
Coupled Relationship between Soil Physicochemical Properties and Plant Diversity in the Process of Vegetation Restoration


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Conditions
2.2. Site Selection and Plant Investigation
2.3. Soil Sample Collection and Determination
2.4. Statistical Analyses
3. Results
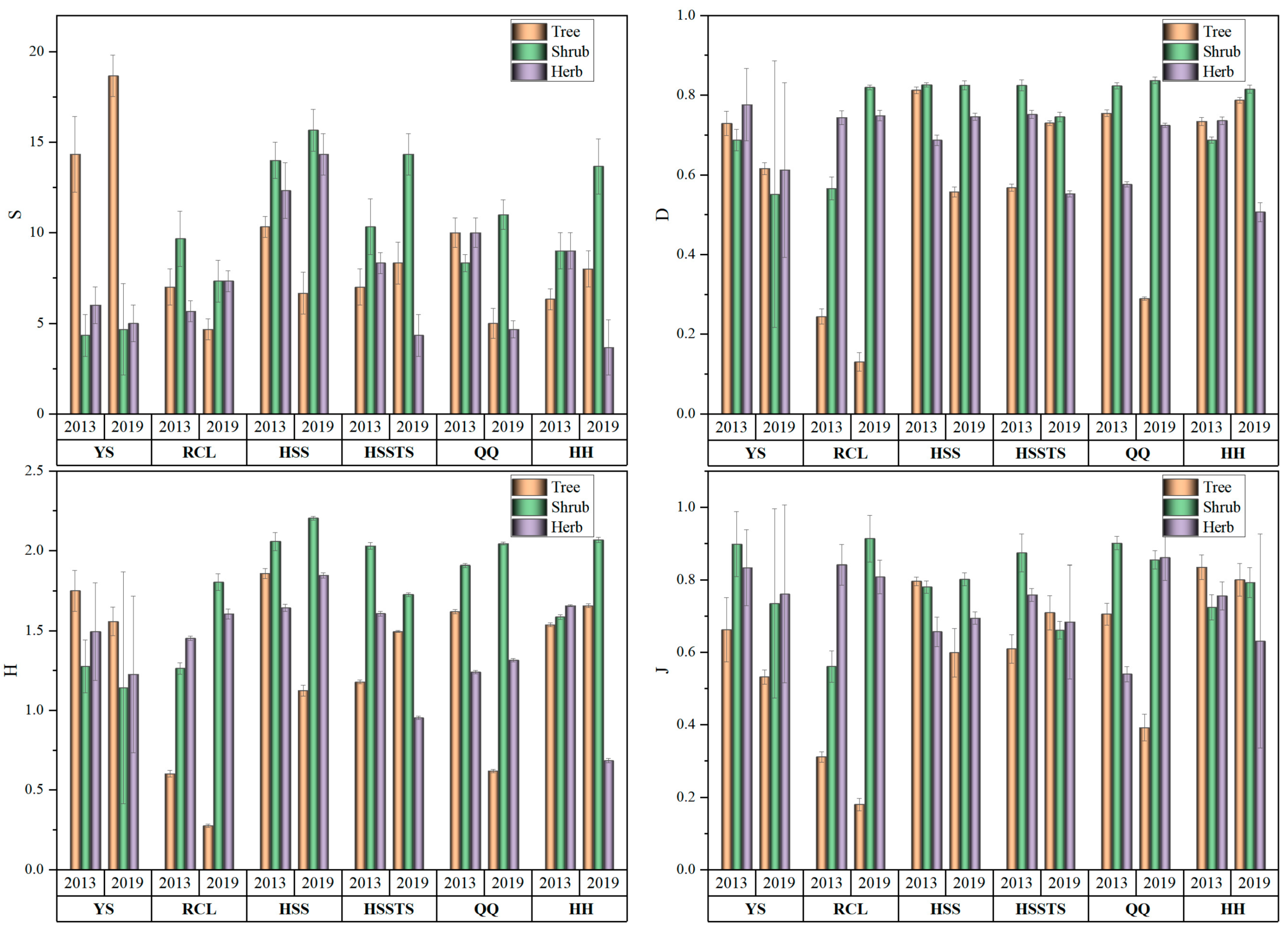
3.1. Species Diversity
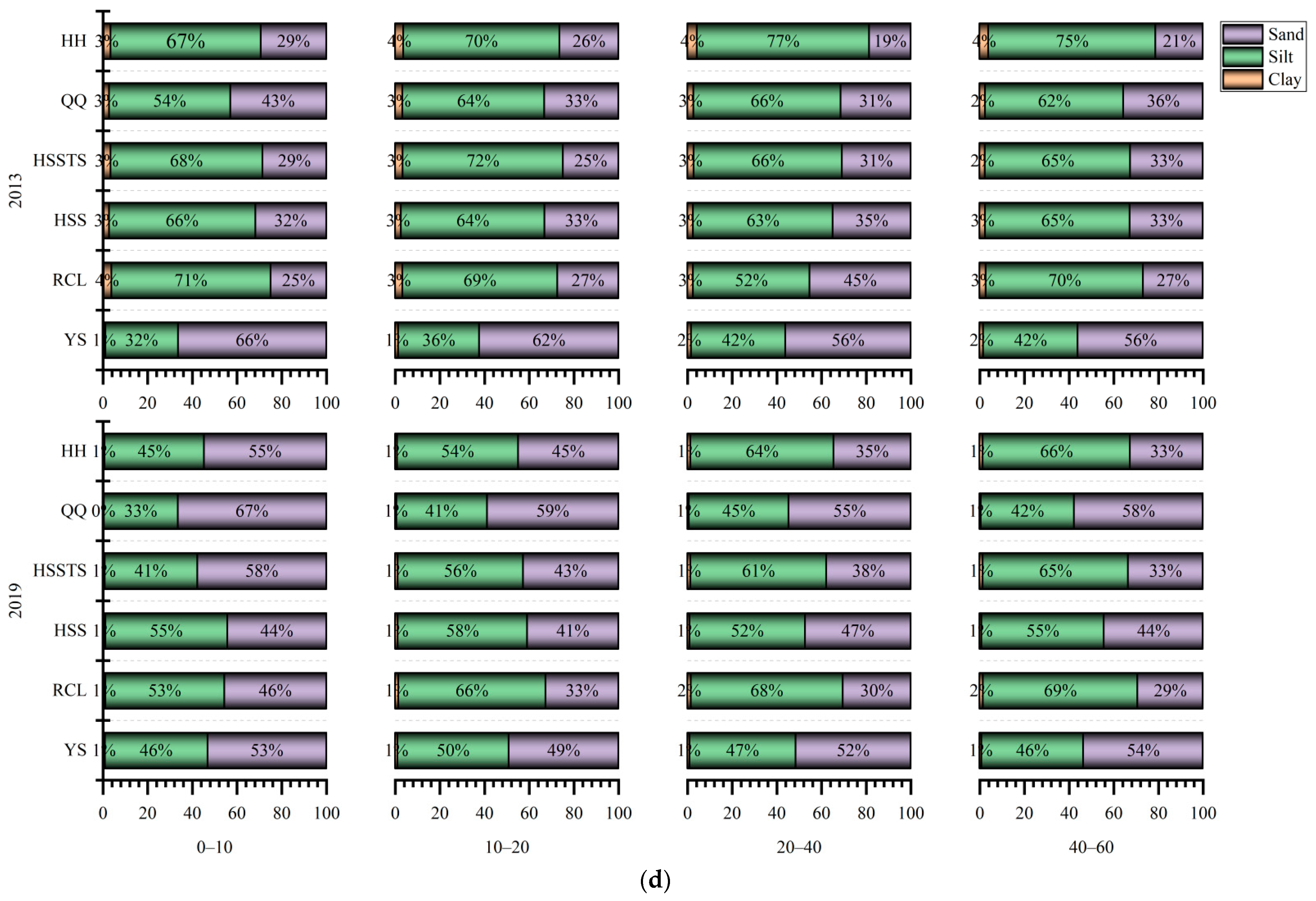
3.2. Soil Physical Properties
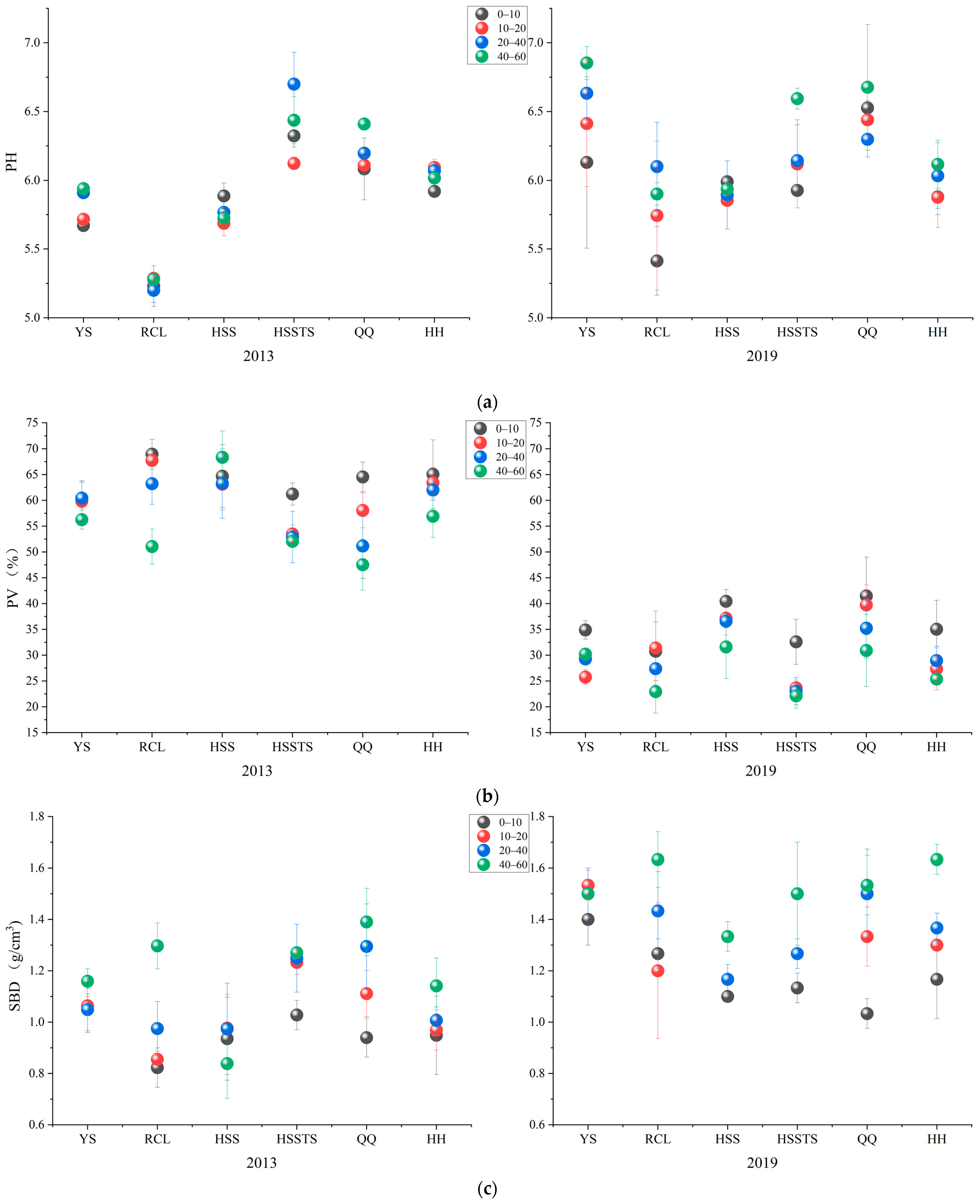
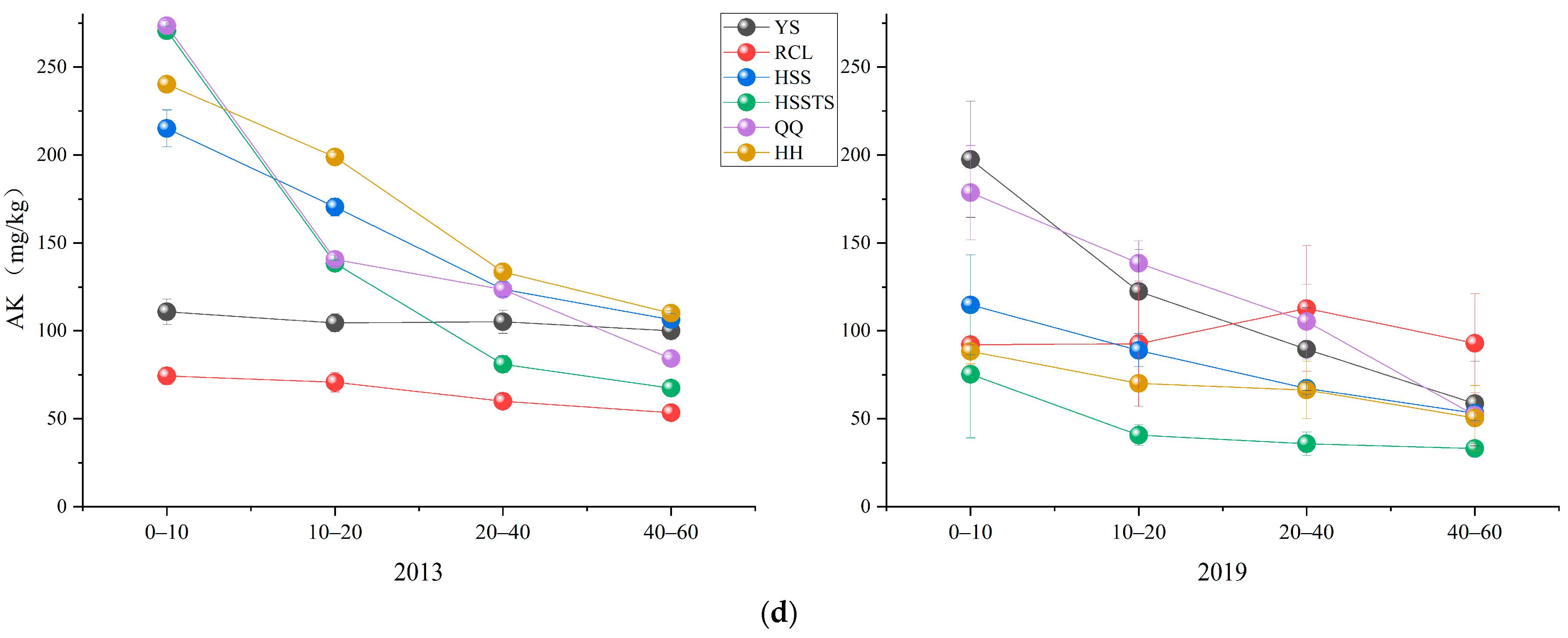
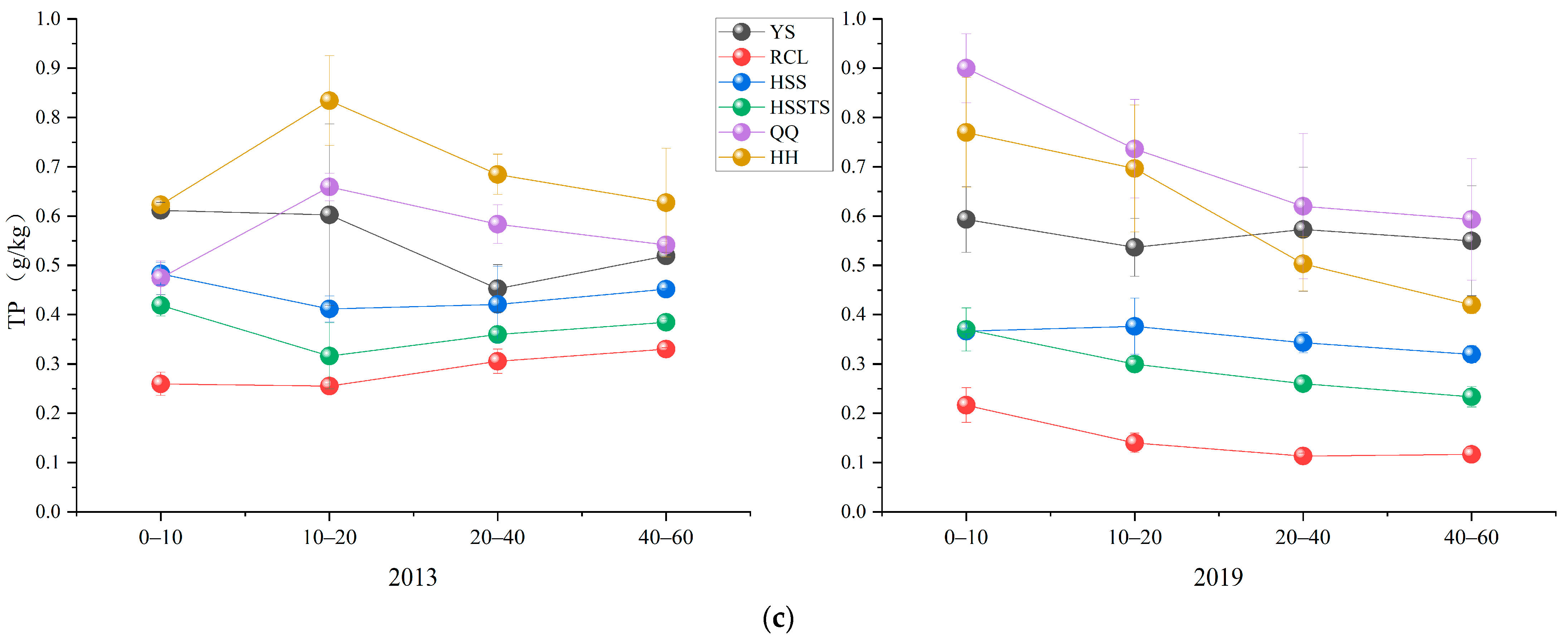
3.3. Soil Chemical Properties
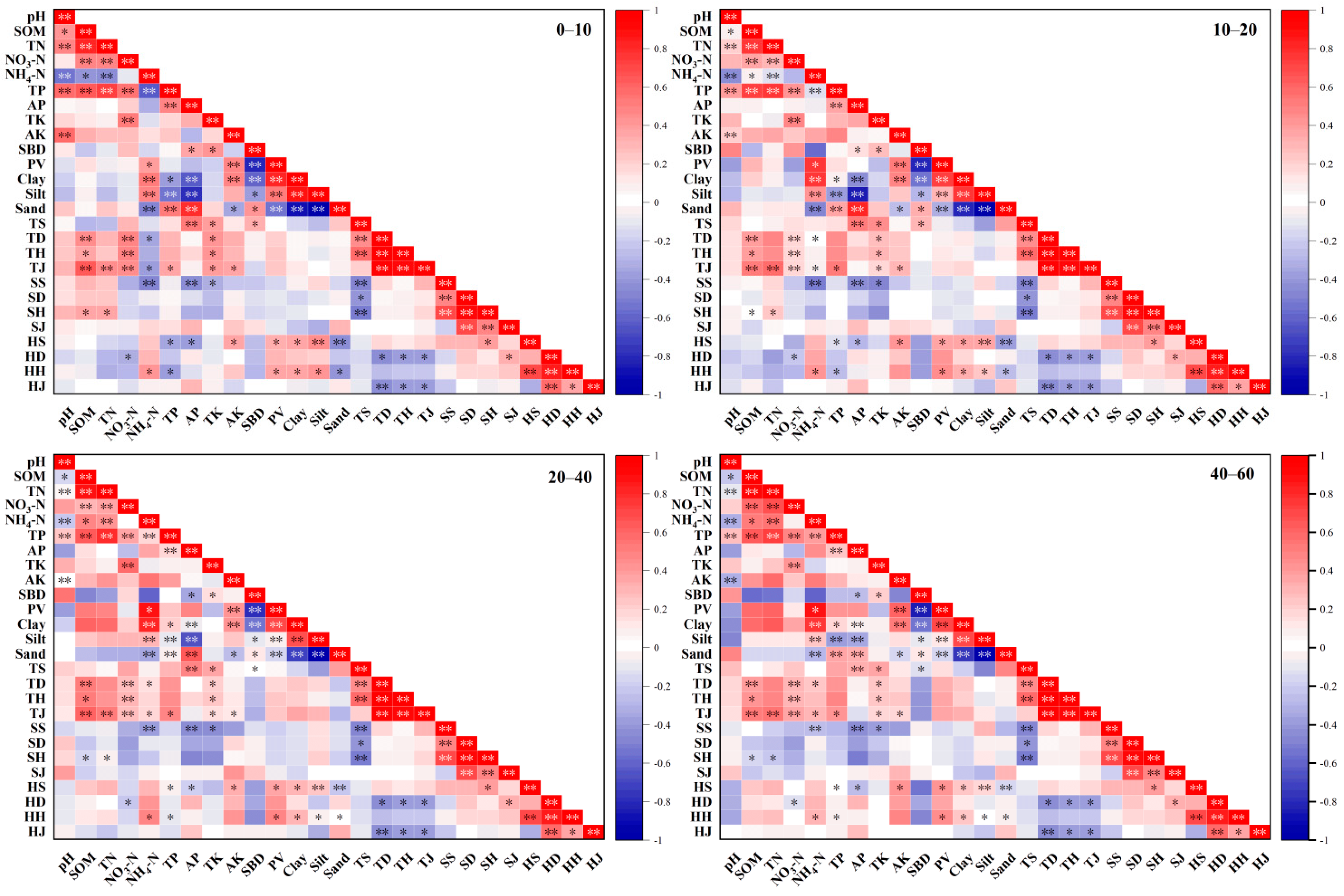
3.4. Correlation between Plant Diversity and Soil Physicochemical Properties
4. Discussion and Conclusions
4.1. Changes in Soil Nutrients in Different Forest Communities
4.2. Plant Diversity in Different Communities
4.3. Relationship between Plant Diversity and Soil Physicochemical Properties
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mureva, A.; Ward, D. Soil microbial biomass and functional diversity in shrub-encroached grasslands along a precipitation gradient. Pedobiologia 2017, 63, 37–45. [Google Scholar] [CrossRef]
- Ma, T.; Deng, X.; Chen, L.; Xiang, W. The soil properties and their effects on plant diversity in different degrees of rocky desertification. Sci. Total Environ. 2020, 736, 139667. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-H.; Huang, B.-H.; Huang, C.-L.; Li, G.; Liao, P.-C. The Aboveground Vegetation Type and Underground Soil Property Mediate the Divergence of Soil Microbiomes and the Biological Interactions. Microb. Ecol. 2018, 75, 434–446. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Eisenhauer, N.; Sierra, C.A.; Bessler, H.; Engels, C.; Griffiths, R.; Mellado-Vázquez, P.G.; Malik, A.A.; Roy, J.; Scheu, S.; et al. Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 2015, 6, 6707. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Xiang, W.; Ouyang, S.; Forrester, D.I.; Zhou, B.; Chen, L.; Ge, T.; Lei, P.; Chen, L.; Zeng, Y.; et al. Linkage between tree species richness and soil microbial diversity improves phosphorus bioavailability. Funct. Ecol. 2019, 33, 1549–1560. [Google Scholar] [CrossRef]
- Hou, L.; Zhang, Y.; Li, Z.; Shao, G.; Song, L.; Sun, Q. Comparison of Soil Properties, Understory Vegetation Species Diversities and Soil Microbial Diversities between Chinese Fir Plantation and Close-to-Natural Forest. Forests 2021, 12, 632. [Google Scholar] [CrossRef]
- Liu, X.; Tan, N.; Zhou, G.; Zhang, D.; Zhang, Q.; Liu, S.; Chu, G.; Liu, J. Plant diversity and species turnover co-regulate soil nitrogen and phosphorus availability in Dinghushan forests, southern China. Plant Soil 2021, 464, 257–272. [Google Scholar] [CrossRef]
- Teste, F.P.; Kardol, P.; Turner, B.L.; Wardle, D.A.; Zemunik, G.; Renton, M.; Laliberté, E. Plant-soil feedback and the mainte-nance of diversity in Mediterranean-climate shrublands. Science 2017, 355, 173–176. [Google Scholar] [CrossRef] [Green Version]
- Van Nuland, M.E.; Ware, I.M.; Bailey, J.K.; Schweitzer, J.A. Ecosystem feedbacks contribute to geographic variation in plant–soil eco-evolutionary dynamics across a fertility gradient. Funct. Ecol. 2019, 33, 95–106. [Google Scholar] [CrossRef] [Green Version]
- da Silva, A.P.; Babujia, L.C.; Franchini, J.C.; Ralisch, R.; Hungria, M.; Guimarães, M.D.F. Soil structure and its influence on microbial biomass in different soil and crop management systems. Soil Tillage Res. 2014, 142, 42–53. [Google Scholar] [CrossRef]
- Gong, Y.; Ling, H.; Lv, G.; Chen, Y.; Guo, Z.; Cao, J. Disentangling the influence of aridity and salinity on community functional and phylogenetic diversity in local dryland vegetation. Sci. Total Environ. 2019, 653, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Levine, J.M.; HilleRisLambers, J. The importance of niches for the maintenance of species diversity. Nature 2009, 461, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Marschner, P.; Cao, W.; Zuo, C.; Qin, W. Influence of salinity and water content on soil microorganisms. Int. Soil Water Conserv. Res. 2015, 3, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Damgaard, C.; Ejrnaes, R.; Stevens, C.J. Intra-specific spatial aggregation in acidic grasslands: Effects of acidification and nitrogen deposition on spatial patterns of plant communities. J. Veg. Sci. 2013, 24, 25–32. [Google Scholar] [CrossRef]
- Cambrollé, J.; Muñoz-Vallés, S.; Mancilla-Leytón, J.M.; Andrades-Moreno, L.; Luque, T.; Figueroa, M.E. Effects of soil physicochemical properties on plant performance of Glaucium flavum Crantz. Plant Soil 2014, 386, 185–193. [Google Scholar] [CrossRef]
- Yang, X.; Long, Y.; Sarkar, B.; Li, Y.; Lü, G.; Ali, A.; Yang, J.; Cao, Y.-E. Influence of soil microorganisms and physicochemical properties on plant diversity in an arid desert of Western China. J. For. Res. 2021, 32, 2645–2659. [Google Scholar] [CrossRef]
- Price, J.; Tamme, R.; Gazol, A.; De Bello, F.; Takkis, K.; Uria-Diez, J.; Kasari, L.; Pärtel, M. Within-community environmental variability drives trait variability in species-rich grasslands. J. Veg. Sci. 2017, 28, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Williams, B.M.; Houseman, G.R. Experimental evidence that soil heterogeneity enhances plant diversity during community assembly. J. Plant Ecol. 2014, 7, 461–469. [Google Scholar] [CrossRef]
- Lü, X.; Hu, Y.; Wolf, A.A.; Han, X. Species richness mediates within-species nutrient resorption: Implications for the biodiversity–productivity relationship. J. Ecol. 2019, 107, 2346–2352. [Google Scholar] [CrossRef]
- Ge, X.M.; Scholl, J.P.; Basinger, U.; Huxman, T.E.; Venable, D.L. Functional trait trade-off and species abundance: Insights from a multi-decadal study. Ecol. Lett. 2019, 22, 583–592. [Google Scholar] [CrossRef]
- Turner, B.; Brenes-Arguedas, T.; Condit, R. Pervasive phosphorus limitation of tree species but not communities in tropical forests. Nature 2018, 555, 367–370. [Google Scholar] [CrossRef]
- Chen, L.; Xiang, W.; Wu, H.; Ouyang, S.; Zhou, B.; Zeng, Y.; Chen, Y.; Kuzyakov, Y. Tree species identity surpasses richness in affecting soil microbial richness and community composition in subtropical forests. Soil Biol. Biochem. 2019, 130, 113–121. [Google Scholar] [CrossRef]
- Bakker, M.R.; Brunner, I.; Ashwood, F.; Bjarnadóttir, B.; Bolger, T.; Børja, I.; Carnol, M.; Cudlin, P.; Dalsgaard, L.; Erktan, A.; et al. Belowground Biodiversity Relates Positively to Ecosystem Services of European Forests. Front. For. Glob. Chang. 2019, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Zhang, F.; Xu, Z.Q.; Jia, Y.L.; You, J.M. Some characteristics of Simpson index and the Shannon-Wiener index and their dilution effect. Pratacult. Sci. 2011, 28, 527–531. [Google Scholar]
- Danise, T.; Innangi, M.; Curcio, E.; Fioretto, A. Covariation between plant biodiversity and soil systems in a European beech forest and a black pine plantation: The case of Mount Faito, (Campania, Southern Italy). J. For. Res. 2021, 33, 239–252. [Google Scholar] [CrossRef]
- Antisari, L.V.; Marinari, S.; Dell’Abate, M.; Baffi, C.; Vianello, G. Plant cover and epipedon SOM stability as factors affecting brown soil profile development and microbial activity. Geoderma 2011, 161, 212–224. [Google Scholar] [CrossRef]
- Danise, T.; Innangi, M.; Curcio, E.; Fioretto, A.; Guggenberger, G. Fast Spectrophotometric Method as Alternative for CuO Oxidation to Assess Lignin in Soils with Different Tree Cover. Forests 2020, 11, 1262. [Google Scholar] [CrossRef]
- Innangi, M.; Menta, C.; Pinto, S.; Danise, T.; D’Alessandro, F.; Fioretto, A. Integrating chemical, biological and soil fauna variables during beech leaf litter decay: A partial least squares approach for a comprehensive view of the decomposition process. Appl. Soil Ecol. 2018, 130, 69–78. [Google Scholar] [CrossRef]
- Zarfos, M.R.; Dovciak, M.; Lawrence, G.B.; McDonnell, T.; Sullivan, T.J. Plant richness and composition in hardwood forest understories vary along an acidic deposition and soil-chemical gradient in the northeastern United States. Plant Soil 2019, 438, 461–477. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Zhong, X.; Lin, F.; Liu, C.; Yang, L.; Wang, M.; Chen, G.; Chen, Y.; Yang, Y. Responses of soil phosphorus fractions after nitrogen addition in a subtropical forest ecosystem: Insights from decreased Fe and Al oxides and increased plant roots. Geoderma 2019, 337, 246–255. [Google Scholar] [CrossRef]
- Chen, F.; Kissel, D.E.; West, L.T.; Rickman, D.; Luvall, J.C.; Adkins, W. Mapping surface soil organic carbon for crop fields with remote sensing. J. Soil Water Conserv. 2005, 60, 51–57. [Google Scholar]
- Fioretto, A.; Innangi, M.; De Marco, A.; Menta, C.; Papa, S.; Pellegrino, A.; De Santo, A.V. Discriminating between Seasonal and Chemical Variation in Extracellular Enzyme Activities within Two Italian Beech Forests by Means of Multilevel Models. Forests 2018, 9, 219. [Google Scholar] [CrossRef] [Green Version]
- Yan, B.S.; Sun, L.P.; Li, J.J.; Gao, L.Q.; Qiao, Z.G.; Gao, X.W.; Wu, C.X.; Wang, G.L. Changes of soil enzyme activity and their stoichiometric characteristics during secondary succession of Quercus liaotungensis forests. Chin. J. Ecol. 2022, 1–9. [Google Scholar] [CrossRef]
- Wei, L.; Archaux, F.; Hulin, F.; Bilger, I.; Gosselin, F. Stand attributes or soil micro-environment exert greater influence than management type on understory plant diversity in even-aged oak high forests. For. Ecol. Manag. 2020, 460, 117897. [Google Scholar] [CrossRef]
- Harpole, W.S.; Tilman, D. Grassland species loss resulting from reduced niche dimension. Nature 2007, 446, 791–793. [Google Scholar] [CrossRef]
- Zilliox, C.; Gosselin, F. Tree species diversity and abundance as indicators of understory diversity in French mountain forests: Variations of the relationship in geographical and ecological space. For. Ecol. Manag. 2014, 321, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Chevalier, R.; Archaux, F.; Gosselin, F. Influence of stand attributes and skid trail area on stand-scale ground flora diversity. Can. J. For. Res. 2015, 45, 1816–1826. [Google Scholar] [CrossRef] [Green Version]
- Yaseen, M.; Fan, G.; Zhou, X.; Long, W.; Feng, G. Plant Diversity and Soil Nutrients in a Tropical Coastal Secondary Forest: Association Ordination and Sampling Year Differences. Forests 2022, 13, 376. [Google Scholar] [CrossRef]
- Wang, L.; Wang, P.; Sheng, M.; Tian, J. Ecological stoichiometry and environmental influencing factors of soil nutrients in the karst rocky desertification ecosystem, southwest China. Glob. Ecol. Conserv. 2018, 16, e00449. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Guo, K.; Qiao, X.; Zhao, H.; Wang, S.; Zhang, L.; Cai, X. Effects of nitrogen, phosphorus and potassium addition on the productivity of a karst grassland: Plant functional group and community perspectives. Ecol. Eng. 2018, 117, 84–95. [Google Scholar] [CrossRef]
- Córdova, S.C.; Olk, D.C.; Dietzel, R.N.; Mueller, K.E.; Archontouilis, S.V.; Castellano, M.J. Plant litter quality affects the accumulation rate, composition, and stability of mineral-associated soil organic matter. Soil Biol. Biochem. 2018, 125, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Tipping, E.; Rowe, E.; Evans, C.; Mills, R.; Emmett, B.; Chaplow, J.; Hall, J. N14C: A plant–soil nitrogen and carbon cycling model to simulate terrestrial ecosystem responses to atmospheric nitrogen deposition. Ecol. Model. 2012, 247, 11–26. [Google Scholar] [CrossRef] [Green Version]
- De Groote, S.R.E.; Vanhellemont, M.; Baeten, L.; De Schrijver, A.; Martel, A.; Bonte, D.; Lens, L.; Verheyen, K. Tree species diversity indirectly affects nutrient cycling through the shrub layer and its high-quality litter. Plant Soil 2018, 427, 335–350. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, W.; Niu, K.; Du, G. Realistic loss of plant species diversity decreases soil quality in a Tibetan alpine meadow. Agric. Ecosyst. Environ. 2019, 279, 25–32. [Google Scholar] [CrossRef]
- Liu, J.; Gou, Q.Q.; Wang, G.H.; Zhao, F.X.; Tang, X.J. Changes of vegetation community and soil characteristics of 50 years old artificial caragana korshinskii in sandy-hilly region of northwest shanxi province. J. Soil Water Conserv. 2022, 36, 219–230. [Google Scholar]
- Zemunik, G.; Turner, B.; Lambers, H.; Laliberté, E. Increasing plant species diversity and extreme species turnover accompany declining soil fertility along a long-term chronosequence in a biodiversity hotspot. J. Ecol. 2016, 104, 792–805. [Google Scholar] [CrossRef] [Green Version]
- Gamfeldt, L.; Snall, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Froberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef]
- Morin, X.; Fahse, L.; Scherer-Lorenzen, M.; Bugmann, H. Tree species richness promotes productivity in temperate forests through strong complementarity between species. Ecol. Lett. 2011, 14, 1211–1219. [Google Scholar] [CrossRef]
- Hagen-Thorn, A.; Callesen, I.; Armolaitis, K.; Nihlgård, B. The impact of six European tree species on the chemistry of mineral topsoil in forest plantations on former agricultural land. For. Ecol. Manag. 2004, 195, 373–384. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, S.; Xie, Z.-P.; Staehelin, C. Analysis of a negative plant-soil feedback in a subtropical monsoon forest. J. Ecol. 2012, 100, 1019–1028. [Google Scholar] [CrossRef]
- Xu, H.; Liu, Q.; Wang, S.; Yang, G.; Xue, S. A global meta-analysis of the impacts of exotic plant species invasion on plant diversity and soil properties. Sci. Total Environ. 2021, 810, 152286. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Bezemer, T.M.; Berendse, F. Soil heterogeneity and plant species diversity in experimental grassland communities: Contrasting effects of soil nutrients and pH at different spatial scales. Plant Soil 2019, 442, 497–509. [Google Scholar] [CrossRef] [Green Version]
- Simkin, S.M.; Allen, E.B.; Bowman, W.D.; Clark, C.M.; Belnap, J.; Brooks, M.L.; Cade, B.S.; Collins, S.L.; Geiser, L.H.; Gilliam, F.S.; et al. Conditional vulnerability of plant diversity to atmospheric nitrogen deposition across the United States. Proc. Natl. Acad. Sci. USA 2016, 113, 4086–4091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Härdtle, W.; von Oheimb, G.; Westphal, C. The effects of light and soil conditions on the species richness of the ground vegetation of deciduous forests in northern Germany (Schleswig-Holstein). For. Ecol. Manag. 2003, 182, 327–338. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Altitude (m) | Slope (°) | Aspect (°) | Age (a) | Stand Density (Trees·ha−1) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | I | II | III | I | II | III | I | II | III | I | II | III | |
| YS | 1497 | 1548 | 1690 | 21 | 25 | 20 | 318 | 291 | 310 | 55 | 50 | 55 | 1548 | 1685 | 1576 |
| RCL | 1597 | 1640 | 1758 | 18 | 20 | 23 | 247 | 263 | 255 | 50 | 55 | 50 | 1589 | 1486 | 1575 |
| HSS | 1460 | 1532 | 1760 | 18 | 24 | 27 | 133 | 147 | 155 | 65 | 60 | 60 | 1286 | 1358 | 1395 |
| HSSTS | 1847 | 1863 | 1897 | 30 | 40 | 37 | 109 | 105 | 87 | 60 | 65 | 55 | 1458 | 1281 | 1588 |
| 1980 | 2003 | 2020 | 10 | 12 | 15 | 203 | 210 | 195 | 75 | 68 | 72 | 986 | 1058 | 978 | |
| HH | 2032 | 2051 | 2131 | 14 | 20 | 25 | 348 | 325 | 335 | 45 | 42 | 48 | 1645 | 1785 | 1652 |
| Layer | Diversity Index | Stepwise Regress Model | Multiple Correlation | Integration F | Significance |
|---|---|---|---|---|---|
| 0–10 | TS | TS = 1.075AP + 5.999 | 0.254 | 11.572 | 0.002 |
| 10–20 | TS = 1.539AP + 5.737 | 0.272 | 12.702 | 0.001 | |
| 20–40 | TS = 1.906AP + 1.673NO3-N + 0.65 | 0.467 | 14.447 | 0 | |
| 40–60 | TS = 0.407TK − 0.17Silt + 11.118 | 0.341 | 8.553 | 0.001 | |
| 0–10 | TD | TD = 0.03NO3-N − 0.013NH4-N + 0.567 | 0.307 | 7.306 | 0.002 |
| 10–20 | TD = 0.18TN + 0.019TK − 0.073 | 0.327 | 8.026 | 0.001 | |
| 20–40 | TD = 0.011SOM + 0.282 | 0.278 | 13.103 | 0.001 | |
| 40–60 | TD = 0.187TN + 0.379 | 0.213 | 9.217 | 0.005 | |
| 0–10 | TH | TH = 0.069NO3-N − 0.026NH4-N + 1.189 | 0.306 | 7.259 | 0.002 |
| 10–20 | TH = 1.033TP − 0.768 | 0.21 | 9.053 | 0.005 | |
| 20–40 | TH = 0.024SOM + 0.615 | 0.275 | 12.879 | 0.001 | |
| 40–60 | TH = 0.422TN + 0.822 | 0.219 | 9.53 | 0.004 | |
| 0–10 | TJ | TJ = 0.018TK + 0.012SOM − 0.383 | 0.49 | 15.852 | 0 |
| 10–20 | TJ = 0.186TN + 0.3 | 0.344 | 17.823 | 0 | |
| 20–40 | TJ = 0.011SOM + 0.285 | 0.373 | 20.197 | 0 | |
| 40–60 | TJ = 0.2TN + 0.381 | 0.301 | 14.644 | 0.001 | |
| 0–10 | SS | SS = −1.337AP − 0.472NH4-N + 20.371 | 0.624 | 27.379 | 0 |
| 10–20 | SS = −1.323AP − 0.495NH4-N + 18.255 | 0.481 | 15.284 | 0 | |
| 20–40 | SS = −1.681AP − 1.461NO3-N + 17.396 | 0.411 | 11.518 | 0 | |
| 40–60 | SS = −1.172AP − 0.503TK + 22.298 | 0.373 | 9.802 | 0 | |
| 20–40 | SD | SD = −0.051NO3-N − 0.048AP + 0.982 | 0.344 | 8.666 | 0.001 |
| 40–60 | SD = −0.068NO3-N − 0.044AP + 1.00 | 0.397 | 10.841 | 0 | |
| 0–10 | SH | SH = 0.278TN − 0.054NO3-N + 1.481 | 0.285 | 6.575 | 0.004 |
| 10–20 | SH = 0.134TN − 0.048TK + 2.449 | 0.242 | 5.279 | 0.01 | |
| 20–40 | SH = −0.196AP − 0.163NO3-N + 2.579 | 0.495 | 16.171 | 0 | |
| 40–60 | SH = −0.188NO3-N − 0.171AP + 2.541 | 0.48 | 15.214 | 0 | |
| 0–10 | SJ | SJ = 00.026AP + 0.001AK + 0.627 | 0.2 | 4.116 | 0.025 |
| 20–40 | SJ = 0.115pH + 0.002AK − 0.06 | 0.326 | 7.979 | 0.001 | |
| 0–10 | HS | HS = 0.11Silt + 1.775 | 0.204 | 8.725 | 0.006 |
| 10–20 | HS = 0.039AK − 0.745NO3-N + 6.153 | 0.376 | 9.957 | 0 | |
| 20–40 | HS = −8.158SBD − 0.977AP + 19.445 | 0.276 | 6.281 | 0.005 | |
| 40–60 | HS = −8.609SBD − 0.892AP + 20.911 | 0.429 | 12.391 | 0 | |
| 10–20 | HD | HD = 0.003PV − 0.072TN + 0.65 | 0.363 | 9.399 | 0.001 |
| 20–40 | HD = 0.004PV − 0.004SOM + 0.591 | 0.279 | 6.378 | 0.005 | |
| 0–10 | HH | HH = 0.094AP − 0.010Sand + 2.007 | 0.271 | 6.125 | 0.005 |
| 10–20 | HH = 0.012PV − 0.013SOM + 1.299 | 0.354 | 9.038 | 0.001 | |
| 20–40 | HH = 0.01PV + 0.927 | 0.223 | 9.784 | 0.004 | |
| 40–60 | HH = 0.006AK + 0.93 | 0.249 | 11.26 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Zhao, M.; Qi, L.; Zhao, C.; Zhang, W.; Zhang, Y.; Wen, W.; Yuan, J. Coupled Relationship between Soil Physicochemical Properties and Plant Diversity in the Process of Vegetation Restoration. Forests 2022, 13, 648. https://doi.org/10.3390/f13050648
Zhao Y, Zhao M, Qi L, Zhao C, Zhang W, Zhang Y, Wen W, Yuan J. Coupled Relationship between Soil Physicochemical Properties and Plant Diversity in the Process of Vegetation Restoration. Forests. 2022; 13(5):648. https://doi.org/10.3390/f13050648
Chicago/Turabian StyleZhao, Yan, Muxin Zhao, Lele Qi, Chensong Zhao, Wenjun Zhang, Yanjun Zhang, Wen Wen, and Jie Yuan. 2022. "Coupled Relationship between Soil Physicochemical Properties and Plant Diversity in the Process of Vegetation Restoration" Forests 13, no. 5: 648. https://doi.org/10.3390/f13050648