Growth and Development of Short-Rotation Woody Crops for Rural and Urban Applications


1. Introduction
2. Applications from Around the Globe
3. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McGovern, P.; Kuzovkina, Y.; Soolanayakanahally, R. Short communication: IPC Salix cultivar database proof-of-concept. Forests 2021, 12, 631. [Google Scholar] [CrossRef]
- Fortier, J.; Truax, B.; Gagnon, D.; Lambert, F. Natural drying and chemical characteristics of hybrid poplar firewood produced from agricultural bioenergy buffers in southern Québec, Canada. Forests 2021, 12, 122. [Google Scholar] [CrossRef]
- Zlatković, M.; Tenorio-Baigorria, I.; Lakatos, T.; Tóth, T.; Koltay, A.; Pap, P.; Marković, M.; Orlović, S. Bacterial canker disease on Populus × euramericana caused by Lonsdalea populi in Serbia. Forests 2020, 11, 1080. [Google Scholar] [CrossRef]
- Galović, V.; Kebert, M.; Popović, B.; Kovačević, B.; Vasić, V.; Joseph, M.; Orlović, S.; Szabados, L. Biochemical and gene expression analyses in different poplar clones: The selection tools for afforestation of halomorphic environments. Forests 2021, 12, 636. [Google Scholar] [CrossRef]
- Zalesny, R.; Pilipović, A.; Rogers, E.; Burken, J.; Hallett, R.; Lin, C.; McMahon, B.; Nelson, N.; Wiese, A.; Bauer, E.; et al. Establishment of regional phytoremediation buffer systems for ecological restoration in the Great Lakes Basin, USA. I. Genotype × environment interactions. Forests 2021, 12, 430. [Google Scholar] [CrossRef]
- Pilipović, A.; Zalesny, R.; Rogers, E.; McMahon, B.; Nelson, N.; Burken, J.; Hallett, R.; Lin, C. Establishment of regional phytoremediation buffer systems for ecological restoration in the Great Lakes Basin, USA. II. New clones show exceptional promise. Forests 2021, 12, 474. [Google Scholar] [CrossRef]
- Hu, Y.; Kamelchuk, D.; Krygier, R.; Thomas, B. Field testing of selected salt-tolerant screened balsam poplar (Populus balsamifera L.) clones for use in reclamation around end-pit lakes associated with bitumen extraction in northern Alberta. Forests 2021, 12, 572. [Google Scholar] [CrossRef]
- Han, Q.; Harayama, H.; Uemura, A.; Ito, E.; Utsugi, H.; Kitao, M.; Maruyama, Y. High biomass productivity of short-rotation willow plantation in boreal Hokkaido achieved by mulching and cutback. Forests 2020, 11, 505. [Google Scholar] [CrossRef]
- Harayama, H.; Han, Q.; Ishihara, M.; Kitao, M.; Uemura, A.; Sasaki, S.; Yamada, T.; Utsugi, H.; Maruyama, Y. Estimation of yield loss due to deer browsing in a short rotation coppice willow plantation in northern Japan. Forests 2020, 11, 809. [Google Scholar] [CrossRef]
- Thiffault, N.; Elferjani, R.; Hébert, F.; Paré, D.; Gagné, P. Intensive mechanical site preparation to establish short rotation hybrid poplar plantations—A case-study in Québec, Canada. Forests 2020, 11, 785. [Google Scholar] [CrossRef]
- González, I.; Sixto, H.; Rodríguez-Soalleiro, R.; Oliveira, N. Nutrient contribution of litterfall in a short rotation plantation of pure or mixed plots of Populus alba L. and Robinia pseudoacacia L. Forests 2020, 11, 1133. [Google Scholar] [CrossRef]
- Oliveira, N.; Pérez-Cruzado, C.; Cañellas, I.; Rodríguez-Soalleiro, R.; Sixto, H. Poplar short rotation coppice plantations under Mediterranean conditions: The case of Spain. Forests 2020, 11, 1352. [Google Scholar] [CrossRef]
- Landgraf, D.; Carl, C.; Neupert, M. Biomass yield of 37 different SRC poplar varieties grown on a typical site in northeastern Germany. Forests 2020, 11, 1048. [Google Scholar] [CrossRef]
- Thevs, N.; Fehrenz, S.; Aliev, K.; Emileva, B.; Fazylbekov, R.; Kentbaev, Y.; Qonunov, Y.; Qurbonbekova, Y.; Raissova, N.; Razhapbaev, M.; et al. Growth rates of poplar cultivars across central Asia. Forests 2021, 12, 373. [Google Scholar] [CrossRef]
- Schiberna, E.; Borovics, A.; Benke, A. Economic modelling of poplar short rotation coppice plantations in Hungary. Forests 2021, 12, 623. [Google Scholar] [CrossRef]
- Kraszkiewicz, A. Productivity of black locust (Robinia pseudoacacia L.) grown on varying habitats in southeastern Poland. Forests 2021, 12, 470. [Google Scholar] [CrossRef]
- Ghezehei, S.; Ewald, A.; Hazel, D.; Zalesny, R.; Nichols, E. Productivity and profitability of poplars on fertile and marginal sandy soils under different density and fertilization treatments. Forests 2021, 12, 869. [Google Scholar] [CrossRef]
- Mola-Yudego, B.; Xu, X.; Englund, O.; Dimitriou, I. Reed canary grass for energy in Sweden: Yields, land-use patterns, and climatic profile. Forests 2021, 12, 897. [Google Scholar] [CrossRef]
- Kolozsvári, I.; Kun, Á.; Jancsó, M.; Bakti, B.; Bozán, C.; Gyuricza, C. Utilization of fish farm effluent for irrigation short rotation willow (Salix alba L.) under lysimeter conditions. Forests 2021, 12, 457. [Google Scholar] [CrossRef]
- Istenič, D.; Božič, G. Short-rotation willows as a wastewater treatment plant: Biomass production and the fate of macronutrients and metals. Forests 2021, 12, 554. [Google Scholar] [CrossRef]
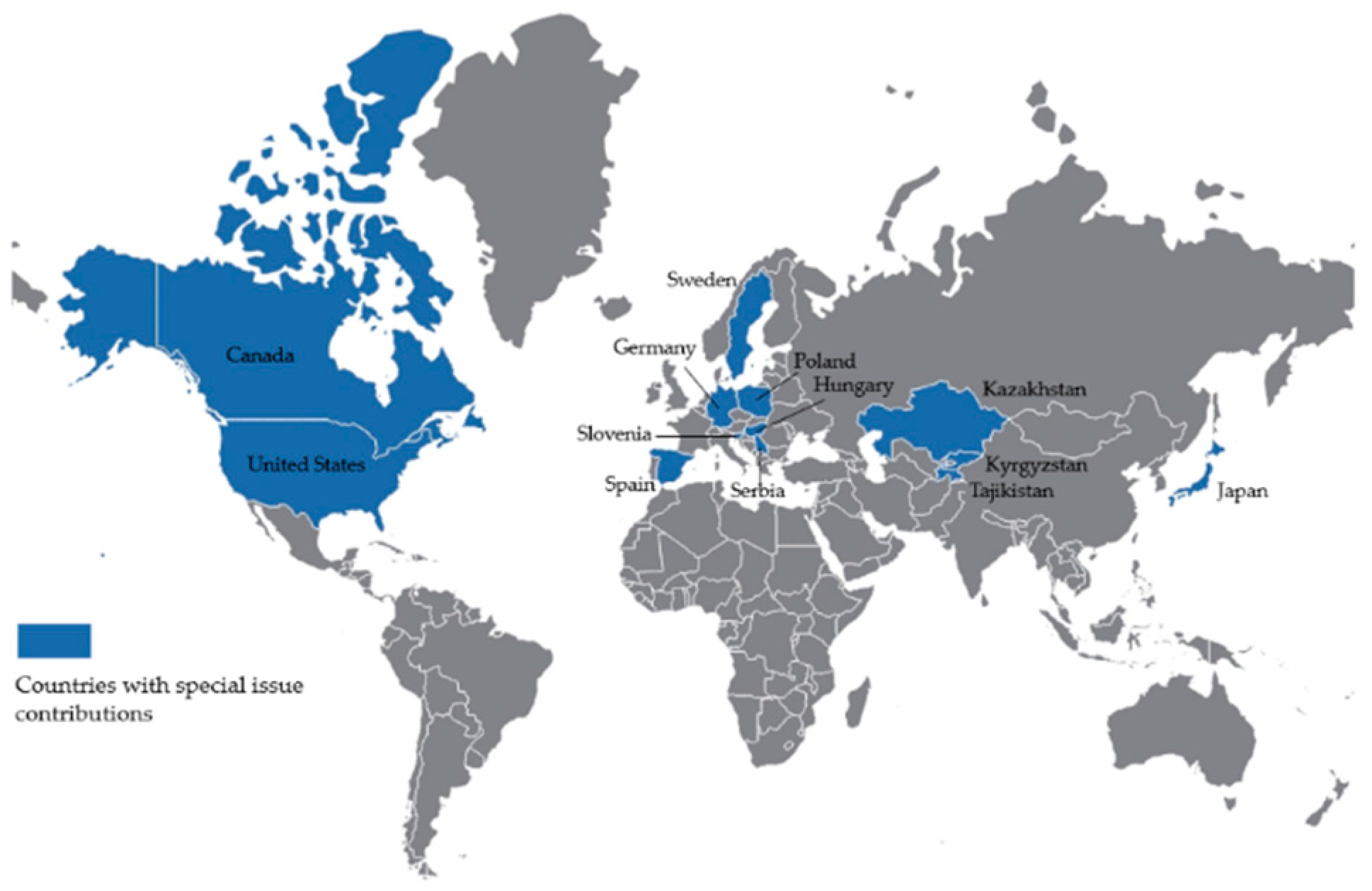


{kind=link}
{kind=link}
| Application | Genus | Location | Contribution | DOI |
|---|---|---|---|---|
| Cultivar Database 1 | Salix | Global | McGovern et al. [1] | https://doi.org/10.3390/f12050631 |
| Forest Buffers | Populus | Canada | Fortier et al. [2] | https://doi.org/10.3390/f12020122 |
| Forest Health Screening | Populus | Serbia | Zlatković et al. [3] | https://doi.org/10.3390/f11101080 |
| Populus | Serbia | Galović et al. [4] | https://doi.org/10.3390/f12050636 | |
| Phytoremediation | Populus | United States | Zalesny et al. [5] | https://doi.org/10.3390/f12040430 |
| Populus | United States | Pilipović et al. [6] | https://doi.org/10.3390/f12040474 | |
| Populus | Canada | Hu et al. [7] | https://doi.org/10.3390/f12050572 | |
| Short Rotation Coppice | Salix | Japan | Han et al. [8] | https://doi.org/10.3390/f11050505 |
| Salix | Japan | Harayama et al. [9] | https://doi.org/10.3390/f11080809 | |
| Populus | Canada | Thiffault et al. [10] | https://doi.org/10.3390/f11070785 | |
| Populus | Spain | González et al. [11] | https://doi.org/10.3390/f11111133 | |
| Robinia | Spain | González et al. [11] | https://doi.org/10.3390/f11111133 | |
| Populus | Spain | Oliveira et al. [12] | https://doi.org/10.3390/f11121352 | |
| Populus | Germany | Landgraf et al. [13] | https://doi.org/10.3390/f11101048 | |
| Populus | Kazakhstan | Thevs et al. [14] | https://doi.org/10.3390/f12030373 | |
| Populus | Kyrgyzstan | Thevs et al. [14] | https://doi.org/10.3390/f12030373 | |
| Populus | Tajikistan | Thevs et al. [14] | https://doi.org/10.3390/f12030373 | |
| Populus | Hungary | Schiberna et al. [15] | https://doi.org/10.3390/f12050623 | |
| Volume Production | Robinia | Poland | Kraszkiewicz [16] | https://doi.org/10.3390/f12040470 |
| Populus | United States | Ghezehei et al. [17] | https://doi.org/10.3390/f12070869 | |
| Phalaris | Sweden | Mola-Yudego et al. [18] | https://doi.org/10.3390/f12070897 | |
| Wastewater Reuse | Salix | Hungary | Kolozsvári et al. [19] | https://doi.org/10.3390/f12040457 |
| Salix | Slovenia | Istenič and Božič [20] | https://doi.org/10.3390/f12050554 |
| Genomic Group 1,2 | Section 3 | Genotype 4 | Contribution |
|---|---|---|---|
| P. alba ‘A’ | Populus | ‘111PK’, ‘Ozolin’ | [11,14] |
| P. balsamifera ‘B’ | Tacamahaca | See [7] | [7] |
| P. deltoides ‘D’ | Aigeiros | ‘7300502’, ‘89M060’, ‘Antonije’, ‘Bora’, ‘Lux’, ‘PE19/66’, ‘Samsun’, ‘Viriato’ | [4,5,6,12,14] |
| P. nigra ‘N’ | Aigeiros | ‘Tr 56/75’, ‘Bordils’, ‘Lombardo Ieones’, ‘Mirza Terek’, ‘Pyramidalis’ | [12,14] |
| P. pamirica ‘P’ | Tacamahaca | Not specified | [14] |
| P. simonii ‘S’ | Tacamahaca | Not specified | [14] |
| P. trichocarpa ‘T’ | Tacamahaca | ‘Fritzi Pauley’, ‘Muhle Larsen’, ‘Trichobel’, ‘Weser 6’ | [13,14] |
| P. alba × P. tremula ‘ATtremula’ | Populus × Populus | ‘4×Göttingen’, ‘P1’ | [13] |
| P. deltoides × P. deltoides ‘DD’ | Aigeiros × Aigeiros | ‘140’, ‘356’ | [17] |
| P. deltoides × P. maximowiczii ‘DM’ | Aigeiros × Tacamahaca | ‘230’, ‘DM114’, ‘NC14106’ | [5,6,17] |
| P. deltoides × P. nigra ‘DN’ | Aigeiros × Aigeiros | ‘9732-11’, ‘9732-24’, ‘9732-31’, ‘9732-36’, ‘99038022’, ‘99059016’, ‘2000 Verde’, ‘AF2’, ‘AF13’, ‘AF15’, ‘AF16’, ‘AF17’, ‘AF18’, ‘AF19’, ‘AF20’, ‘AF24’, ‘AF24’, ‘AF28’, ‘Agathe-F’, ‘BL Constanzo’, ‘Bellini’, ‘Blanc du Poitou’, ‘Branagesi’, ‘B-1M’, ‘Campeador’, ‘Canadá Blanco’, ‘DN2’, ‘DN5’, ‘DN34’, ‘DN177’, ‘Dorskamp’, ‘E-298’, ‘Flevo’, ‘Guardi’, ‘H-8’, ‘H-11’, ‘H-17’, ‘H-33’, ‘H-328’, ‘Harff’, ‘Heidemij’, ‘I-214’, ‘I-45/51’, ‘I-454/40’, ‘Isières’, ‘Jacometti 78 B’, ‘Koltay’, ‘Kopecky’, ‘Kornik-21’, ‘Luisa Avanzo’, ‘MC’, ‘Oudenberg’, ‘Orion’, ‘Pannonia’, ‘Robusta’, ‘Tiepolo’, ‘Triplo’, ‘Veronese’, ‘Vesten’ | [3,4,5,6,12,13,14,15] |
| P. maximowiczii × P. nigra ‘MN’ | Tacamahaca × Aigeiros | ‘Rochester’ | [13] |
| P. maximowiczii × P. trichocarpa ‘MT’ | Tacamahaca × Tacamahaca | ‘Androscoggin’, ‘Fastwood-1’, ‘Fastwood-2’, ‘Matrix-11’, ‘Matrix-24’, ‘Matrix-49’, ‘NE42’ | [13,14] |
| P. nigra × P. maximowiczii ‘NM’ | Aigeiros × Tacamahaca | ‘Max-1’, ‘Max-3’, ‘Max-4’, ‘NM2’, ‘NM5’, ‘NM6’ | [5,6,13,14] |
| P. trichocarpa × P. deltoides ‘TD’ | Tacamahaca × Aigeiros | ‘Beaupre’, ‘Boelare’, ‘Raspalje’, ‘Unal’, ‘185’, ‘49-177’ | [12,17] |
| P. tremula × P. tremuloides ‘TtremulaTtremuloides’ | Populus × Populus | ‘Esch5’ | [13] |
| (P. deltoides × P. nigra) × P. maximowiczii ‘DN×M’ | (Aigeiros × Aigeiros) × Tacamahaca | ‘DN × M-915508’ | [2] |
| P. laurifolia × (P. deltoides × P. nigra) ‘L×DN’ | Tacamahaca × (Aigeiros × Aigeiros) | ‘Kazakhstani’, ‘Kyzyl-Tan’ | [14] |
| (P. maximowiczii × P. deltoides) × P. trichocarpa ‘MD×T’ | (Tacamahaca × Aigeiros) × Tacamahaca | Not specified | [10] |
| P. trichocarpa × (P. trichocarpa × P. deltoides) ‘T×TD’ | Tacamahaca × (Tacamahaca × Aigeiros) | ‘AF8’ | [13] |
| (P. trichocarpa × P. deltoides) × P. alba ‘TD×A’ | (Tacamahaca × Aigeiros) × Populus | ‘I-114/69’ | [12] |
| (P. trichocarpa × P. deltoides) × P. nigra ‘TD×N’ | (Tacamahaca × Aigeiros) × Aigeiros | ‘AF6’, ‘Monviso’ | [13] |
| R. pseudoacacia ‘Probinia’ | na 5 | ‘Nyirsegy’ | [11] |
| S. alba ‘Asalix’ | na | ‘Naperti’, ‘V-160’ | [19,20] |
| S. pet-susu ‘Psalix’ | na | ‘P.C.51’, ‘P.G.12’, ‘P.G.D’, ‘P.I.62’, ‘P.I.81’, ‘P.I.82’, ‘P.T.59’ | [8,9] |
| S. sachalinensis ‘Ssalix’ | na | ‘S.I.27’, ‘S.I.44’, ‘S.I.67’, ‘S.S.3’, ‘S.T.27’ | [8,9] |
| S. alba × S. alba ‘AsalixAsalix’ | na | ‘V-052’, ‘V-093’ | [20] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zalesny, R.S., Jr.; Pilipović, A. Growth and Development of Short-Rotation Woody Crops for Rural and Urban Applications. Forests 2022, 13, 867. https://doi.org/10.3390/f13060867
Zalesny RS Jr., Pilipović A. Growth and Development of Short-Rotation Woody Crops for Rural and Urban Applications. Forests. 2022; 13(6):867. https://doi.org/10.3390/f13060867
Chicago/Turabian StyleZalesny, Ronald S., Jr., and Andrej Pilipović. 2022. "Growth and Development of Short-Rotation Woody Crops for Rural and Urban Applications" Forests 13, no. 6: 867. https://doi.org/10.3390/f13060867