
Two Nothofagus Species in Southernmost South America Are Recording Divergent Climate Signals

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Climatic Condition in Southernmost South America

2.2. Tree-Ring Sample from Southernmost South America
2.3. Samples Preparation and Chronology Building
2.4. Principal Component and Regime Shift Analyses
2.5. Climate Data
3. Results
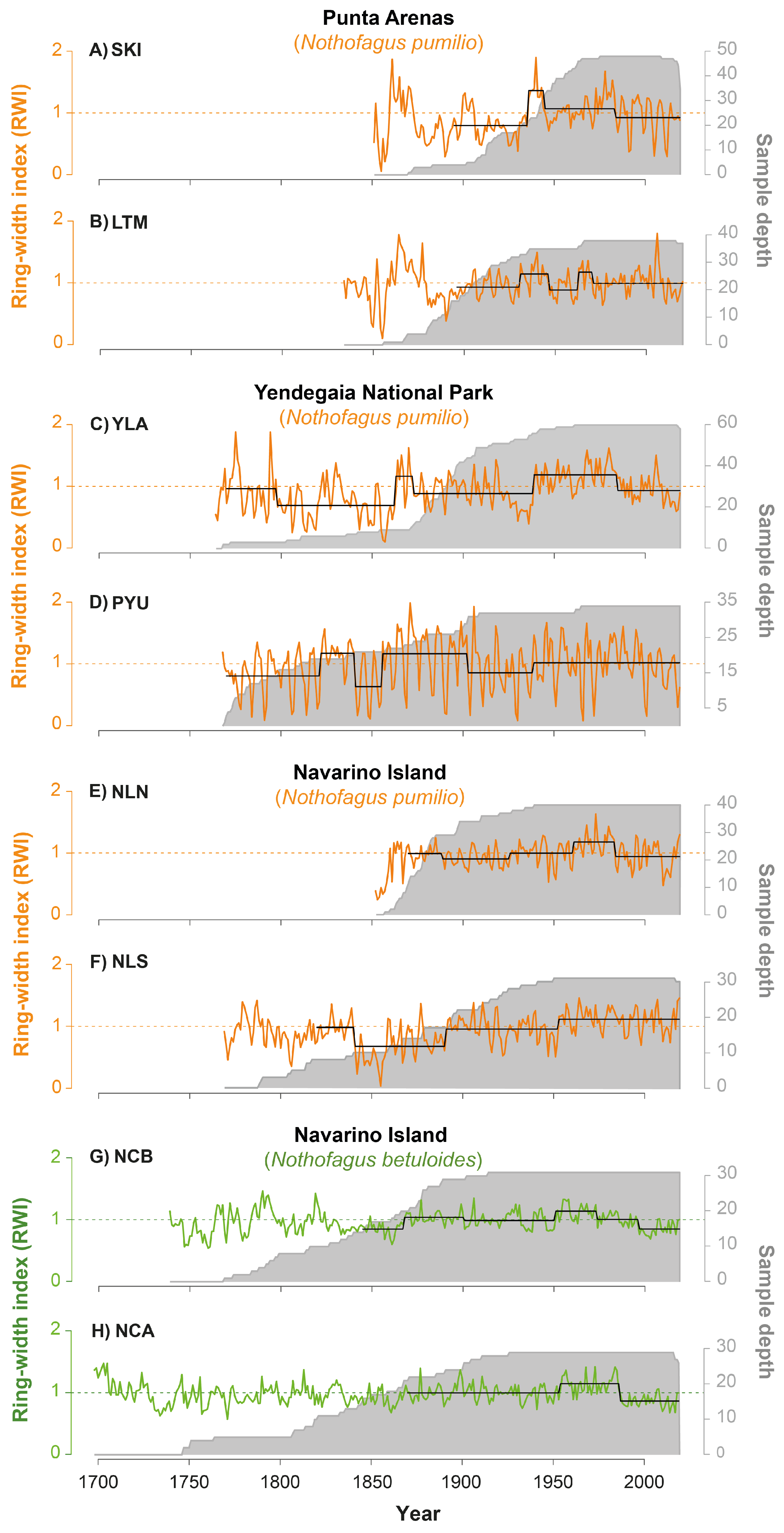
3.1. Tree Growth Patterns
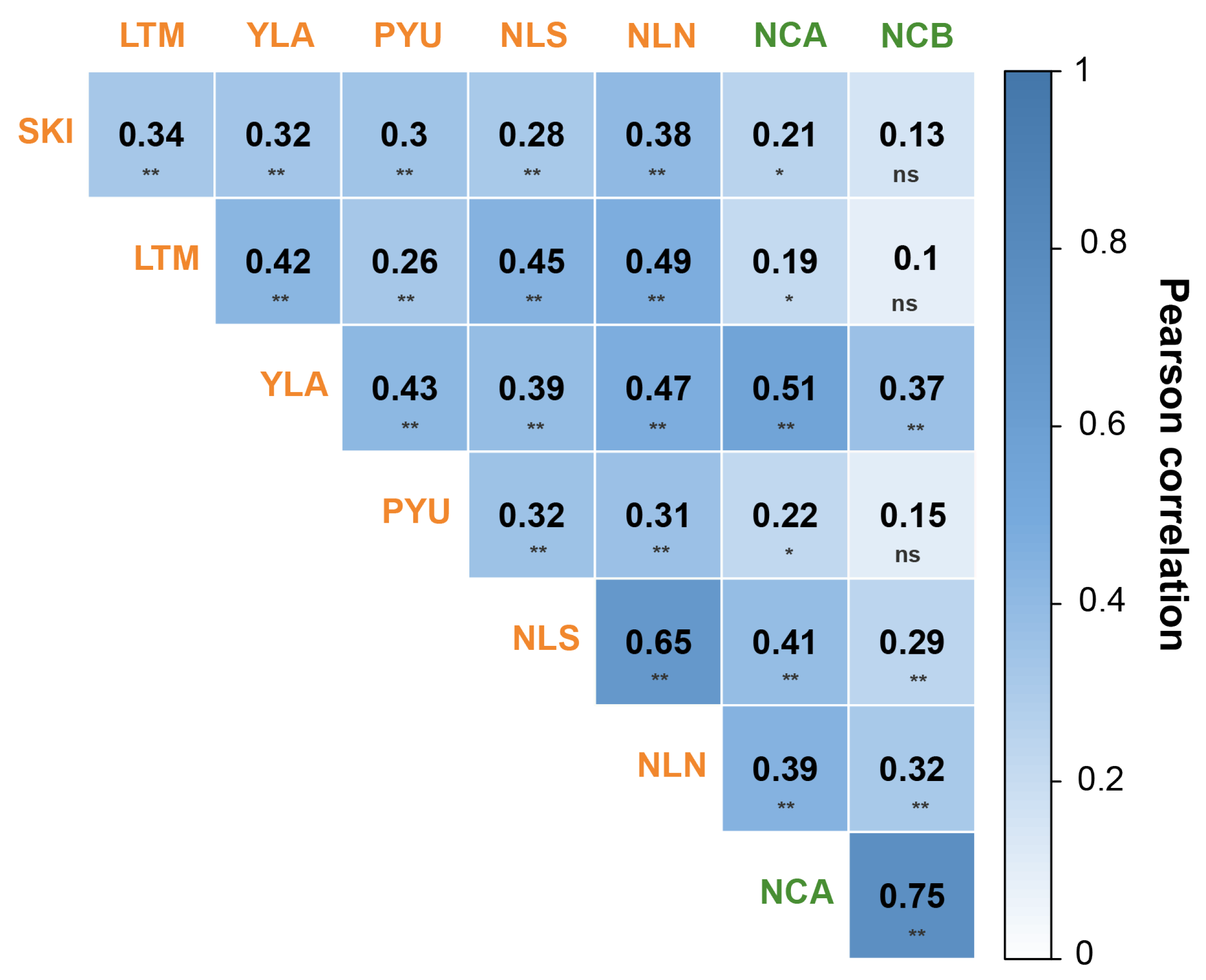
3.2. Relationships between the Individual Ring-Width Index Chronologies
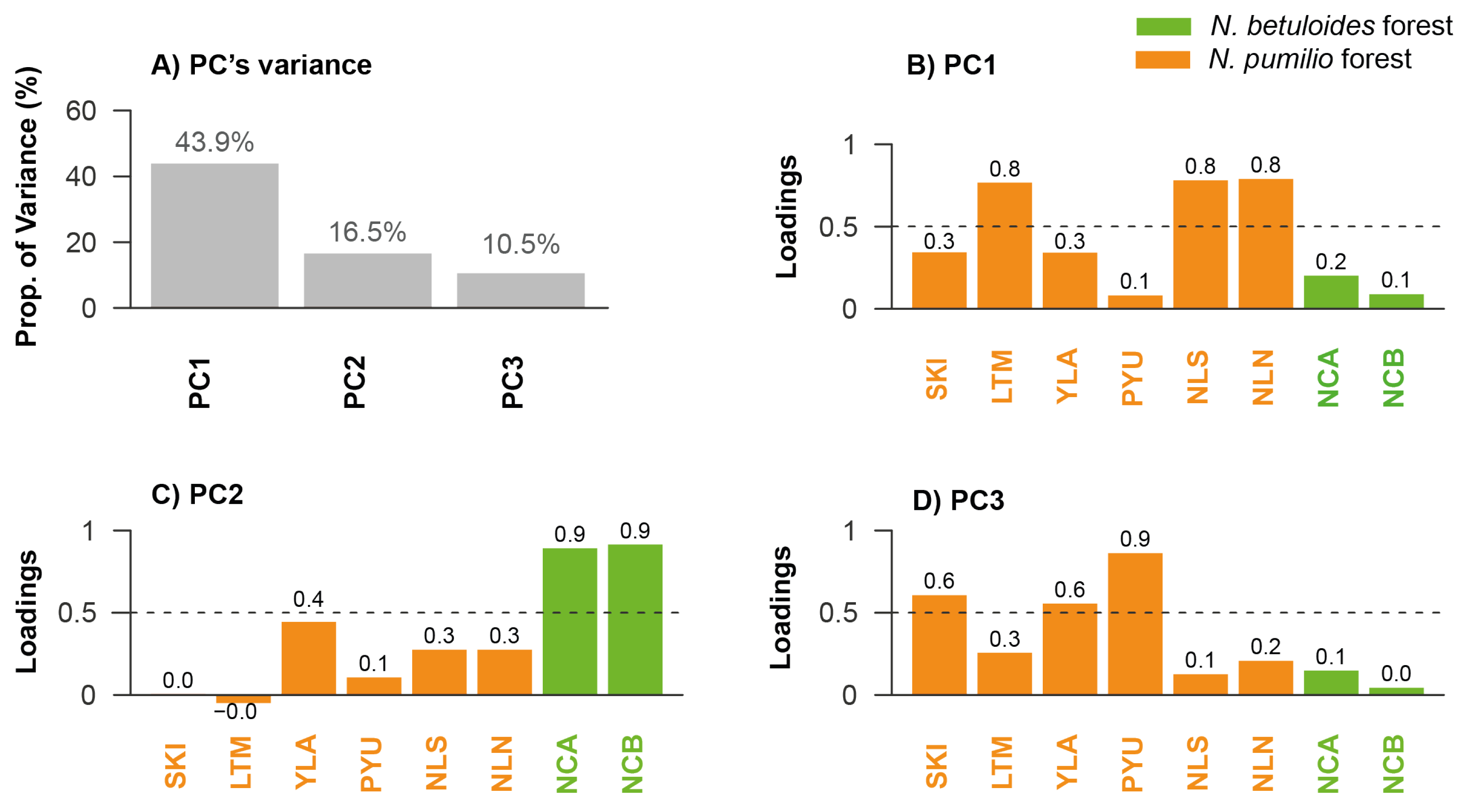
3.3. Evaluation of Regional Tree Growth Patterns
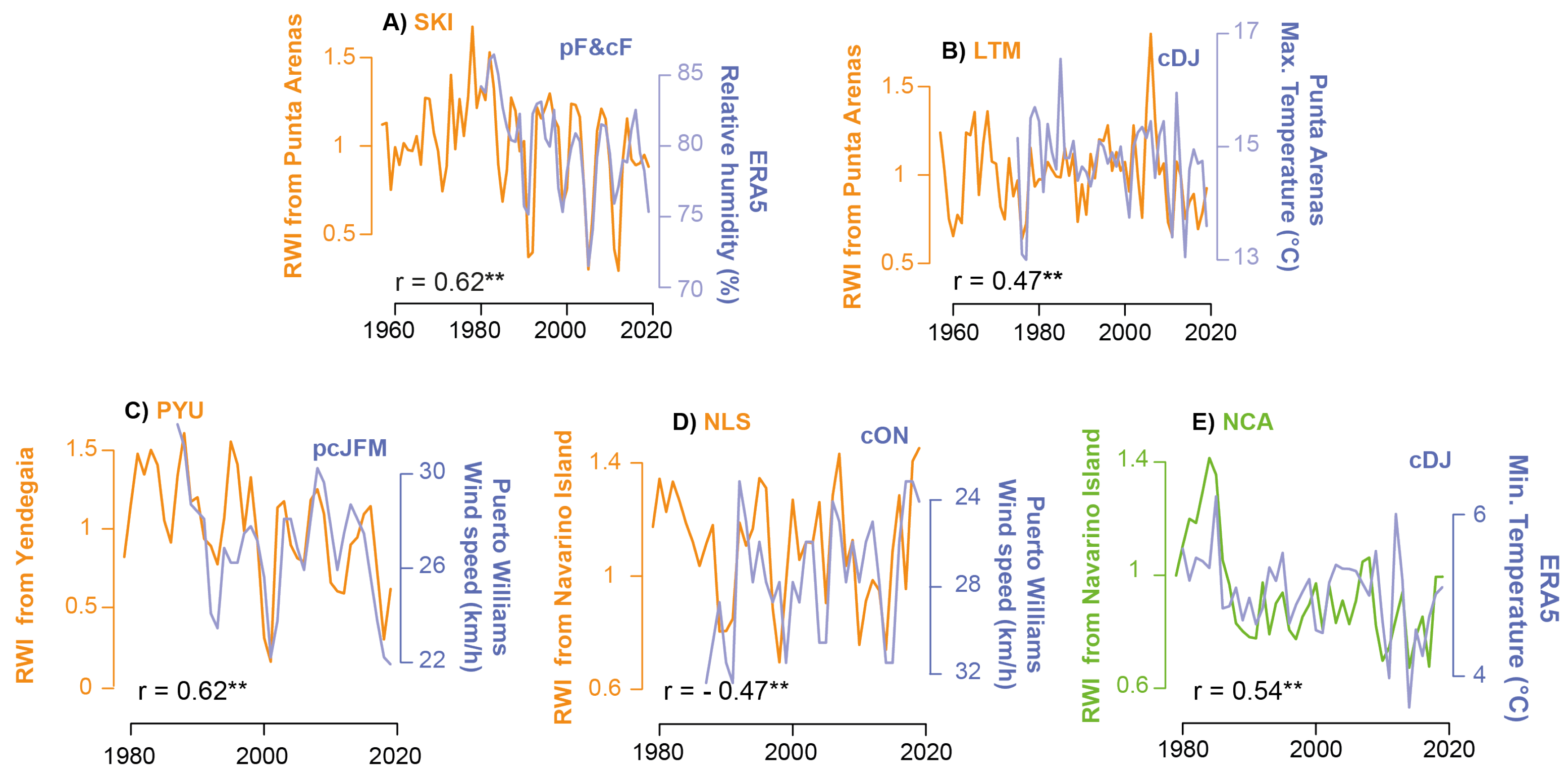
3.4. Influence of Local Climate on the RWI Chronologies
3.5. Influence of Large-Scale Circulation Forcings on the RWI Chronologies
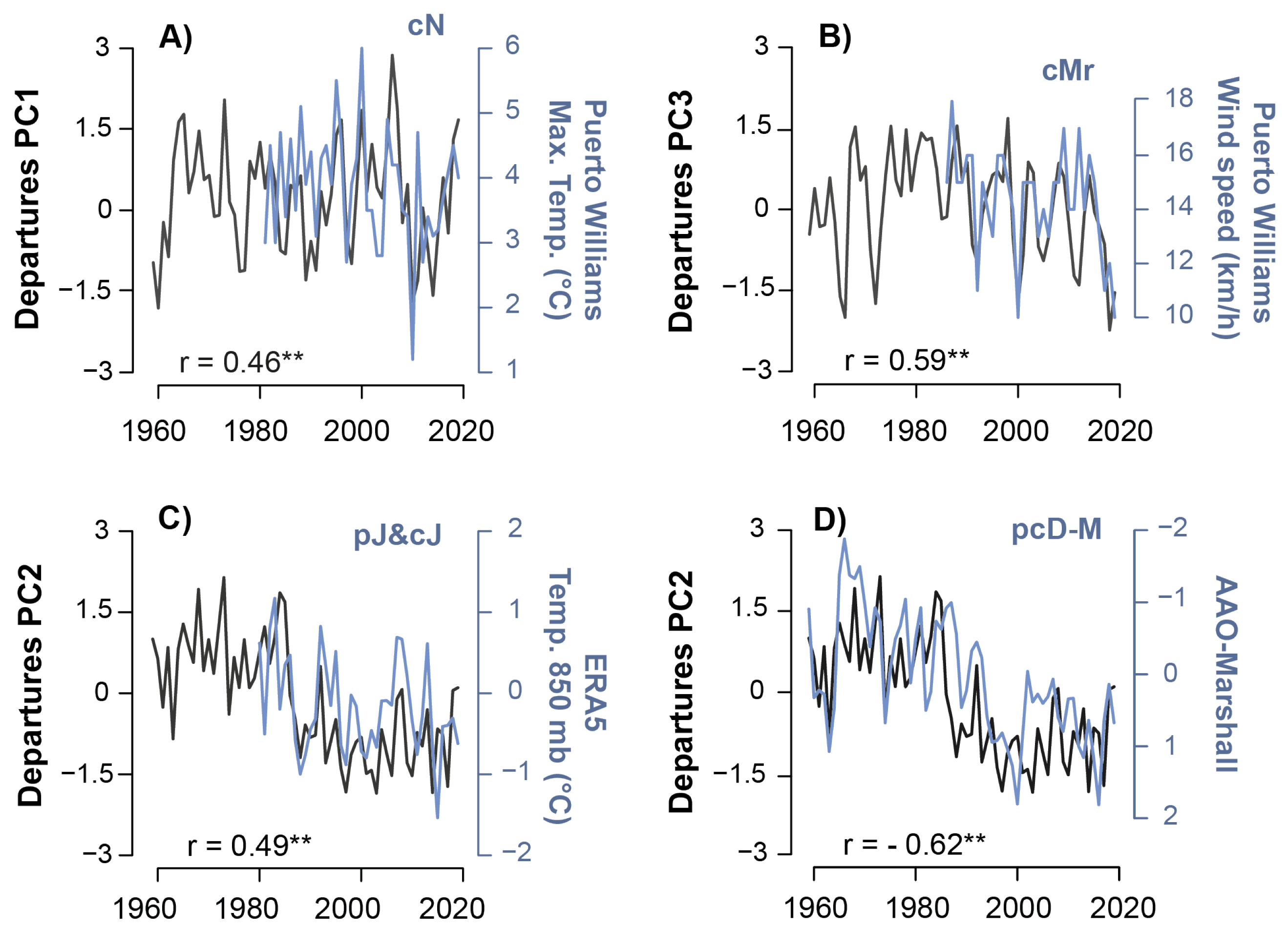
3.6. Influence of the Local and Large-Scale (AAO) Climate on Principal Components
4. Discussion
4.1. Dendrochronological and Dendroclimatic Potential of the Southernmost Nothofagus Forests
4.2. High Diversity of Climatic Responses in SSA Nothofagus Forests
4.3. Different Intra- and Inter-Species Sensitivities to Climate
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.T.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S.W. Drought-Induced Reduction in Global Terrestrial Net Primary Production from 2000 Through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [Green Version]
- Anderegg, W.R.L.; Schwalm, C.; Biondi, F.; Camarero, J.J.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.G.; Allen, C.D.; Anderson-Teixeira, K.; Aukema, B.H.; Bond-Lamberty, B.; Chini, L.; Clark, J.S.; Dietze, M.; Grossiord, C.; Hanbury-Brown, A.; et al. Pervasive shifts in forest dynamics in a changing world. Science 2020, 368, eaaz9463. [Google Scholar] [CrossRef]
- Büntgen, U.; Urban, O.; Krusic, P.J.; Rybníček, M.; Kolář, T.; Kyncl, T.; Ač, A.; Koňasová, E.; Čáslavský, J.; Esper, J.; et al. Recent European drought extremes beyond Common Era background variability. Nat. Geosci. 2021, 14, 190–196. [Google Scholar] [CrossRef]
- Arias, P.; Bellouin, N.; Coppola, E.; Jones, R.; Krinner, G.; Marotzke, J.; Naik, V.; Palmer, M.; Plattner, G.K.; Rogelj, J.; et al. Technical Summary. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 33–144. [Google Scholar]
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate forest trees and stands under severe drought: A review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef] [Green Version]
- Villalba, R.; Veblen, T.T. Influences of Large-Scale Climatic Variability on Episodic Tree Mortality in Northern Patagonia. Ecology 1998, 79, 2624. [Google Scholar] [CrossRef]
- Suarez, M.L.; Ghermandi, L.; Kitzberger, T. Factors predisposing episodic drought-induced tree mortality in Nothofagus—Site, climatic sensitivity and growth trends. J. Ecol. 2004, 92, 954–966. [Google Scholar] [CrossRef]
- Mundo, I.A.; Mujtar, V.A.E.; Perdomo, M.H.; Gallo, L.A.; Villalba, R.; Barrera, M.D. Austrocedrus chilensis growth decline in relation to drought events in northern Patagonia, Argentina. Trees 2010, 24, 561–570. [Google Scholar] [CrossRef]
- Rodríguez-Catón, M.; Villalba, R.; Srur, A.M.; Luckman, B. Long-term trends in radial growth associated with Nothofagus pumilio forest decline in Patagonia: Integrating local- into regional-scale patterns. For. Ecol. Manag. 2015, 339, 44–56. [Google Scholar] [CrossRef]
- Rodríguez-Catón, M.; Villalba, R.; Morales, M.; Srur, A. Influence of droughts on Nothofagus pumilio forest decline across northern Patagonia, Argentina. Ecosphere 2016, 7, e01390. [Google Scholar] [CrossRef]
- Tarabini, M.; Gomez, F.; Ángel Calderón, M.; Manna, L.L. Role of abiotic factors in Nothofagus pumilio forest mortality: The sensitivity of ecotones. For. Ecol. Manag. 2021, 494, 119316. [Google Scholar] [CrossRef]
- Urrutia-Jalabert, R.; Barichivich, J.; Rozas, V.; Lara, A.; Rojas, Y.; Bahamondez, C.; Rojas-Badilla, M.; Gipoulou-Zuñiga, T.; Cuq, E. Climate response and drought resilience of Nothofagus obliqua secondary forests across a latitudinal gradient in south-central Chile. For. Ecol. Manag. 2021, 485, 118962. [Google Scholar] [CrossRef]
- Puchi, P.F.; Camarero, J.J.; Battipaglia, G.; Carrer, M. Retrospective analysis of wood anatomical traits and tree-ring isotopes suggests site-specific mechanisms triggering Araucaria araucana drought-induced dieback. Glob. Chang. Biol. 2021, 27, 6394–6408. [Google Scholar] [CrossRef]
- Urrutia-Jalabert, R.; Malhi, Y.; Barichivich, J.; Lara, A.; Delgado-Huertas, A.; Rodríguez, C.G.; Cuq, E. Increased water use efficiency but contrasting tree growth patterns in Fitzroya cupressoides forests of southern Chile during recent decades. J. Geophys. Res. Biogeosci. 2015, 120, 2505–2524. [Google Scholar] [CrossRef]
- Lavergne, A.; Daux, V.; Villalba, R.; Pierre, M.; Stievenard, M.; Srur, A.M. Improvement of isotope-based climate reconstructions in Patagonia through a better understanding of climate influences on isotopic fractionation in tree rings. Earth Planet. Sci. Lett. 2017, 459, 372–380. [Google Scholar] [CrossRef]
- Pisano, E. Fitogeografía de Fuego-Patagonia chilena I- Comunidades vegetales entre las latitudes 52 y 56 s. An. Inst. Patagon. 1977, 8, 121–250. [Google Scholar]
- Donoso, C. Tipos Forestales de los Bosques Nativos de Chile; Santiago, Chile, 1981; p. 70. Available online: https://bibliotecadigital.infor.cl/handle/20.500.12220/6178 (accessed on 1 March 2022).
- Srur, A.M.; Villalba, R.; Villagra, P.E.; Hertel, D. Influencias de las variaciones en el clima y en la concentración de CO2 sobre el crecimiento de Nothofagus pumilio en la Patagon. Rev. Chil. Hist. Nat. 2008, 81, 239–256. [Google Scholar] [CrossRef]
- Llancabure, J.C. Relaciones entre el Crecimiento de Nothofagus betuloides y el Clima Local y de Gran Escala en Bosques Subantárticos de la Isla Navarino. Bachelor’s Thesis, Universidad Austral de Chile, Valdivia, Chile, 2011. [Google Scholar]
- Villalba, R.; Lara, A.; Masiokas, M.; Urrutia, R.; Luckman, B.H.; Marshall, G.J.; Mundo, I.A.; Christie, D.A.; Cook, E.R.; Neukom, R.; et al. Unusual Southern Hemisphere tree growth patterns induced by changes in the Southern Annular Mode. Nat. Geosci. 2012, 5, 793–798. [Google Scholar] [CrossRef]
- Soto-Rogel, P.; Aravena, J. Potencial dendroclimático de Nothofagus betuloides en la cordillera de Darwin, Tierra del Fuego, Chile. Bosque 2017, 38, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, M.; Aravena, J.C.; Seim, A.; Linderholm, H.W. Assessing the dendroclimatic potential of Nothofagus betuloides (Magellan’s beech) forests in the southernmost Chilean Patagonia. Trees 2019, 33, 557–575. [Google Scholar] [CrossRef] [Green Version]
- Marshall, G.J. Trends in the Southern Annular Mode from observations and reanalyses. J. Clim. 2003, 16, 4134–4143. [Google Scholar] [CrossRef]
- Carrasco, J.F. Decadal Changes in the Near-Surface Air Temperature in the Western Side of the Antarctic Peninsula. Atmos. Clim. Sci. 2013, 3, 33653. [Google Scholar] [CrossRef] [Green Version]
- González-Reyes, Á.; Aravena, J.C.; Muñoz, A.A.; Soto-Rogel, P.; Aguilera-Betti, I.; Toledo-Guerrero, I. Variabilidad de la precipitación en la ciudad de Punta Arenas, Chile, desde principios del siglo XX. An. Inst. Patagon. 2017, 45, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Soto-Rogel, P.; Aravena, J.C.; Meier, W.J.H.; Gross, P.; Pérez, C.; González-Reyes, Á.; Griessinger, J. Impact of Extreme Weather Events on Aboveground Net Primary Productivity and Sheep Production in the Magellan Region, Southernmost Chilean Patagonia. Geosciences 2020, 10, 318. [Google Scholar] [CrossRef]
- Dirección Meteorológica de Chile. Available online: http://www.meteochile.gob.cl/PortalDMC-web/index.xhtml (accessed on 1 May 2022).
- Massaccesi, G.; Roig, F.A.; Martínez Pastur, G.J.; Barrera, M.D. Growth patterns of Nothofagus pumilio trees along altitudinal gradients in Tierra del Fuego, Argentina. Trees 2008, 22, 245–255. [Google Scholar] [CrossRef]
- Soliani, C.; Mattera, M.G.; Marchelli, P.; Azpilicueta, M.M.; Dalla-Salda, G. Different drought-adaptive capacity of a native Patagonian tree species (Nothofagus pumilio) resulting from local adaptation. Eur. J. For. Res. 2021, 140, 1147–1161. [Google Scholar] [CrossRef]
- Carrasco, J.F.; Casassa, G.; Rivera, A. Meteorological and Climatological Aspects of the Southern Patagonia Icefield. In The Patagonian Icefields: A Unique Natural Laboratory for Environmental and Climate Change Studies; Casassa, G., Sepúlveda, F.V., Sinclair, R.M., Eds.; Springer: Boston, MA, USA, 2002; pp. 29–41. [Google Scholar] [CrossRef]
- Luebert, F.; Pliscoff, P. Sinopsis Bioclimática y Vegetacional de Chile; Editorial Universitaria: Santiago, Chile, 1977; p. 384. [Google Scholar]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Arizona Press: Tucson, AZ, USA, 1996; p. 73. [Google Scholar]
- Schulman, E. Dendroclimatic Changes in Semiarid America; University of Arizona Press: Tucson, AZ, USA, 1956; p. 142. [Google Scholar]
- Robinson, W.J.; Evans, R. A Microcomputer-Based Tree-Ring Measuring System. Tree-Ring Bull. 1980, 40, 59–64. [Google Scholar]
- Holmes, R.L. Computer-Assisted Quality Control in Tree-Ring Dating and Measurement. Tree-Ring Bull. 1983, 43, 51–67. [Google Scholar]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK, 1976; p. 567. [Google Scholar]
- Briffa, K.; Jones, P. Basic chronology statistics and assessment. In Methods of Dendrochronology: Applications in the Environmental Sciences; Cook, E., Kairiukstis, L., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; pp. 137–152. [Google Scholar]
- Cook, E.R.; Briffa, K.R.; Jones, P.D. Spatial Regression Methods in Dendroclimatology: A review and comparison of two techniques. Int. J. Climatol. 1994, 14, 379–402. [Google Scholar] [CrossRef]
- Wilks, D.S. Statistical Methods in the Atmospheric Sciences; Elsevier Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2011. [Google Scholar]
- Revelle, W. psych: Procedures for Psychological, Psychometric, and Personality Research; R Package Version 2.1.9; Northwestern University: Evanston, IL, USA, 2021. [Google Scholar]
- Rodionov, S.N. A sequential algorithm for testing climate regime shifts. Geophys. Res. Lett. 2004, 31. [Google Scholar] [CrossRef] [Green Version]
- Hersbach, H.; Bell, B.; Berrisford, P.; Hirahara, S.; Horányi, A.; Muñoz-Sabater, J.; Nicolas, J.P.; Peubey, C.; Radu, R.; Schepers, D.; et al. The ERA5 global reanalysis. Q. J. R. Meteorol. Soc. 2020, 146, 1999–2049. [Google Scholar] [CrossRef]
- Climate Prediction Center. Available online: https://www.cpc.ncep.noaa.gov/products/precip/CWlink/daily_ao_index/aao/aao.shtml (accessed on 1 May 2022).
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the Average Value of Correlated Time Series, with Applications in Dendroclimatology and Hydrometeorology. J. Appl. Meteorol. Climatol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Veblen, T.T.; Kitzberger, T.; Donoso, C.; Rebertus, A.J. Ecology of southern Chilean and Argentinean Nothofagus forests. In The Ecology and Biogeography of Nothofagus Forests; Veblen, T.T., Hill, R.S., Read, J., Eds.; Yale University Press: New Haven, CT, USA, 1996; p. 403. [Google Scholar]
- Aravena, J.C.; Lara, A.; Wlodarsky-FrankeE, A.; Villalba, R.; Cuq, E. Tree-ring growth patterns and temperature reconstruction from Nothofagus pumilio (Fagaceae) forests at the upper tree line of southern Chilean Patagonia. Rev. Chil. Hist. Nat. 2002, 75, 361–376. [Google Scholar] [CrossRef]
- Villalba, R.; Lara, A.; Boninsegna, J.A.; Masiokas, M.; Delgado, S.; Aravena, J.C.; Roig, F.A.; Schmelter, A.; Wolodarsky, A.; Ripalta, A. Large-scale temperature changes across the Southern Andes: 20th-century variations in the context of the past 400 years. Clim. Chang. 2003, 59, 177–232. [Google Scholar] [CrossRef]
- Matskovsky, V.; Roig, F.A.; Martínez Pastur, G. Removal of a non-climatically induced seven-year cycle from Nothofagus pumilio tree-ring width chronologies from Tierra del Fuego, Argentina for their use in climate reconstructions. Dendrochronologia 2019, 57, 125610. [Google Scholar] [CrossRef]
- Varela, S.A.; Gyenge, J.E.; Fernández, M.E.; Schlichter, T. Seedling drought stress susceptibility in two deciduous Nothofagus species of NW Patagonia. Trees 2010, 24, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Bucci, S.J.; Silletta, L.M.C.; Garré, A.; Cavallaro, A.; Efron, S.T.; Arias, N.S.; Goldstein, G.; Scholz, F.G. Functional relationships between hydraulic traits and the timing of diurnal depression of photosynthesis. Plant Cell Environ. 2019, 42, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Meier, W.H.; Aravena, J.C.; Jaña, R.; Braun, M.; Hochreuther, P.; Soto-Rogel, P.; Grießinger, J. A tree-ring d18O series from southernmost Fuego-Patagonia is recording flavors of the Antarctic Oscillation. Glob. Planet. Chang. 2020, 195, 103302. [Google Scholar] [CrossRef]
- Han, Y.; Wang, Y.; Liu, B.; Huang, R.; Camarero, J.J. Moisture mediates temperature-growth couplings of high-elevation shrubs in the Tibetan plateau. Trees 2022, 36, 273–281. [Google Scholar] [CrossRef]
- Mueller, B.; Seneviratne, S.I. Hot days induced by precipitation deficits at the global scale. Proc. Natl. Acad. Sci. USA 2012, 109, 12398–12403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J. Compound climate events increase tree drought mortality across European forests. Sci. Total Environ. 2021, 816, 151604. [Google Scholar] [CrossRef]
- Boninsegna, J.A.; Keegan, J.; Jacoby, G.C.; D’Arrigo, R.; Holmes, R.L. Dendrochronological studies in Tierra del Fuego, Argentina. In Quaternary of South America and Antarctic Peninsula; Rabassa, J., Ed.; CRC Press: Boca Raton, FL, USA, 1989; pp. 305–326. [Google Scholar]
- Smith, V.C.; Ennos, A.R. The effects of air flow and stem flexure on the mechanical and hydraulic properties of the stems of sunflowers Helianthus annuus L. J. Exp. Bot. 2003, 54, 845–849. [Google Scholar] [CrossRef]
- Iogna, P.A.; Bucci, S.J.; Scholz, F.G.; Goldstein, G. Homeostasis in leaf water potentials on leeward and windward sides of desert shrub crowns: Water loss control vs. high hydraulic efficiency. Oecologia 2013, 173, 675–687. [Google Scholar] [CrossRef]
- Seo, J.W.; Choi, E.B.; Park, J.H.; Kim, Y.J.; Lim, H.I. The Role of Aging and Wind in Inducing Death and/or Growth Reduction in Korean Fir (Abies Koreana Wilson) on Mt. Halla, Korea. Atmosphere 2021, 12, 1135. [Google Scholar] [CrossRef]
- Bahamonde, H.A.; Sánchez-Gómez, D.; Gyenge, J.; Peri, P.L.; Cellini, J.M.; Aranda, I. Thinking in the sustainability of Nothofagus antarctica silvopastoral systems, how differ the responses of seedlings from different provenances to water shortage? Agrofor. Syst. 2019, 93, 689–701. [Google Scholar] [CrossRef]
- Oyarzún, C.; Nahuelhual, L.; Núñez, D. Los servicios ecosistémicos del bosque templado lluvioso: Producción de agua y su valoración económica. Rev. Ambi. Desarro. 2005, 20, 88–95. [Google Scholar]
- Jobbágy, E.G. Servicios hídricos de los ecosistemas y su relación con el uso de la tierra en la llanura Chaco-Pampeana. In Valoración de Servicios Ecosistémicos Conceptos, Herramientas y Aplicaciones para el Ordenamiento Territorial; Laterra, P., Jobbágy, E.G., Paruelo, J., Eds.; Ediciones INTA: New York, NY, USA, 2011; pp. 163–183. [Google Scholar]
- Hoyos-Santillan, J.; Miranda, A.; Lara, A.; Sepulveda-Jauregui, A.; Zamorano-Elgueta, C.; Gómez-González, S.; Vásquez-Lavín, F.; Garreaud, R.D.; Rojas, M. Diversifying Chile’s climate action away from industrial plantations. Environ. Sci. Policy 2021, 124, 85–89. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Code | sp* | Lat (S) Long (W) | Elevation m asl | Timespan AD Years | Number of Trees (Radii) | Trees (Radii) in Chronology | EPS (>0.85) Start.Year |
|---|---|---|---|---|---|---|---|
| Punta Arenas area | |||||||
| SKI | Np | 53°9′36″ 71°1′54″ | 550 | 1851–2019 | 29/(58) | 26 (49) | 1895 |
| LTM | Np | 53°18′59″ 71°16′48″ | 558 | 1834–2020 | 32/(64) | 27 (40) | 1896 |
| Yendegaia Valley area | |||||||
| YLA | Np | 54°48′35″ 68°48′35″ | 566 | 1764–2019 | 33/(66) | 31 (61) | 1770 |
| PYU | Np | 54°50′30″ 68°38′39″ | 423 | 1768–2019 | 22/(44) | 21 (35) | 1770 |
| Navarino Island area | |||||||
| NLS | Np | 55°00′38″ 67°36′35″ | 450 | 1769–2019 | 20/(40) | 20 (32) | 1820 |
| NLN | Np | 54°59′45″ 67°36′00″ | 403 | 1852–2019 | 21/(42) | 21 (41) | 1870 |
| NCA | Nb | 54°59′29″ 67°36′9″ | 350 | 1699–2019 | 21 /(42) | 20 (30) | 1870 |
| NCB | Nb | 54°59′1″ 67°36′32″ | 275 | 1739–2019 | 21 /(42) | 19 (32) | 1845 |
| Weather Station | Lat (S) Long (W) | Variable | Period | Source |
|---|---|---|---|---|
| Punta Arenas1 | pre: precipitation | 1900–2019 | ||
| (puq1) | 53°0′6″ | tmn: minimum temperature | 1930–2019 | DMC |
| 70°50′50″ | tmx: maximum temperature | 1930–2019 | ||
| ws: wind speed | 1967–2019 | |||
| Punta Arenas2 | 53°7′24″ | pre: precipitation | ||
| (puq2) | 70°52′38″ | tmn: minimum temperature | 1974–2019 | DGA |
| tmx: maximum temperature | ||||
| Ushuaia | 54°48′0″ | pre: precipitation | 1928–2013 | SMN |
| (ush) | 68°18′6″ | tm: mean temperature | 1901–2014 | |
| Puerto Williams | pre: precipitation | 1970–2019 | ||
| (ptw) | 54°55′54″ | tmn: minimum temperature | 1980–2019 | DMC |
| 67°36′36″ | tmx: maximum temperature | 1980–2019 | ||
| ws: wind speed | 1986-2019 | |||
| ERA5 | pre: precipitation | |||
| tmn: minimum temperature | ||||
| T2m: temperature 2 m | ||||
| Reanalysis | WS10m: near surface wind speed | 1979–2019 | [48] | |
| rH: relative humidity | ||||
| MSLP: Sea Level Pressure | ||||
| MZ850: Geopotential Height 850 mb |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soto-Rogel, P.; Aravena, J.C.; Villalba, R.; Bringas, C.; Meier, W.J.-H.; Gonzalez-Reyes, Á.; Grießinger, J. Two Nothofagus Species in Southernmost South America Are Recording Divergent Climate Signals. Forests 2022, 13, 794. https://doi.org/10.3390/f13050794
Soto-Rogel P, Aravena JC, Villalba R, Bringas C, Meier WJ-H, Gonzalez-Reyes Á, Grießinger J. Two Nothofagus Species in Southernmost South America Are Recording Divergent Climate Signals. Forests. 2022; 13(5):794. https://doi.org/10.3390/f13050794
Chicago/Turabian StyleSoto-Rogel, Pamela, Juan Carlos Aravena, Ricardo Villalba, Christian Bringas, Wolfgang Jens-Henrik Meier, Álvaro Gonzalez-Reyes, and Jussi Grießinger. 2022. "Two Nothofagus Species in Southernmost South America Are Recording Divergent Climate Signals" Forests 13, no. 5: 794. https://doi.org/10.3390/f13050794
APA StyleSoto-Rogel, P., Aravena, J. C., Villalba, R., Bringas, C., Meier, W. J.-H., Gonzalez-Reyes, Á., & Grießinger, J. (2022). Two Nothofagus Species in Southernmost South America Are Recording Divergent Climate Signals. Forests, 13(5), 794. https://doi.org/10.3390/f13050794