li, J.; Dai, X.; Zhao, Y. A role for auxin response factor 19 in auxin and ethylene signaling in Arabidopsis. Plant Physiol. 2006, 140, 899–908. [Google Scholar] [CrossRef] [Green Version]
- Calderwood, A.; Kopriva, S.; Morris, R. Transcript abundance explains mRNA mobility data in Arabidopsis thaliana. Plant Cell 2016, 28, 610–615. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Thieme, C.; Kollwig, G.; Apelt, F.; Yang, L.; Winter, N.; Andresen, N.; Walther, D.; Kragler, F. tRNA-related sequences trigger systemic mRNA transport in plants. Plant Cell 2016, 28, 1237–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Yu, P.; Zhao, J.; Jiang, H.; Wang, H.; Zhu, Y.; Botella, M.A.; Samaj, J.; Li, C.; Lin, J. Expression of tomato prosystemin gene in Arabidopsis reveals systemic translocation of its mRNA and confers necrotrophic fungal resistance. New Phytol. 2018, 217, 799–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
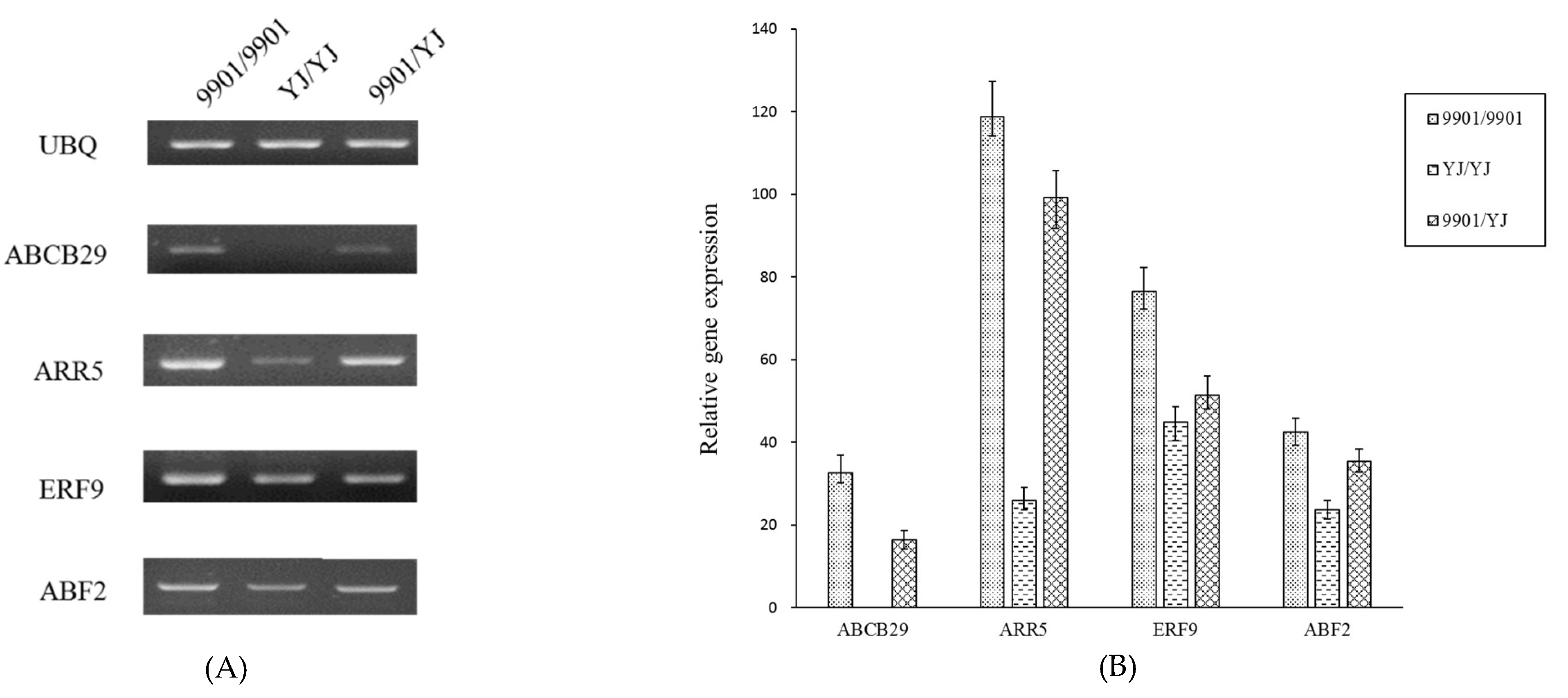{kind=link}
| mRNA Length (bp) | Mobile mRNA Amount | Total Transcribed Gene Amount | Mobile mRNA Ratio in Total Transcribed Genes of the Group (%) a | Mobile mRNA Propotion in All Mobile mRNAs (%) b |
|---|---|---|---|---|
| 0–500 | 44 | 4883 | 0.90 | 9.67 |
| 501–1000 | 96 | 12,127 | 0.79 | 21.10 |
| 1001–1500 | 121 | 11,219 | 1.08 | 26.59 |
| 1501–2000 | 65 | 6441 | 1.01 | 14.29 |
| 2001–2500 | 46 | 3444 | 1.34 | 10.11 |
| 2501–3000 | 35 | 1901 | 1.84 | 7.69 |
| 3001–3500 | 17 | 1151 | 1.48 | 3.74 |
| 3501–4000 | 11 | 558 | 1.97 | 2.42 |
| 4001– | 20 | 835 | 2.40 | 4.39 |
| GC Content (%) | Mobile mRNA Amount | Total Transcribed Gene Amount | Mobile mRNA Ratio in Total Transcribed Gene of the Group (%) | Mobile mRNA Proportion in All Mobile mRNA (%) |
|---|---|---|---|---|
| 25–30 | 0 | 2 | 0 | 0 |
| 30–35 | 0 | 72 | 0 | 0 |
| 35–40 | 13 | 1353 | 0.96 | 2.86 |
| 40–45 | 292 | 25,776 | 1.13 | 64.18 |
| 45–50 | 128 | 13,471 | 0.95 | 28.13 |
| 50–55 | 21 | 1752 | 1.2 | 4.62 |
| 55– | 1 | 133 | 0.75 | 0.21 |
| mRNA Expression Abundance (FPKM) | Mobile mRNAs Amount | Total Transcribed Gene Amount | Mobile mRNAs Ratio in Total Transcribed Gene of the Group (%) | Mobile mRNAs Proportion in All Mobile mRNAs (%) |
|---|---|---|---|---|
| 0–10 | 288 | 33,241 | 0.87 | 63.30 |
| 10–20 | 96 | 7113 | 1.12 | 21.10 |
| 20–30 | 39 | 3168 | 0.85 | 8.57 |
| 30–40 | 16 | 1734 | 0.69 | 3.52 |
| 40–50 | 12 | 1077 | 1.11 | 2.63 |
| 50–60 | 4 | 631 | 0.63 | 0.88 |
| Pathway | ko-id | p-Value | Mobile mRNAs Amount |
|---|---|---|---|
| Spliceosome | ko03040 | 0.0606 | 11 |
| Biosynthesis of amino acids | ko01230 | 0.1549 | 11 |
| Plant-pathogen interaction | ko04626 | 0.0588 | 10 |
| Protein processing in endoplasmic reticulum | ko04141 | 0.1816 | 10 |
| Plant hormone signal transduction | ko04075 | 0.5378 | 8 |
| RNA transport | ko03013 | 0.3316 | 7 |
| Carbon metabolism | ko01200 | 0.7074 | 7 |
| Ribosome | ko03010 | 0.9146 | 7 |
| Fructose and mannose metabolism | ko00051 | 0.1093 | 4 |
| Carbon fixation in photosynthetic organisms | ko00710 | 0.2043 | 4 |
| Mobile mRNAs | Gene Function | Expression Abundance * | Transfer Ratio | ||
|---|---|---|---|---|---|
| 9901/9901 | YJ/YJ | 9901/YJ | |||
| AUX/IAA (K14484) | activation of the downstream transcription factor ARF | 0.50(0) | 0(0.14) | 0.37(0.26) | 58.73% |
| CRE1 (K14489) | transmembrane histidine kinase cytokinin receptor activity | 0.29(0) | 0(0.03) | 0.12(0.12) | 50.00% |
| B-ARR (K14491) | sequence-specific DNA binding | 0.45(0) | 0(0.11) | 0.04(0.04) | 50.00% |
| TF (K16189) | protein dimerization activity | 0.10(0) | 0(0.09) | 0.04(0.06) | 40.00% |
| ABF2 (K14432) | sequence-specific DNA binding | 4.20(0) | 0(0.84) | 0.08(0.40) | 16.67% |
| ABF5 (K14432) | sequence-specific DNA binding | 0.20(0) | 0(0.31) | 0.03(0.18) | 14.29% |
| JAZ (K13464) | transcription corepressor activity | 0.80(0) | 0(0.31) | 0.28(0.13) | 68.29% |
| NRP1 (K14508) | Regulatory protein | 0.23(0) | 0(0.38) | 0.10(0.37) | 21.28% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, P.; Lan, B.; Liu, X.; Cui, Y.; Wang, Y.; Xu, J. Mobile Messenger RNAs in Grafts of Salix matsudana Are Associated with Plant Rooting. Forests 2022, 13, 354. https://doi.org/10.3390/f13020354
Yin P, Lan B, Liu X, Cui Y, Wang Y, Xu J. Mobile Messenger RNAs in Grafts of Salix matsudana Are Associated with Plant Rooting. Forests. 2022; 13(2):354. https://doi.org/10.3390/f13020354
Chicago/Turabian StyleYin, Peng, Baoliang Lan, Xiao Liu, Yu Cui, Yan Wang, and Jichen Xu. 2022. "Mobile Messenger RNAs in Grafts of Salix matsudana Are Associated with Plant Rooting" Forests 13, no. 2: 354. https://doi.org/10.3390/f13020354
APA StyleYin, P., Lan, B., Liu, X., Cui, Y., Wang, Y., & Xu, J. (2022). Mobile Messenger RNAs in Grafts of Salix matsudana Are Associated with Plant Rooting. Forests, 13(2), 354. https://doi.org/10.3390/f13020354