Abstract
Green alder (Alnus alnobetula) is currently the most expanding shrub species in the Alps. Because dense thickets impair tree establishment, understanding how climate affects shrub growth is essential for predictions of treeline dynamics. We evaluated ring width data from >50 A. alnobetula stems sampled at treeline on Mt. Patscherkofel (Central European Alps, Austria) to identify main climatic drivers and influence of climate warming on radial stem growth (RG). We also compared RG of A. alnobetula with RG of the co-occurring treeline conifer Swiss stone pine (Pinus cembra). We addressed our questions through calculation of response functions and evaluation of climate in years showing exceptional growth deviations. Response function analyses and evaluation of growth trends during 1991–2020 revealed that RG of A. alnobetula is significantly and directly related to summer temperatures. Precipitation in January also showed a direct relationship to RG, indicating effects of frost drought on RG. Surprisingly, nitrogen fixing A. alnobetula showed strikingly lower RG compared to P. cembra, and the latter also responded more strongly to the increase in summer temperature in the course of climate warming. We explain these findings by different carbon allocation strategies, i.e., preference of “vertical” stem growth in late successional P. cembra vs. favoring “horizontal” spread in the pioneer shrub A. alnobetula.
1. Introduction
In the European Alps global change, especially land abandonment and the decrease in grazing pressure in recent decades, initiated expansion of green alder (Alnus alnobetula (Ehrh.) K. Koch = Alnus viridis (Chaix) DC) across the treeline ecotone. A. alnobetula is currently the most expanding shrub species in the Austrian, French, Italian and Swiss Alps (e.g., [1,2,3,4]). Expansion has also been described for various cold biomes in northern tundra ecosystems (e.g., [5,6,7,8]. Although, in the Alps A. alnobetula is naturally restricted to avalanche slide path, screes and steep, north-facing slopes exhibiting high water availability [9,10,11], expansion into subalpine grasslands and well-drained areas [12] is enhanced by its clonal growth, high seed production and the symbiosis with N2-fixing actinobacteria [13,14] and ectomycorrhizal fungi [15]. Expansion of A. alnobetula, which forms 2–4 m tall canopies, causes considerable environmental changes that have mostly a negative effect on the conservation of vascular plant diversity [4,5,16] and drive grasslands and meadows into N-saturated, species-poor shrubland [17]. Although the life span of A. alnobetula individuals is approximately 60 years (maximum age 110 years [18]), it may persist for centuries due to its ability of clonal growth by layering [19].
There is extensive evidence that at high elevation, low temperature during the growing season directly limits metabolic activity, i.e., cell division and differentiation in meristematic tissues, which leads to growth reductions and ultimately to formation of the treeline (e.g., [20,21,22]). There are also reports that range expansion of A. alnobetula is a consequence of recent climate warming [23,24,25]. However, Mitchell and Ruess [23] suggest that in the future, growth of A. alnobetula may be reduced due to increasing constraint of low water availability with ongoing warming trend, especially when combined with lack of rainfall. A. alnobetula belongs to anisohydric species, i.e., it is dehydration tolerant and keeps its stomata open even under high vapour pressure deficit [26,27,28].
There is a growing need to better understand the drivers of growth of A. alnobetula to improve projections of vegetation change at the alpine treeline ecotone. Radial growth indices are known to be valuable long-term measures of overall tree and shrub vigour (e.g., [29,30,31]) and dendroclimatological methods (“response functions”) are frequently applied to identify the climatic factors most closely associated with variations in tree and shrub growth (e.g., [32,33,34,35]). Because shrubs can provide a multi-decadal record of environmental change, analysing growth over time and climate–growth relationships allow determination of the influence of climate warming and climate extremes on growth processes.
Our understanding of shrub growth at the alpine treeline remains incomplete. Therefore, the foremost goal of this study was to determine the main climatic drivers of radial stem growth (RG) of A. alnobetula at the alpine treeline ecotone on Mt. Patscherkofel, where Swiss stone pine (Pinus cembra L.) is the dominating tree species. Plant growth at high elevation is primarily constrained by the prevailing temperature during the growing season (e.g., [21,36]). Therefore, we expected that RG of A. alnobetula (i) is closely related to summer temperature and (ii) high water availability at the start of the growing season. Furthermore, we expected that (iii) the N2-fixing capacity of A. alnobetula enables considerably higher RG than in the co-occurring tree species P. cembra.
2. Materials and Methods
2.1. Study Area
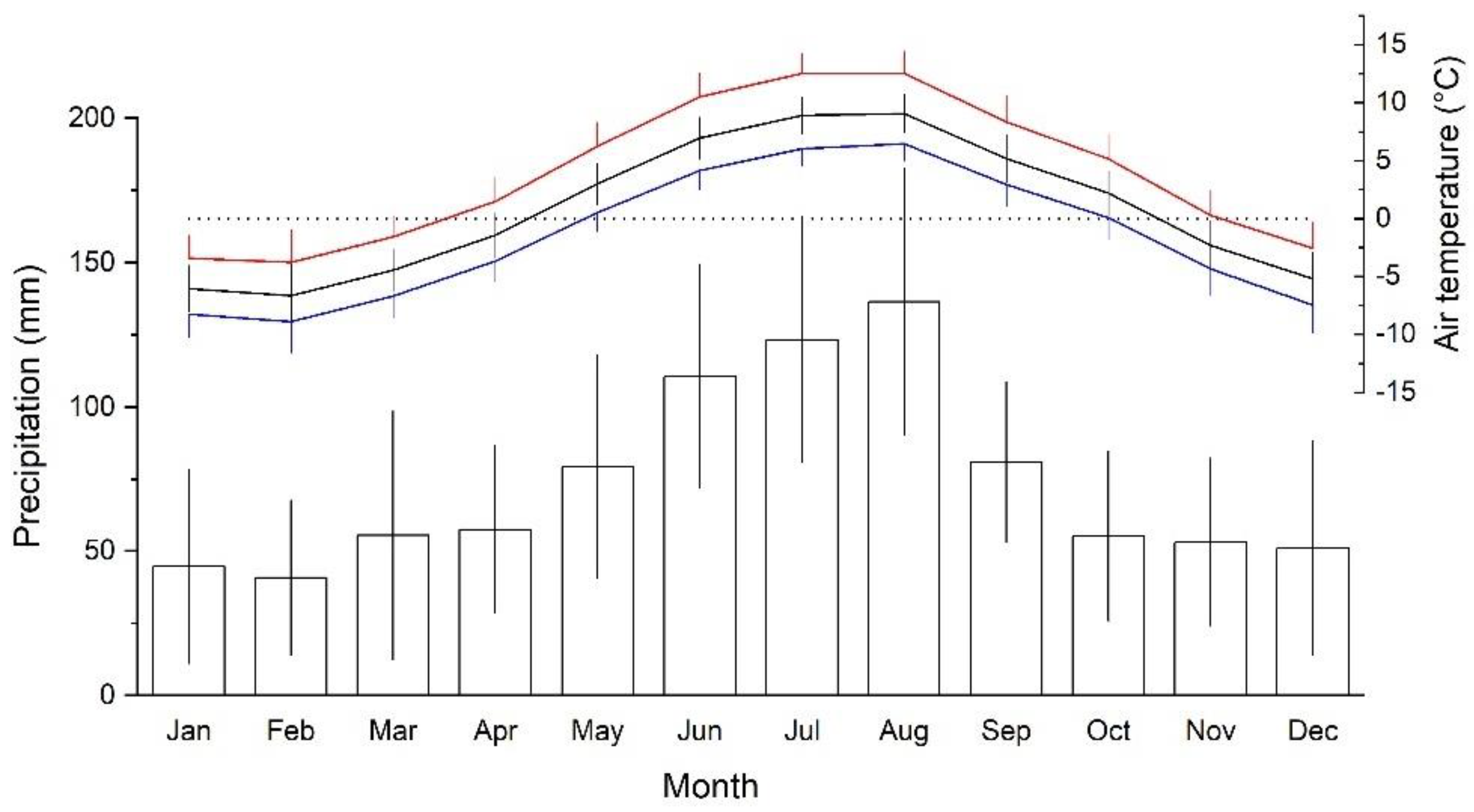
The study area is situated at the treeline ecotone stretching from c. 1950 up to 2200 m on Mt. Patscherkofel (2246 m asl; Tyrol, Austria; 47°12′ N, 11°27′ E). Mt. Patscherkofel belongs to the Central European Alps and is located in an inner alpine dry zone where the local climate is characterized by frequent occurrence of strong southerly winds (Föhn). During the study period 1991–2020, mean annual precipitation at the top of Mt. Patscherkofel (location of meteorological station) was 889 ± 128 mm (mean ± standard deviation, SD). Precipitation maximum is reached during summer (long-term mean (LTM): 371 ± 74 mm during June through August), while winter is the driest season (LTM: 132 ± 60 mm during December through February; Figure 1). Both low winter precipitation and strong winds contribute to frequent occurrence of frost drought effects (i.e., desiccation of above ground organs) across the treeline ecotone in the dominating tree species Swiss stone pine (Pinus cembra; [36,37,38]).
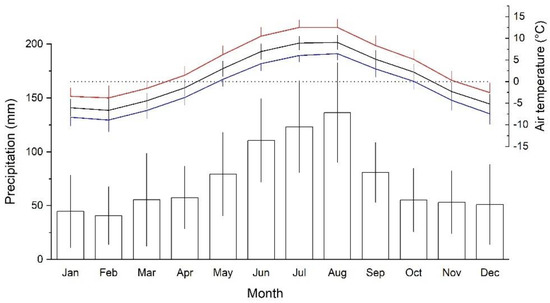
Figure 1.
Mean monthly minimum (blue line), maximum (red line) and mean (black line) daily air temperature and mean monthly precipitation sums (bars) during the study period 1991 to 2020 at the meteorological station on top of Mt. Patscherkofel (2246 m asl). Standard deviations are indicated.
Mean annual temperature during the period 1991–2020 amounted to 0.8 ± 0.7 °C and the coldest and warmest months were February (−6.6 °C) and July (8.9 °C), respectively (Figure 1). Air temperature at 2246 m asl can reach a maximum of 20 °C during summer. Snow depth on Mt. Patscherkofel is mostly less than 1 m but shows pronounced spatial variation due to irregular distribution by frequently occurring strong winds. Whereas south-facing slopes show only small patches of snow in early to mid-April, an almost continuous snow cover persists on steep north-facing slopes from late October until May.
The geology of Mt. Patscherkofel is dominated by gneisses and schists [39] and the soil type within the study area is classified as haplic podzol [40,41]. Alnus alnobetula stands are developed along the treeline ecotone primarily in leeward avalanche gullies but they also occur at wind exposed sites on south-to south-east facing slopes. This tall shrub forms dense monospecific thickets with mean canopy height of c. 2 m.
2.2. Stem Sampling and Growth Ring Measurements
A. alnobetula shows clonal growth leading to stocks from which several stems sprout (mainly > 10). Within each selected plot, stem discs were sampled from shoots belonging to different stocks. Stem discs were taken as close to the stem base as possible. Five to ten stems were sampled in each plot. In order to cover a large range of environmental conditions, eight plots were selected within the treeline ecotone in relation to different geomorphic features (Table 1). The selected plots are located close to each other within an area of c. 0.5 km2, having a size between 100 and 700 m2, except for the plot at the highest elevation (plot 5), which is composed of several scattered bushes covering 5–10 m2 each.

Table 1.
Site description and characteristics of selected Alnus alnobetula stands with elevation (m above sea level (asl)), slope (S, °), soil depth (SoD, cm), stem length (StL, m), canopy height (CH, m), age (years) mean sensitivity (MS, %), ring width (RW, µm) and sample depth (n radii/stems). Mean values ± standard deviation (SD) are shown for stem length, age and ring width.
Due to prostrate growth form, wedging rings and eccentricity of the pith are characteristics of RG of A. alnobetula. Serial sectioning, i.e., sampling of discs at multiple points along the stem, was conducted on a sub-selection of three stems per plot to check for possibly missing rings [25,42]. Final averaging of several radii along the stem was found to be useful for correct dating and the construction of time series of RG [43,44]. Stem discs were marked and harvested at regular intervals (every 20 cm) along the shoot. Depending on shoot length, between 5 and 15 stem discs were sampled per stem. Canopy height and stem length were determined by measuring the vertical distance of the canopy (i.e., branch tips) to the ground and the length from the base of the stem to the tip, respectively.
Additionally, increment cores were sampled from co-occurring Pinus cembra individuals (n = 25 trees; two radii per tree taken perpendicular to the slope to avoid reaction wood). Trees with a height of 2–4 m, which do not show severe damage due to, e.g., wind breakage, were sampled between 2050 and 2150 m asl on south to east facing slopes. Stem discs and increment cores were air dried and the surface was prepared with a sharp razor blade. For contrast enhancement of ring boundaries, the surface was treated with a diluted wood stain or chalk for samples of A. alnobetula and P. cembra, respectively. Ring widths were measured to the nearest 1µm (two radii) using a light microscope (Olympus SZ61, Tokyo, Japan) fitted with a LINTAB measuring system (Frank Rinn, Heidelberg, Germany).
Inherent age/size trends in ring width measurements of both species were determined by averaging cambial-age-aligned ring width series (e.g., [45]). Trends in these ring width series, expressed as a cubic function, were used to detrend raw measurements for “biological-noise” (i.e., age/size trend) by calculating residuals (i.e., ring width series minus trend function). This procedure makes it possible to capture the long-term growth response to climate warming. Correct dating of measured ring width time series was statistically checked with COFECHA [46]. The partially low agreement in ring width variations among radii within and between A. alnobetula stems, which was caused by frequent occurrence of asymmetric growth, anomalous growth patterns and high portions of discontinuous rings (Supplementary Material, Figure S1), made it necessary to exclude about a quarter of the measured radii when creating the composite A. alnobetula chronology. Finally, the developed composite ring width chronologies comprised 86 radii taken from 51 stems of A. alnobetula and 24 radii from 16 stems of P. cembra. We refrained from analysing site-specific time series of radial stem growth because low sample depth and different series length would have made a comparative analysis impossible.
2.3. Climate–Growth Relationships
The relationships between climate and growth were determined by elaborating “response functions”. This is a form of principal component regression used to account for the collinearity of monthly climate predictors [47]. Total monthly precipitation and mean maximum, minimum and mean monthly air temperatures, which were collected on Mt. Patscherkofel (2246 m asl), were used in response function analyses by applying the software package Dendroclim2002 [48]. The meteorological station was <500 m in linear distance from the selected plots. Prior to calculating response functions, the ring width time series were standardized by the ARSTAN program [49]. To remove differences in growth rate of individual stems and the low-frequency variability that is assumed to be unrelated to climate (age/size trend, stand development), we applied a conservative detrending method, i.e., a linear regression of negative slope or a negative exponential decline was fitted to each ring series. Dimensionless radial growth indices were formed by dividing the observed ring width value by the predicted ring width value. Climatic variables used in the response function analysis included climate variables for individual months June–December of the previous year’s growth and January–September of the current year’s growth. The statistical relationship between ring-width and each monthly climatic variable was examined over the period 1991–2020 (n = 30 yrs) at the 95% confidence level.
Extreme growth years in the standardized ring width chronology of A. alnobetula were determined according to [50] by calculating the difference between the current year’s growth and the average growth of the last five years and subsequent division by the standard deviation for the period 1991–2020. Years showing growth deviation >±1 SD were related to climate factors that were found to significantly limit RG.
3. Results
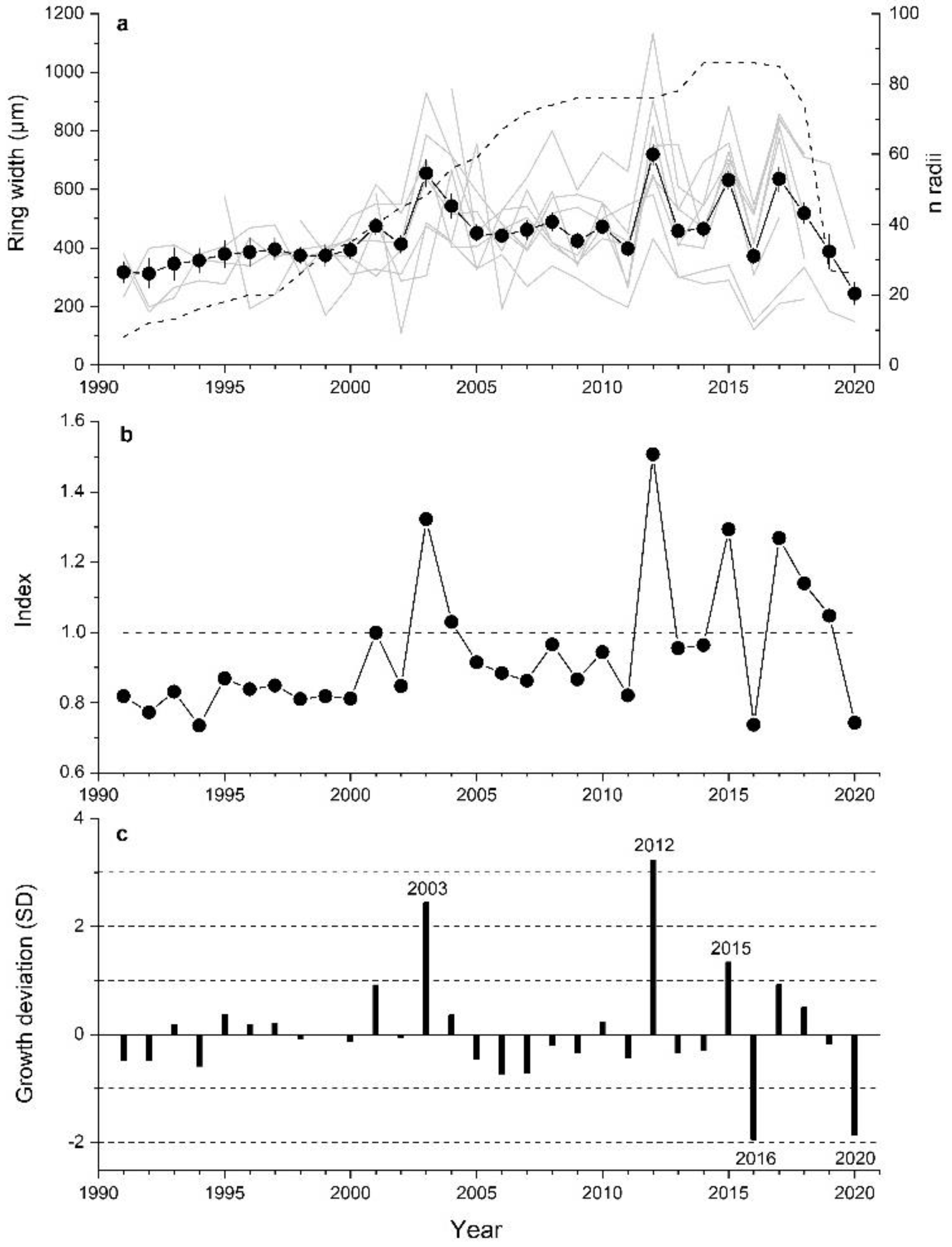
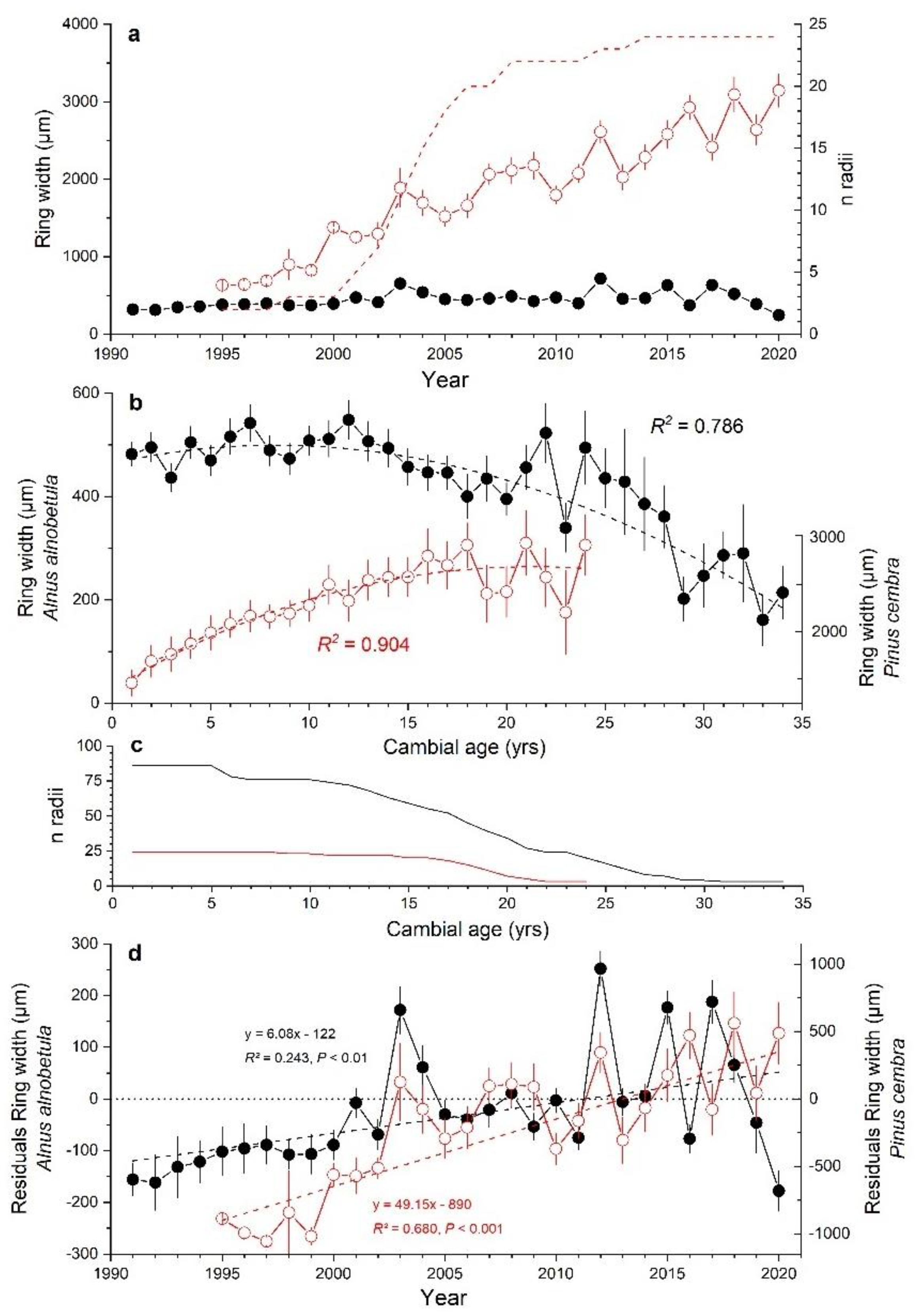
Selected A. alnobetula stands were located at elevations between 2050 to 2190 m asl, on slopes ranging from 20 to 35° and facing all aspects (Table 1). Soil depth varied from very shallow (c. 5 cm) to a maximum of 20 cm. Stems had mean age of 18 ± 7 yrs (mean value ± standard deviation) and the oldest stem had 47 annual rings. Canopy height varied between 0.6 and 3.5 m and mean length of sampled stems amounted to 2.3 ± 0.12 m. Mean ring width of all stands amounted to 519 ± 92 µm with a mean sensitivity of 37% (Table 1). After cross-dating, 86 radii from 51 stems were used to establish the composite chronology (Figure 2a). Individual site chronologies show a low degree of agreement in year-to-year variations of ring widths at low cambial age, and in the most recent decade a decreasing trend in radial growth is obvious at two plots having shallow soil depth (plots 2 and 8). In Table S1 (Supplementary Material), the agreement of year-to-year changes in ring width series among all study plots is quantified using percentage of sign agreement W [51] and the correlation coefficient t [52]. We obtained a 30-year-long ring width chronology of A. alnobetula with a sample depth of at least 8 radii. In this composite, chronology periods of predominantly constant growth in the 1990s and between 2004 and 2011 alternate with years showing strong fluctuations in RG. Distinct deviations from mean growth occurred in 2003 and in the most recent decade (Figure 2b,c).
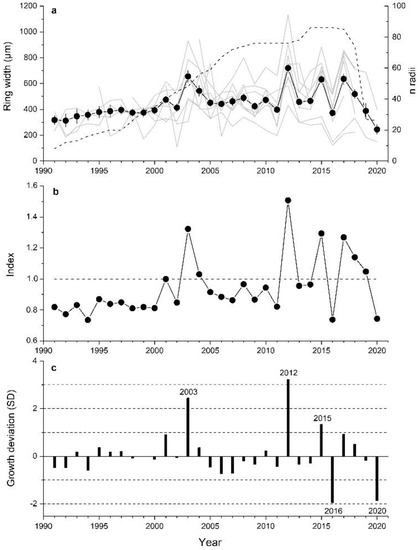
Figure 2.
(a) Composite ring width time series (±standard error; closed symbols and bold line) and site chronologies (grey lines) of Alnus alnobetula stems from 8 study plots (see Table 1). Number of stems included in the composite chronology (n > 5) is shown as dashed line. (b) Standardized chronology used as predictand in response function analysis. (c) Current year growth in relation to average value of previous 5 years given in standard deviation units (SD). Extreme growth years showing deviation >±1 SD are indicated (for details see Materials and Methods).
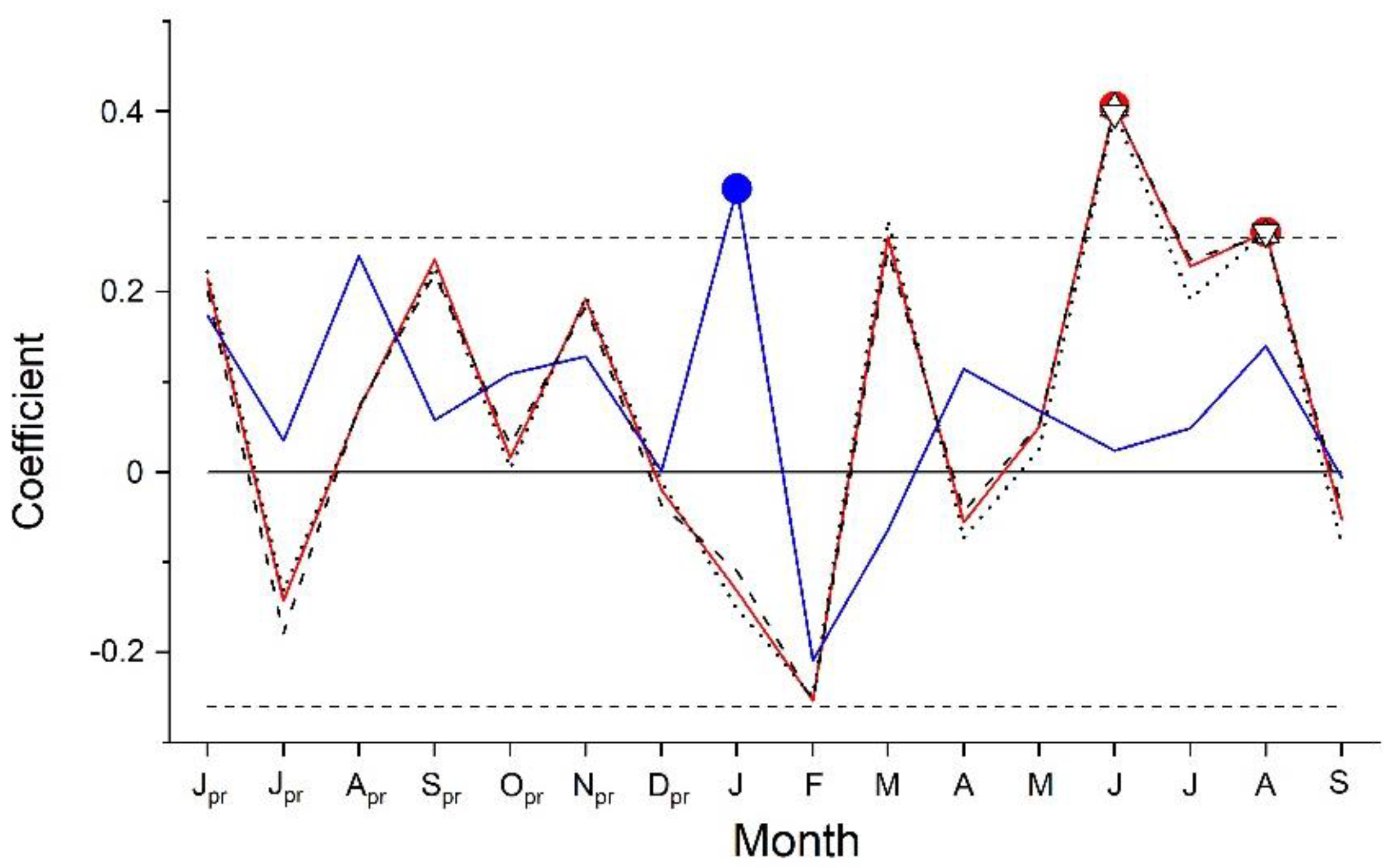
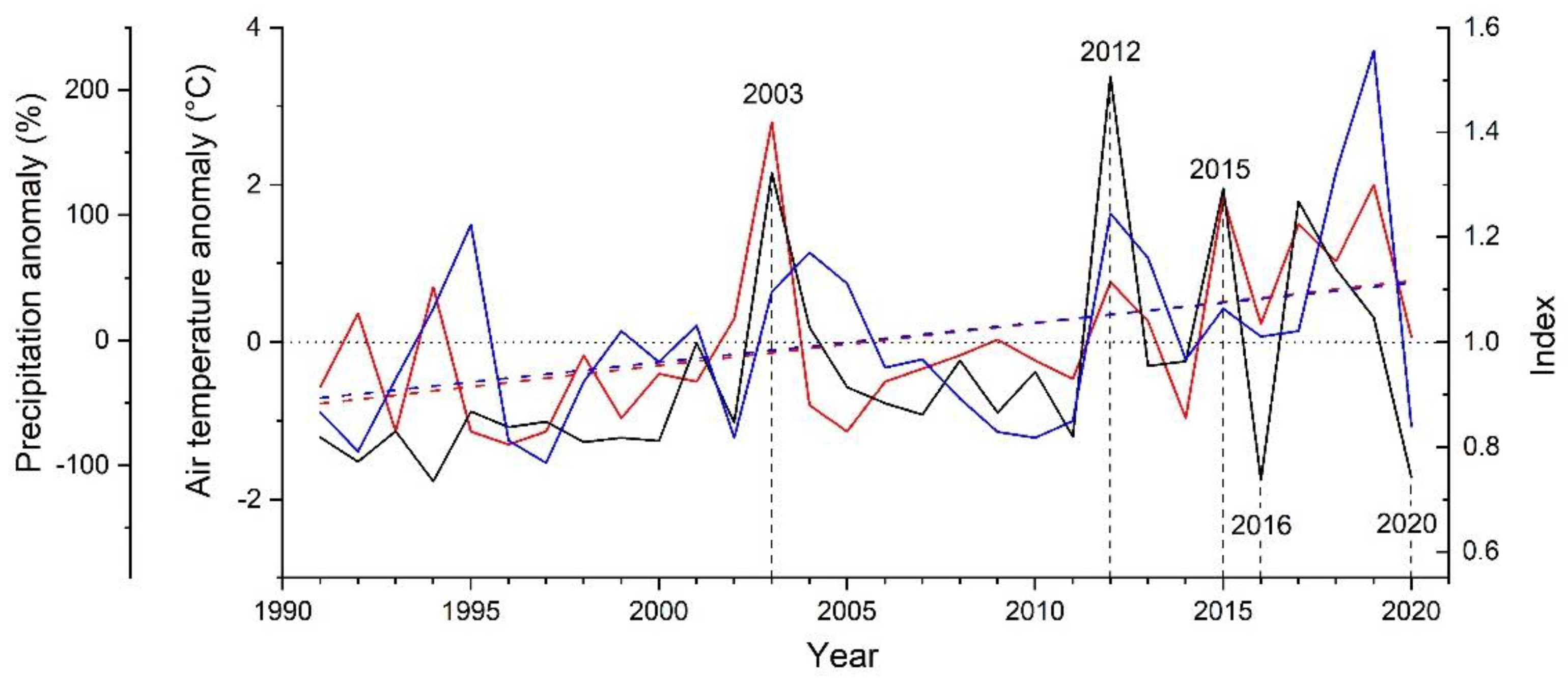
Results of response function analyses are depicted in Figure 3. Climate factors significantly (p < 0.05) controlling RG of A. alnobetula were air temperature in June and August and precipitation in January. Anomalies in summer (June through August) air temperature were more consistent with extreme growth deviations than anomalies in January precipitation (Figure 4, Supplementary Material Table S2). Distinct positive deviations in RG in 2003, 2012 and 2015 are in line with above LTM summer temperature and January precipitation. However, in years when distinct negative growth deviations occurred, these climate variables decreased compared to the previous year but were similar to LTM in 2016. In 2020, January precipitation was strongly reduced (−69%) compared to LTM (Supplementary Material, Table S2). During the period 1991–2020, mean summer temperature und January precipitation show an increase by c. 1.5 °C and c. 100 mm, respectively.
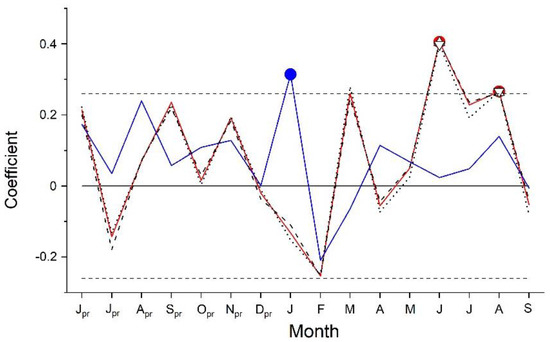
Figure 3.
Response function analysis between standardized composite chronology of Alnus alnobetula and monthly precipitation sum (blue line and circle) and monthly mean, maximum and minimum air temperature (red line and filled circle, dashed black line and upward triangle, dotted line and downward triangle, respectively). Horizontal dashed lines represent 95% confidence intervals. Symbols indicate significant relationships at p < 0.05 (pr = previous year climate variable).
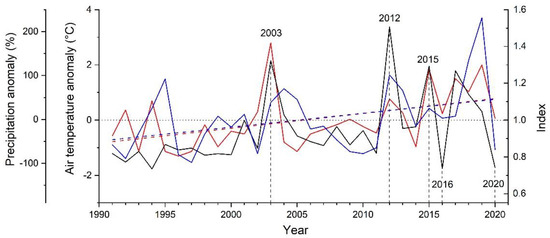
Figure 4.
Comparison of standardized radial stem growth (black line) of Alnus alnobetula with anomalies in summer mean monthly air temperature (June–August; red line) and January precipitation (blue line) from long-term mean 1991 to 2020. Long-term means of air temperature during summer and January precipitation amounted to 8.3 °C and 45 mm, respectively. Trend lines (linear regressions) for summer temperature (R2 = 0.212) and January precipitation (R2 = 0.138) are indicated by dashed lines. Extreme growth years are labelled and marked with vertical dashed lines.
Mean age of P. cembra trees amounted to 18 ± 4 yrs (mean ± standard deviation) and was not significantly different from mean age of A. alnobetula stems (18 ± 7 yrs). However, a comparison of RG during the period 1995–2020 revealed striking differences (Figure 5a–d): (i) mean ring width of P. cembra (2311 ± 628 µm; mean sensitivity 24%) was more than four-times larger than that of A. alnobetula (Figure 2a and Figure 5a), (ii) P. cembra showed markedly increasing growth rates with age/size while A. alnobetula stems showed only slightly increasing growth during the first decade which steadily declined thereafter (Figure 5b), (iii) the long-term trend in ring widths after eliminating age/size trend was more pronounced in P. cembra than in A. alnobetula (Figure 5d), and (iv) up to 2015 there was some synchrony in the ring width time series between A. alnobetula and P. cembra which is lost thereafter (Figure 5d). The different growth response to environmental conditions prevailing in 2016 and continuously decreasing RG in A. alnobetula since 2017 stand out in particular.
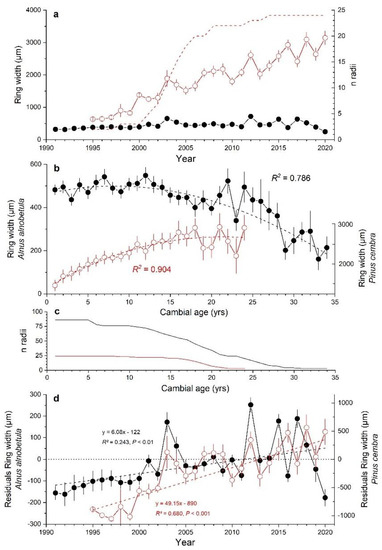
Figure 5.
(a) Ring width time series of Pinus cembra (red line and open circles; dashed line indicates sample depth) and Alnus alnobetula (black line and filled circles), (b) ring widths aligned by cambial age for Alnus alnobetula (left y-axis; black lines and filled circles) and P. cembra (right y-axis; red lines and open circles); age/size trends in ring width are expressed as cubic functions and R2-values are indicated, (c) sample depth for age/size trend analysis and (d) residuals taking age/size trends into account (for details see Materials and Methods). Equations for the linear trend fitted, the squared correlation coefficient (R2) and the significance of the regression slope (p-value) are indicated. Time series of P. cembra and A. alnobetula are shown by red lines and open circles and black lines and filled circles, respectively. Mean values ± standard error are shown.
Although A. alnobetula shows strikingly lower RG compared to co-occurring P. cembra, aerial photographs of the study area dating back to 1974 show that A. alnobetula stands are spreading rapidly within the treeline ecotone (Figure 6), amounting to c. 450 m2 ha−1 decade−1 at some sites (F Bernich, unpublished data). Spread of A. alnobetula within the treeline ecotone as seen from the ground is shown in Figure S2 (Supplementary Material).

Figure 6.
Example plot showing the spread of green alder (Alnus alnobetula) within the treeline ecotone on Mt. Patscherkofel. Orthophotos were taken in 1974 (a) and 2019 (b); elevation range: c. 2100–2200 m asl. The marked rectancle includes stands that were sampled in this study. Source: Orthophoto Land Tirol, tiris (https://lba.tirol.gv.at/public/karte.xhtml; 24 February 2022, and https://maps.tirol.gv.at/; 24 February 2022).
4. Discussion
4.1. Age Structure and Growth Characteristics of Alnus alnobetula
Due to frequent re-sprouting from the rootstock and formation of adventitious shoots, the age of a particular stem can be determined by counting the number of tree rings but not the precise age of the rootstock. Nevertheless, the different age structure of selected stands indicates that A. alnobetula is spreading at the treeline ecotone on Mt. Patscherkofel into habitats that are outside their primary ecological habitat requirements, i.e., north-facing slopes with a long snow cover or moist sites with high geomorphic activity [9,10,53,54]. This view is corroborated by evaluation of times series of aerial photographs of the study area which revealed that A. alnobetula stands are rapidly expanding on south-east facing slopes. That A. alnobetula is capable of developing stands on shallow, drought-prone south to south-east facing slopes is in accordance with findings in several other studies (see [12] and references therein). The small-scale fragmentation of stands indicates that the development of A. alnobetula depends on microsite conditions, i.e., adequate water availability during the growing season and/or a sufficient depth of snow as a protection against frost drought. Although these environmental factors might be a prerequisite for successful establishment, low synchronicity in annual increments among individual stands, which was predominantly found at low cambial age, most likely is related to site-specific differences in mechanical strain of young stems due to varying extent of snow load and/or snow creep.
4.2. Climate Forcing of Radial Stem Growth of Alnus alnobetula
Although the geomorphic characteristics of the selected stands differed considerably, ring width series of 86 radii from 8 plots could be assembled into a composite chronology, suggesting that common climatic drivers control RG of A. alnobetula in the study area. In our composite chronology RG varied greatly from year-to-year (mean sensitivity amounted to 37%) and response function analysis revealed that this variability was associated with air temperature during summer, confirming our first hypothesis. That shrub growth in cold environments is constrained by low temperatures during summer was reported by several authors (e.g., [33,44,55,56,57,58]). This is in accordance with the growth limitation hypothesis of treeline formation put forward by [21,59], which states that low temperatures during the growing season (global mean: 6.4 ± 0.7 °C) restrict meristematic activity and tissue renewal at high-elevation treelines around the globe [60].
Our second hypothesis, that precipitation at the start of the growing season favours RG by supporting cambial activity and expansion growth (cell enlargement), could not be confirmed. Our findings suggest that the root system of A. alnobetula is capable of providing adequate water supply during bud-break, leaf unfolding and onset of secondary growth, i.e., earlywood formation. As the soil water content is generally above field capacity at the start of the growing season [61,62], differences in January snow depth hardly affect water availability for A. alnobetula at the time of bud-break and onset of RG. Ample summer precipitation ensures sufficient water availability during the growing season. Unexpectedly, snow cover during winter (i.e., precipitation in January) positively affected RG. It is well known that at high elevation in the temperate climate zone, snow cover during winter is a crucial factor for tree growth, preventing adverse effects of winter desiccation (frost drought). Frost drought is a frequent phenomenon affecting growth of tree species at the treeline within the study area [36,37], where it is favoured by a precipitation minimum in winter (Figure 1). That the distribution of A. alnobetula is affected by snow cover (depth and duration) was already suggested by [10,11]. Occurrence of frost drought in A. alnobetula is also supported by an experimental study carried out by Frey [63], who found that seedlings suffer from frost drought if snow protection was not provided. These findings suggest that high snow depth is crucial to protect overwintering buds from frost drought. Ref. [31] also suggested that reduction of snow cover could predispose alder stems to frost drought, reducing their vitality and inducing decline of stands with the participation of biotic agents (fungi, insects). Because A. alnobetula has symbiotic association with nitrogen-fixing bacteria, it is highly unlikely that the direct relationship found between January precipitation (as a proxy of snow depth) and RG is related to an increase in nutrient supply caused by increase in microbial activity under higher snow depth [64,65].
Climate in extreme growth years mostly confirmed results of response function analysis, i.e., in years showing distinct positive deviations in RG (2003, 2012 and 2015) above average temperature and precipitation prevailed during summer and January, respectively. In 2016, when RG showed a striking decrease, climate variables found to constrain RG were lower compared to the previous year but corresponded to LTM. However, it should be noted that alternating warm and cold phases may cancel each other out and are therefore not reflected in monthly averages. Furthermore, a late frost event in late April through early May 2016 caused daily mean temperatures to drop below −10 °C. Although A. alnobetula does survive freezing temperatures down to −45 °C in the frost-hardened state during winter [66], frost resistance was found to strongly decrease during the development of the current year’s shoot growth, i.e., during bud-break and expansion growth temperatures <−8 °C cause extensive frost damage (e.g., [67]). Because leaf primordia and young leaves are a main source of the hormone auxin [68], which is a key factor in the control of cambial cell division [69], damage to young shoots by this late frost event may have indirectly affected RG. Growth decrease in 2020 is most likely related to effects of frost drought (decrease in January precipitation by −69% compared to LTM) and/or below average temperature in June (−0.8 °C compared to LTM).
4.3. Long Term Trend in Radial Stem Growth of Co-Occurring Alnus alnobetula and Pinus cembra
Climate records show an increase in summer temperature during the study period 1991–2020 amounting to c. 1.5 °C and also an increase in January precipitation by about 40 mm. Because both climate variables are directly related to RG of A. alnobetula and P. cembra [36,70], an increase in annual increments in recent decades was expected. Time series of annual increments are inherently influenced by ontogeny of the stem (age/size), i.e., ring widths in trees and shrubs are increasing in first decades and subsequently decreasing with increasing age/size (e.g., [71,72]). After correcting for the age/size trend in ring width time series, A. alnobetula and P. cembra show an increase in RG during the study period, which is more pronounced in the tree species P. cembra. Several authors [34,73,74] also reported diverging growth trends to climate warming among coexisting shrubs and tree species in cold environments (alpine and polar biomes). Authors suggested that growth of trees is more closely coupled to rising temperature than growth of shrubs, indicating marked influence of ground microclimate or indirect effects, e.g., reduced period of snow cover, on shrub growth. Although microclimatic effects may also play a role in the minor response of A. alnobetula to climate warming, differences in growth rates (four-fold larger annual increments in P. cembra compared to A. alnobetula), in age/size and long-term growth trends of single stemmed P. cembra and multi-stemmed A. alnobetula may be due to different carbon allocation strategies, i.e., preference of aboveground stem growth in late successional tree species P. cembra vs. clonal growth in the pioneer shrub A. alnobetula. Ref. [75] already stated that the development of adventive shoots and root suckers are key parts of the “horizontal competition strategy” of A. alnobetula. In contrast to other treeline conifers, e.g., Larix decidua or Picea abies, P. cembra does not have the ability to clonally propagate [53].
We are aware that the age/size trend used to correct the ring width chronologies of both species for non-climatic trends is based on a limited number of samples [76,77]. Although a cautious interpretation of the long-term growth trend is therefore required, our trend analysis for P. cembra is supported by Oberhuber et al. [78], who reported that several treeline conifers in the Eastern Central Alps (including P. cembra) respond to climate warming with a comparable increase in radial stem growth. Whether the declining growth in A. alnobetula in recent years is the start of a general growth decline as found by Pisetta et al. [31] in the Trentino mountains (Italy) requires further monitoring in combination with detailed physiological studies (e.g., intra-annual records of radial growth, sapflow, leaf water potential and soil moisture availability) to be able to clarify the underlying causes.
5. Conclusions
As expected, this study revealed predominant influence of air temperature during summer on RG of A. alnobetula. Additionally, results suggest that frost drought effects caused by low precipitation in winter and late frost events constrain radial growth. Hence, if other abiotic and biotic factors are disregarded, ongoing climate warming will promote further spread of A. alnobetula across the alpine treeline ecotone as long as (i) adequate winter precipitation minimizes the effects of frost drought and (ii) the occurrence of shoot damage due to late frost events in a warming induced lengthened growing season (e.g., [79]) is not a frequently recurring phenomenon.
The comparison of composite RG chronologies among co-occurring A. alnobetula and P. cembra at the treeline ecotone on Mt. Patscherkofel let us conclude that two widespread species at the alpine treeline ecotone of the Central European Alps, which belong to distinct functional groups (deciduous shrub having prostrate growth form vs. evergreen tree with arborescent growth), strongly differ with respect to RG at the stem base. In multi-stemmed A. alnobetula clonal propagation, i.e., spatial extension by development of adventitious shoots and root suckers might be favoured in contrast to what is seen in single-stemmed P. cembra. By favouring clonal propagation over individual stem growth, A. alnobetula is able to quickly spread at the alpine treeline ecotone, which was frequently reported to occur in the European Alps (e.g., [12]) and elsewhere (e.g., [7,80]). Because a reduction in grazing pressure facilitates shrub encroachment [81,82,83], changes in ecosystem properties, e.g., decrease in biodiversity, increase in nitrate leaching and suppression of forest succession [4,19], can be expected at the alpine treeline ecotone with ongoing land abandonment and global change.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f13030440/s1, Figure S1: Cross-sections of Alnus alnobetula wood samples. Figure S2: Spread of A. alnobetula within the treeline ecotone on Mt. Patscherkofel. Table S1: Agreement of year-to-year changes in ring width series of A. alnobetula among study plots. Table S2: Anomalies in January precipitation and summer air temperature in extreme growth years of A. alnobetula.
Author Contributions
W.O., G.W. and A.G. conceived the study. W.O. co-ordinated the overall project. F.B., A.G. and W.O. collected, prepared and analysed the data. W.O., G.W., A.G. and F.B. interpreted and discussed the data. W.O. wrote the manuscript and G.W. and A.G. provided editorial advice. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Austrian Science Fund (FWF), P34706-B. For the purpose of open access, the author has applied a CC BY public copyright licence to any Author Accepted Manuscript version arising from this submission.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The ring width data presented in this study are openly available in Zenodo at 10.5281/zenodo.5931440.
Acknowledgments
We thank Ethan Salino and Natalie Wallner for their help with field work and radial growth measurements. Open Access Funding by the Austrian Science Fund (FWF).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wiedmer, E.; Senn-Irlet, B. Biomass and primary productivity of an Alnus viridis stand—A case study from the Schächental valley, Switzerland. Bot. Helv. 2006, 116, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Gehrig-Fasel, J.; Guisan, A.; Zimmermann, N.E. Tree line shifts in the Swiss Alps: Climate change or land abandonment? J. Veg. Sci. 2007, 18, 571–582. [Google Scholar] [CrossRef]
- Boscutti, F.; Poldini, L.; Buccheri, M. Green alder communities in the Alps: Phytosociological variability and ecological features. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2013, 148, 917–934. [Google Scholar] [CrossRef]
- Bühlmann, T.; Hiltbrunner, E.; Körner, C. Alnus viridis expansion contributes to excess reactive nitrogen release, reduces biodiversity and constrains forest succession in the Alps. Alp. Bot. 2014, 124, 187–191. [Google Scholar] [CrossRef] [Green Version]
- Anthelme, F.; Michalet, R.; Barbaro, L.; Brun, J.-J. Environmental and Spatial Influences of Shrub Cover (Alnus viridis DC.) on Vegetation Diversity at the Upper Treeline in the Inner Western Alps. Arct. Antarct. Alp. Res. 2003, 35, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Beck, P.S.A.; Horning, N.; Goetz, S.J.; Loranty, M.M.; Tape, K.D. Shrub Cover on the North Slope of Alaska: A circa 2000 Baseline Map. Arct. Antarct. Alp. Res. 2011, 43, 355–363. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Forbes, B.C.; Wilmking, M.; Hallinger, M.; Lantz, T.; Blok, D.; Tape, K.D.; Macias-Fauria, M.; Sass-Klaassen, U.; Lévesque, E.; et al. Shrub expansion in tundra ecosystems: Dynamics, impacts and research priorities. Environ. Res. Lett. 2011, 6, 045509. [Google Scholar] [CrossRef] [Green Version]
- Frost, G.V.; Epstein, H.E. Tall shrub and tree expansion in Siberian tundra ecotones since the 1960s. Glob. Chang. Biol. 2014, 20, 1264–1277. [Google Scholar] [CrossRef]
- Schröter, C. Das Pflanzenleben der Alpen: Eine Schilderung der Hochgebirgsflora; Albert Raustein: Zürich, Switzerland, 1908; p. 806. [Google Scholar]
- Richard, L. Ecologie de l’Aulne Vert (Alnus viridis Chaix): Facteurs climatiques et édaphiques. Doc. Carte. Veg. Alpes. 1968, 6, 107–158. [Google Scholar]
- Richard, L. Une interpretation eco-physiologique de la repartition de l’aune vert (Alnus viridis). Doc. Carte. Veg. Alpes. 1969, 7, 7–23. [Google Scholar]
- Caviezel, C.; Hunziker, M.; Kuhn, N.J. Green alder encroachment in the European Alps: The need for analyzing the spread of a native-invasive species across spatial data. Catena 2017, 159, 149–158. [Google Scholar] [CrossRef]
- Schwencke, J.; Carú, M. Advances in Actinorhizal Symbiosis: Host Plant- Frankia Interactions, Biology, and Applications in Arid Land Reclamation. A Review. Arid Land Res. Manag. 2001, 15, 285–327. [Google Scholar] [CrossRef]
- Dawson, J. Ecology of actinorhizal plants. In Nitrogen-Fixing Actinorhizal Symbioses; Pawlowski, K., Newton, W., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 199–234. [Google Scholar]
- Chatarpaul, L.; Chakravarty, P.; Subramaniam, P. Studies in tetrapartite symbioses. Plant Soil 1989, 118, 145–150. [Google Scholar] [CrossRef]
- Salmon, V.G.; Breen, A.L.; Kumar, J.; Lara, M.; Thornton, P.E.; Wullschleger, S.D.; Iversen, C.M. Alder Distribution and Expansion Across a Tundra Hillslope: Implications for Local N Cycling. Front. Plant Sci. 2019, 10, 1099. [Google Scholar] [CrossRef] [PubMed]
- Bühlmann, T.; Körner, C.; Hiltbrunner, E. Shrub Expansion of Alnus viridis Drives Former Montane Grassland into Nitrogen Saturation. Ecosystems 2016, 19, 968–985. [Google Scholar] [CrossRef]
- Rubli, D. Waldbauliche Untersuchungen in Grünerlenbeständen. Ph.D. Thesis, ETH Zurich, Zurich, Switzerland, 1974. [Google Scholar]
- Hiltbrunner, E.; Aerts, R.; Bühlmann, T.; Huss-Danell, K.; Magnusson, B.; Myrold, D.; Reed, S.; Sigurdsson, B.D.; Körner, C. Ecological consequences of the expansion of N2-fixing plants in cold biomes. Oecologia 2014, 176, 11–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Körner, C. Alpine Treelines. In Functional Ecology of the Global High Elevation Tree Limits; Springer: Basel, Switzerland, 2012; p. 220. [Google Scholar]
- Wieser, G.; Oberhuber, W.; Gruber, A. Effects of Climate Change at Treeline: Lessons from Space-for-Time Studies, Manipulative Experiments, and Long-Term Observational Records in the Central Austrian Alps. Forest 2019, 10, 508. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, J.S.; Ruess, R.W. Seasonal patterns of climate controls over nitrogen fixation by Alnus viridis subsp. fruticosa in a secondary successional chronosequence in interior Alaska. Ecoscience 2009, 16, 341–351. [Google Scholar] [CrossRef]
- Lantz, T.C.; Gergel, S.E.; Henry, G.H.R. Response of green alder (Alnus viridis subsp. Fruticosa) patch dynamics and plant community composition to fire and regional temperature in north-western Canada. J. Biogeogr. 2010, 37, 1597–1610. [Google Scholar]
- Myers-Smith, I.H.; Hallinger, M.; Blok, D.; Sass-Klaassen, U.; Rayback, S.A.; Weijers, S.; Trant, A.J.; Tape, K.D.; Naito, A.; Wipf, S.; et al. Methods for measuring arctic and alpine shrub growth: A review. Earth Sci. Rev. 2015, 140, 1–13. [Google Scholar] [CrossRef]
- Körner, C.; Jussel, U.; Schiffer, K. Transpiration, Diffusionswiderstand und Wasserpotential in verschiedenen Schichten eines Grünerlenbestandes. In Ökologische Analysen von Almflächen im Gasteiner Tal. Veröff Österr MaB-Hochgebirgsprogramm Hohe Tauern Band 2; Cernusca, A., Ed.; Universitätsverlag Wagner: Innsbruck, Austria, 1978; pp. 81–98. [Google Scholar]
- Herbst, M.; Eschenbach, C.; Kappen, L. Water use in neighbouring stands of beech (Fagus sylvatica L.) and black alder (Alnus glutinosa (L.) Gaertn.). Ann. Des. Sci. For. 1999, 56, 107–120. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Bigler, C.; Bugmann, H. Growth-dependent tree mortality models based on tree rings. Can. J. For. Res. 2003, 33, 210–221. [Google Scholar] [CrossRef]
- Dobbertin, M. Tree growth as indicator of tree vitality and of tree reaction to environmental stress: A review. Forstwiss. Cent. 2005, 124, 319–333. [Google Scholar] [CrossRef]
- Pisetta, M.; Montecchio, L.; Longa, C.M.O.; Salvadori, C.; Zottele, F.; Maresi, G. Green alder decline in the Italian Alps. For. Ecol. Manag. 2012, 281, 75–83. [Google Scholar] [CrossRef]
- Liang, E.; Lu, X.; Ren, P.; Li, X.; Zhu, L.; Eckstein, D. Annual increments of juniper dwarf shrubs above the tree line on the central Tibetan Plateau: A useful climatic proxy. Ann. Bot. 2012, 109, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, R.H.; Hallinger, M.; Ahlgrimm, S.; Friemel, J.; Kollmann, J.; Meilby, H. Growth response to climatic change over 120 years for Alnus viridis and Salix glauca in West Greenland. J. Veg. Sci. 2015, 26, 155–165. [Google Scholar] [CrossRef]
- Dearborn, K.D.; Danby, R.K. Topographic influences on ring widths of trees and shrubs across alpine treelines in southwest Yukon. Arctic. Antarct. Alp. Res. 2018, 50, 1495445. [Google Scholar] [CrossRef] [Green Version]
- Weijers, S.; Beckers, N.; Löffler, J. Recent spring warming limits near-treeline deciduous and evergreen alpine dwarf shrub growth. Ecosphere 2018, 9, 02328. [Google Scholar] [CrossRef]
- Oberhuber, W. Influence of climate on radial growth of Pinus cembra within the alpine timberline ecotone. Tree Physiol. 2004, 24, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Tranquillini, W. Physiological Ecology of the Alpine Timberline: Tree Existence in High Altitudes with Special Reference to the European Alps; Springer: Berlin/Heidelberg, Germany, 1979; Volume 31, p. 137. [Google Scholar]
- Wieser, G. Lessons from the timberline ecotone in the Central Tyrolean Alps: A review. Plant Ecol. Divers. 2012, 5, 127–139. [Google Scholar] [CrossRef]
- Tollmann, A. Geologie von Österreich Band 1. Die Zentralalpen; Deuticke: Wien, Austria, 1977; p. 766. [Google Scholar]
- Neuwinger, I. Böden der subalpinen und alpinen Stufe in den Tiroler Alpen. Mitt. Ostalpin Din. Ges. Veg. 1970, 11, 135–150. [Google Scholar]
- FAO; ISRIC; ISSS. World Reference Base for Soil Resources; FAO: Rome, Italy, 1998; p. 109. [Google Scholar]
- Kolishchuk, V.G. Dendroclimatological study of prostrate woody plants. In Methods of Dendrochronology: Applications in the Environmental Sciences; Cook, E.R., Kairiukstis, L.A., Eds.; Kluwer: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 1990; pp. 51–55. [Google Scholar]
- Bär, A.; Bräuning, A.; Löffler, J. Ring-Width Chronologies of the Alpine Dwarf Shrub Empetrum Hermaphroditum from the Norwegian Mountains. IAWA J. 2007, 28, 325–338. [Google Scholar] [CrossRef]
- Hallinger, M.; Manthey, M.; Wilmking, M. Establishing a missing link: Warm summers and winter snow cover promote shrub expansion into alpine tundra in Scandinavia. New Phytol. 2010, 186, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Esper, J.; Cook, E.R.; Krusic, P.J.; Peters, K.; Schweingruber, F.H. Tests of the RCS Method for Preserving Low-Frequency Variability in Long Tree-Ring Chronologies. Tree-Ring Res. 2003, 59, 81–98. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Briffa, K.R.; Cook, E.R. Methods of response function analysis. In Methods of Dendrochronology; Cook, E.R., Kairiukstis, L.A., Eds.; Kluwer: Dordrecht, The Netherlands, 1990; pp. 165–178. [Google Scholar]
- Biondi, F.; Waikul, K. Dendroclim2002: A C++ program for statistical calibration of climate signals in tree-ring chronologies. Comput. Geosci. 2004, 30, 303–311. [Google Scholar] [CrossRef]
- Cook, E.R.; Holmes, R.L. Program ARSTAN User Manual; Laboratory of Tree Ring Research, University of Arizona: Tucson, AZ, USA, 1984; p. 14. [Google Scholar]
- Neuwirth, B.; Esper, J.; Schweingruber, F.H.; Winiger, M. Site ecological differences to the climatic forcing of spruce pointer years from the Lötschental, Switzerland. Dendrochronologia 2004, 21, 69–78. [Google Scholar] [CrossRef]
- Eckstein, D.; Bauch, J. Beitrag zur Rationalisierung eines dendrochronologischen Verfahrens und zur Analyse seiner Aussagesicherheit. Forstwiss. Cent. 1969, 88, 230–250. [Google Scholar] [CrossRef]
- Baillie, M.G.; Pilcher, J.R. A simple crossdating program for tree-ring research. Tree Ring Bull. 1973, 38, 35–43. [Google Scholar]
- Holtmeier, F.K. Mountain Timberlines; Ecology, Patchiness and Dynamics. Advances in Global Change Research; Springer: Dordrecht, The Netherlands, 2009; Volume 36, p. 437. [Google Scholar]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas mit den Alpen in Ökologischer, Dynamischer und Historischer Sicht; UTB: Stuttgart, Germany, 2010; p. 1357. [Google Scholar]
- Wettstein, S. Der Einfluss abiotischer Faktoren auf die Morphologie der Grünerle. Bot. Helv. 2001, 111, 31–44. [Google Scholar]
- Forbes, B.C.; Fauria, M.M.; Zetterberg, P. Russian Arctic warming and ‘greening’ are closely tracked by tundra shrub willows. Glob. Chang. Biol. 2010, 16, 1542–1554. [Google Scholar] [CrossRef]
- Rayback, S.A.; Henry, G.H.R. Reconstruction of Summer Temperature for a Canadian High Arctic Site from Retrospective Analysis of the Dwarf Shrub, Cassiope tetragona. Arct. Antarct. Alp. Res. 2006, 38, 228–238. [Google Scholar] [CrossRef] [Green Version]
- Buchwal, A.; Rachlewicz, G.; Fonti, P.; Cherubini, P.; Gärtner, H. Temperature modulates intra-plant growth of Salix polaris from a high Arctic site (Svalbard). Polar Biol. 2013, 36, 1305–1318. [Google Scholar] [CrossRef] [Green Version]
- Körner, C. Carbon limitation in trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, J.; Körner, C. A climate-based model to predict potential treeline position around the globe. Alp. Bot. 2014, 124, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Guggenberger, H. Untersuchungen zum Wasserhaushalt der Alpinen Zwergstrauchheide Patscherkofel. Ph.D. Thesis, University of Innsbruck, Innsbruck, Austria, 1980. [Google Scholar]
- Wieser, G.; Gruber, A.; Oberhuber, W. Sap flow characteristics and whole-tree water use of Pinus cembra across the treeline ecotone of the central Tyrolean Alps. Forstwiss. Cent. 2014, 133, 287–295. [Google Scholar] [CrossRef]
- Frey, W. The Influence of Snow on Growth and Survival of Planted Trees. Arct. Alp. Res. 1983, 15, 241. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bilbrough, C.; Welker, J.M. Increased snow depth affects microbial activity and nitrogen mineralization in two Arctic tundra communities. Soil Biol. Biochem. 2004, 36, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Larsen, K.S.; Grogan, P.; Jonasson, S.; Michelsen, A. Respiration and Microbial Dynamics in Two Subarctic Ecosystems during Winter and Spring Thaw: Effects of Increased Snow Depth. Arct. Antarct. Alp. Res. 2007, 39, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Neuner, G.; Kreische, B.; Kaplenig, D.; Monitzer, K.; Miller, R. Deep supercooling enabled by surface impregnation with lipophilic substances explains the survival of overwintering buds at extreme freezing. Plant Cell Environ. 2019, 42, 2065–2074. [Google Scholar] [CrossRef] [PubMed]
- Taschler, D.; Beikircher, B.; Neuner, G. Frost resistance and ice nucleation in leaves of five woody timberline species measured in situ during shoot expansion. Tree Physiol. 2004, 24, 331–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y. Auxin Biosynthesis and Its Role in Plant Development. Annu. Rev. Plant Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhalerao, R.; Fischer, U. Environmental and hormonal control of cambial stem cell dynamics. J. Exp. Bot. 2017, 68, 79–87. [Google Scholar] [CrossRef] [PubMed]
- OberhuberM, W.; Kofler, W.; Pfeifer, K.; Seeber, A.; Gruber, A.; Wieser, G. Long-term changes in tree-ring-climate relationships at Mt. Patscherkofel (Tyrol, Austria) since the mid-1980s. Trees 2008, 22, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bräker, O.U. Alterstrend bei Jahrringdichten und Jahrringbreiten von Nadelholzern und sein Ausgleich. Mitt. Vienna Forstl. Bundesver. 1981, 142, 75–102. [Google Scholar]
- Rixen, C.; Schwoerer, C.; Wipf, S. Winter climate change at different temporal scales in Vaccinium myrtillus, an Arctic and alpine dwarf shrub. Polar Res. 2010, 29, 85–94. [Google Scholar] [CrossRef]
- Pellizzari, E.; Camarero, J.J.; Gazol, A.; Granda, E.; Shetti, R.; Wilmking, M.; Moiseev, P.; Pividori, M.; Carrer, M. Diverging shrub and tree growth from the Polar to the Mediterranean biomes across the European continent. Glob. Chang. Biol. 2017, 23, 3169–3180. [Google Scholar] [CrossRef] [PubMed]
- Šenfeldr, M.; Kaczka, R.; Buras, A.; Samusevich, A.; Herrmann, C.; Spyt, B.; Menzel, A.; Treml, V. Diverging growth performance of co-occurring trees (Picea abies) and shrubs (Pinus mugo) at the treeline ecotone of Central European mountain ranges. Agric. For. Meteorol. 2021, 308–309, 108608. [Google Scholar] [CrossRef]
- Mallik, A.; Bell, F.; Gong, Y. Regeneration behavior of competing plants after clear cutting: Implications for vegetation management. For. Ecol. Manag. 1997, 95, 1–10. [Google Scholar] [CrossRef]
- Cook, E.R.; Briffa, K.; Meko, D.M.; Graybill, D.A.; Funkhouser, G. The ‘segment length curse’ in long tree-ring chronology development for palaeoclimatic studies. Holocene 1995, 5, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Peters, R.L.; Groenendijk, P.; Vlam, M.; Zuidema, P.A. Detecting long-term growth trends using tree rings: A critical evaluation of methods. Glob. Chang. Biol. 2015, 21, 2040–2054. [Google Scholar] [CrossRef] [PubMed]
- Oberhuber, W.; Bendler, U.; Gamper, V.; Geier, J.; Hölzl, A.; Kofler, W.; Krismer, H.; Waldboth, B.; Wieser, G. Growth Trends of Coniferous Species along Elevational Transects in the Central European Alps Indicate Decreasing Sensitivity to Climate Warming. Forest 2020, 11, 132. [Google Scholar] [CrossRef] [Green Version]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Tape, K.D.; Hallinger, M.; Welker, J.M.; Ruess, R.W. Landscape Heterogeneity of Shrub Expansion in Arctic Alaska. Ecosystems 2012, 15, 711–724. [Google Scholar] [CrossRef]
- Tasser, E.; Tappeiner, U. Impact of land use changes on mountain vegetation. Appl. Veg. Sci. 2002, 5, 173–184. [Google Scholar] [CrossRef]
- Anthelme, F.; Villaret, J.C.; Brun, J.J. Shrub encroachment in the Alps gives rise to the convergence of sub-alpine communities on a regional scale. J. Veg. Sci. 2007, 18, 355–362. [Google Scholar] [CrossRef]
- Jørgensen, R.H.; Meilby, H.; Kollmann, J. Shrub expansion in SW Greenland under modest regional warming: Disentangling effects of human disturbance and grazing. Arct. Antarct. Alp. Res. 2013, 45, 515–525. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).