
Changes in the Parasitism Rate and Parasitoid Community Structure of the Horse Chestnut Leafminer, Cameraria ohridella (Lepidoptera: Gracillariidae), in the Czech Republic



Abstract
:1. Introduction
2. Materials and Methods
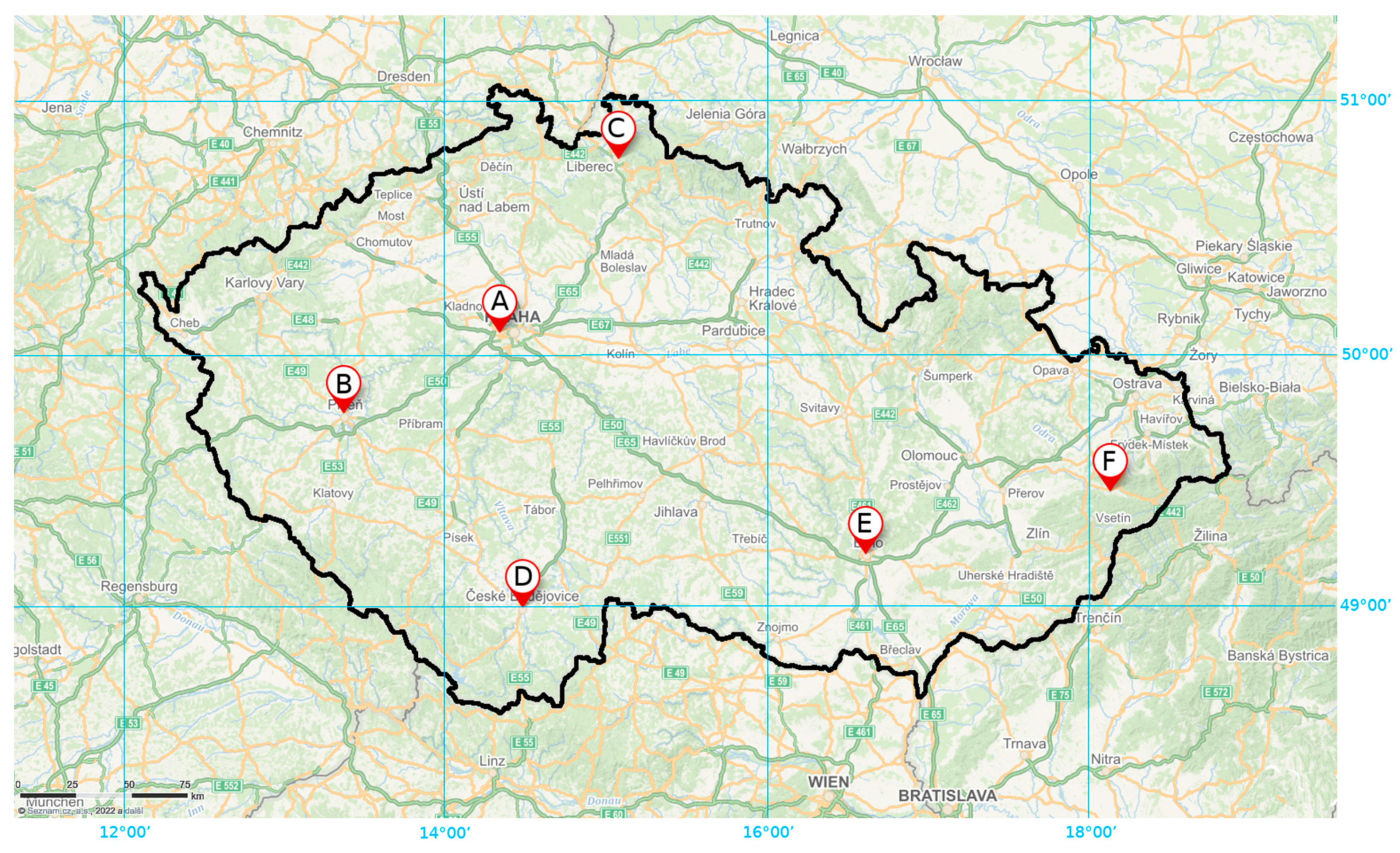
2.1. Sampling Sites
2.2. Leaf Sampling
2.3. Estimation of Population Density of C. ohridella
2.4. Mine Dissection, Rearing of Parasitoids, and Species Determination
2.5. Calculation of Parasitism and Other Mortality Rates
2.6. Statistical Analysis
3. Results
3.1. Dates of the Emergence of the First Generation and Population Density of C. ohridella
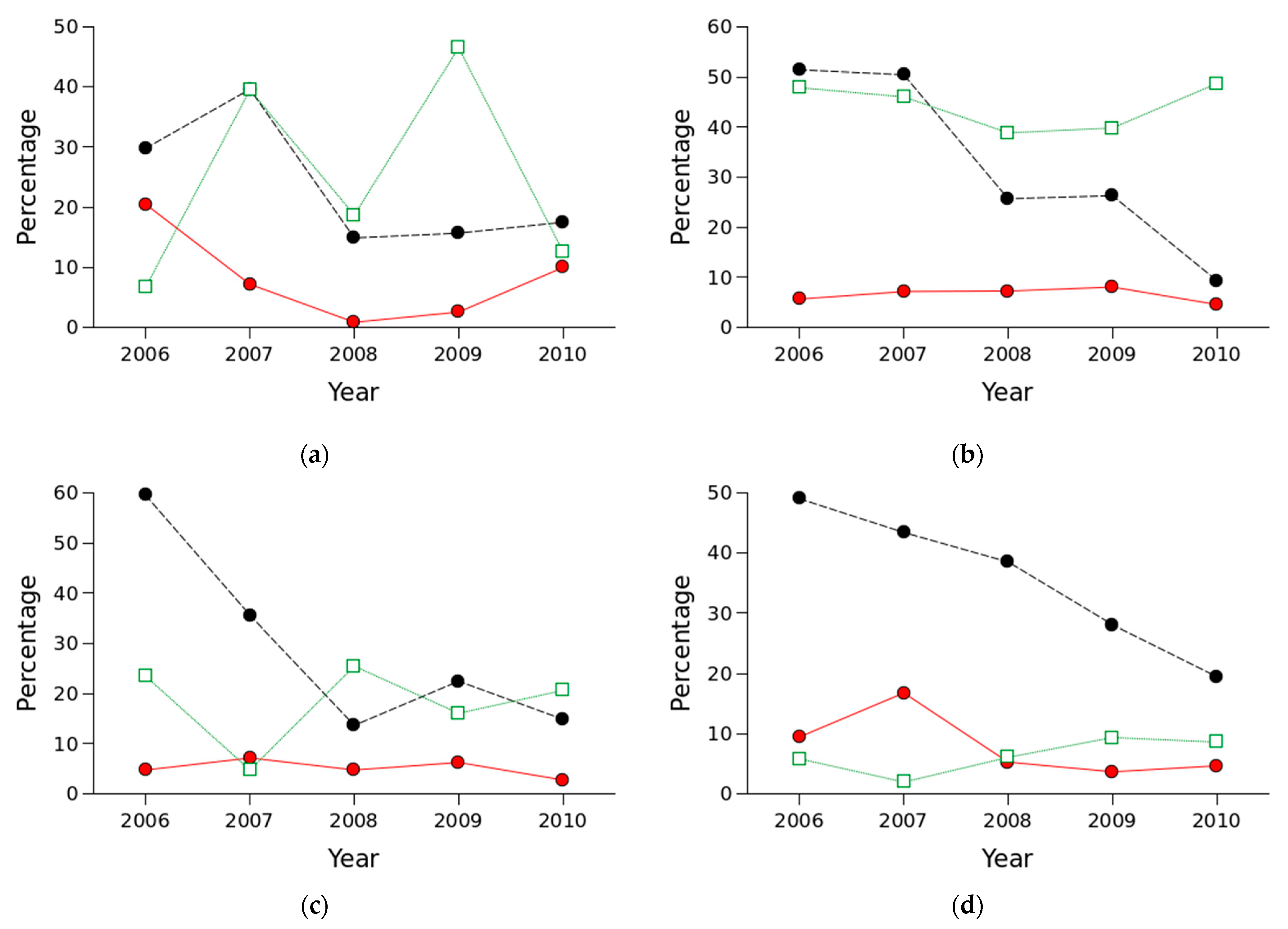
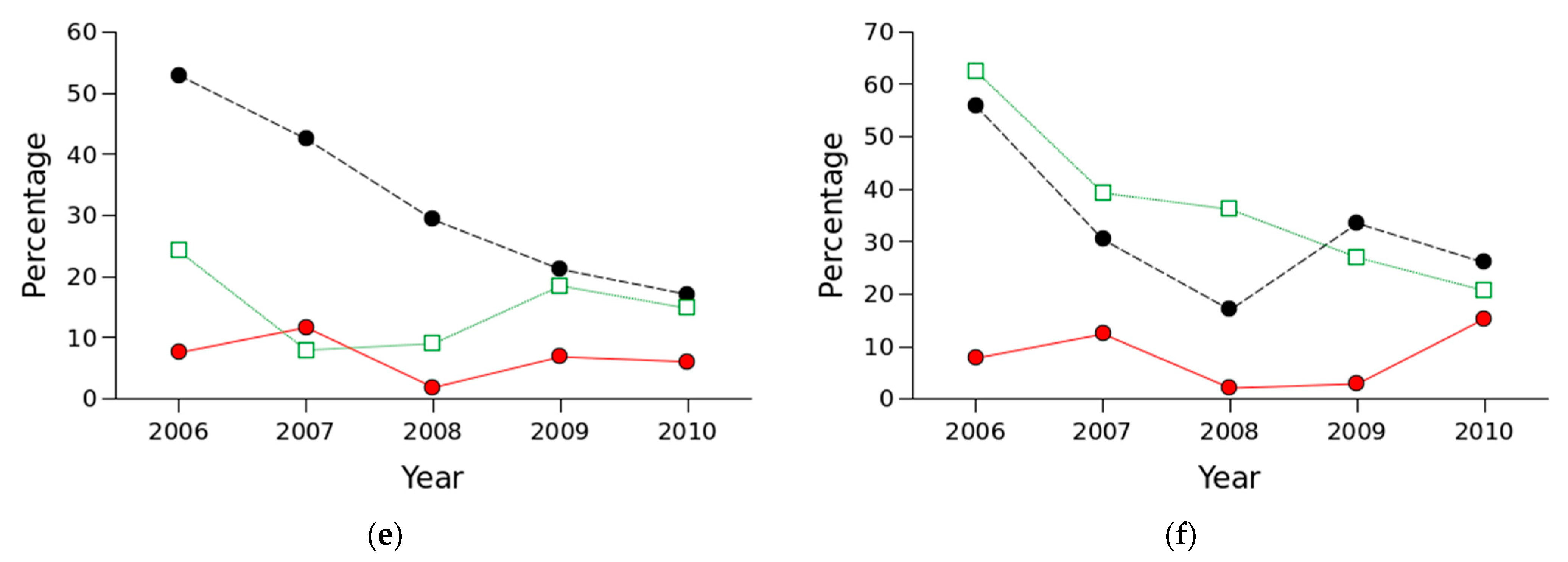
3.2. Parasitism and Other Mortality Rates
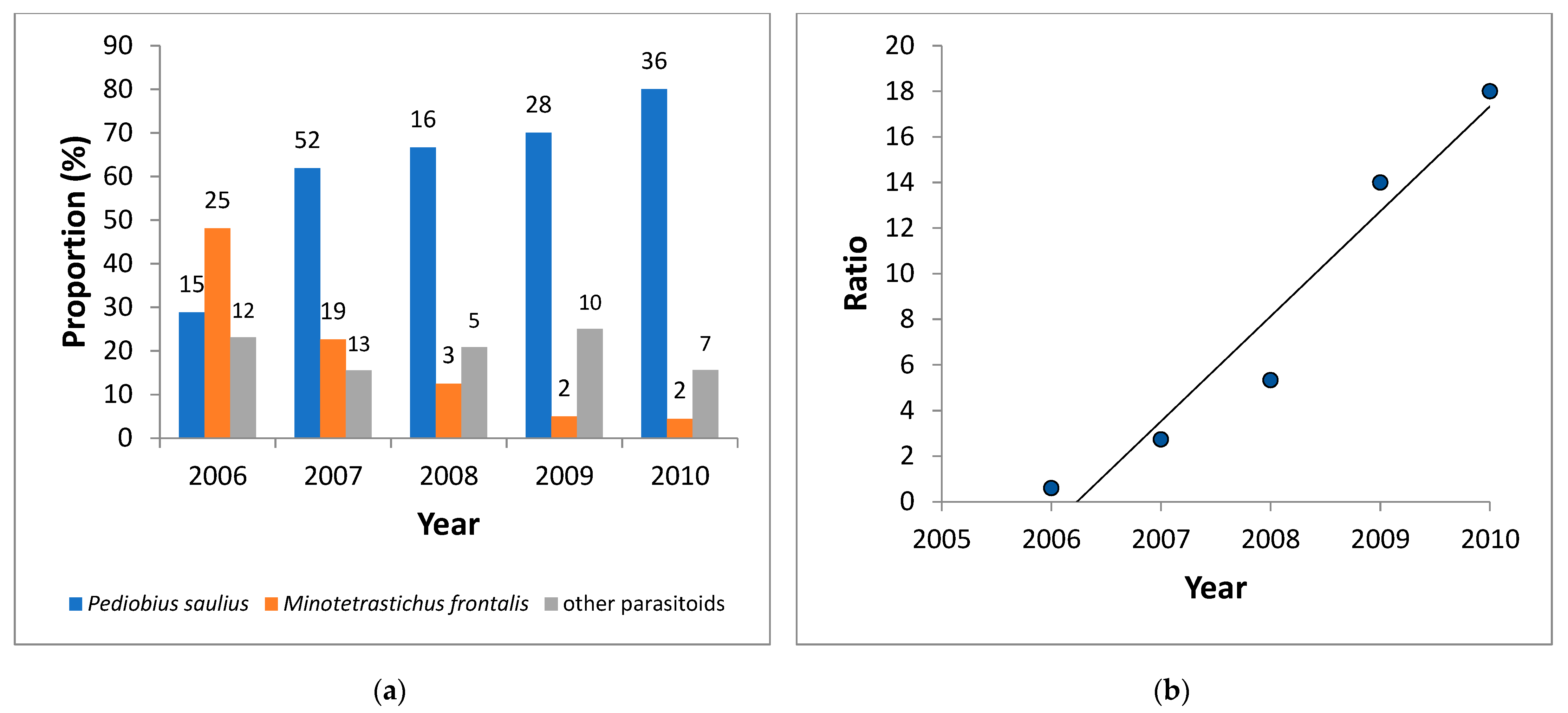
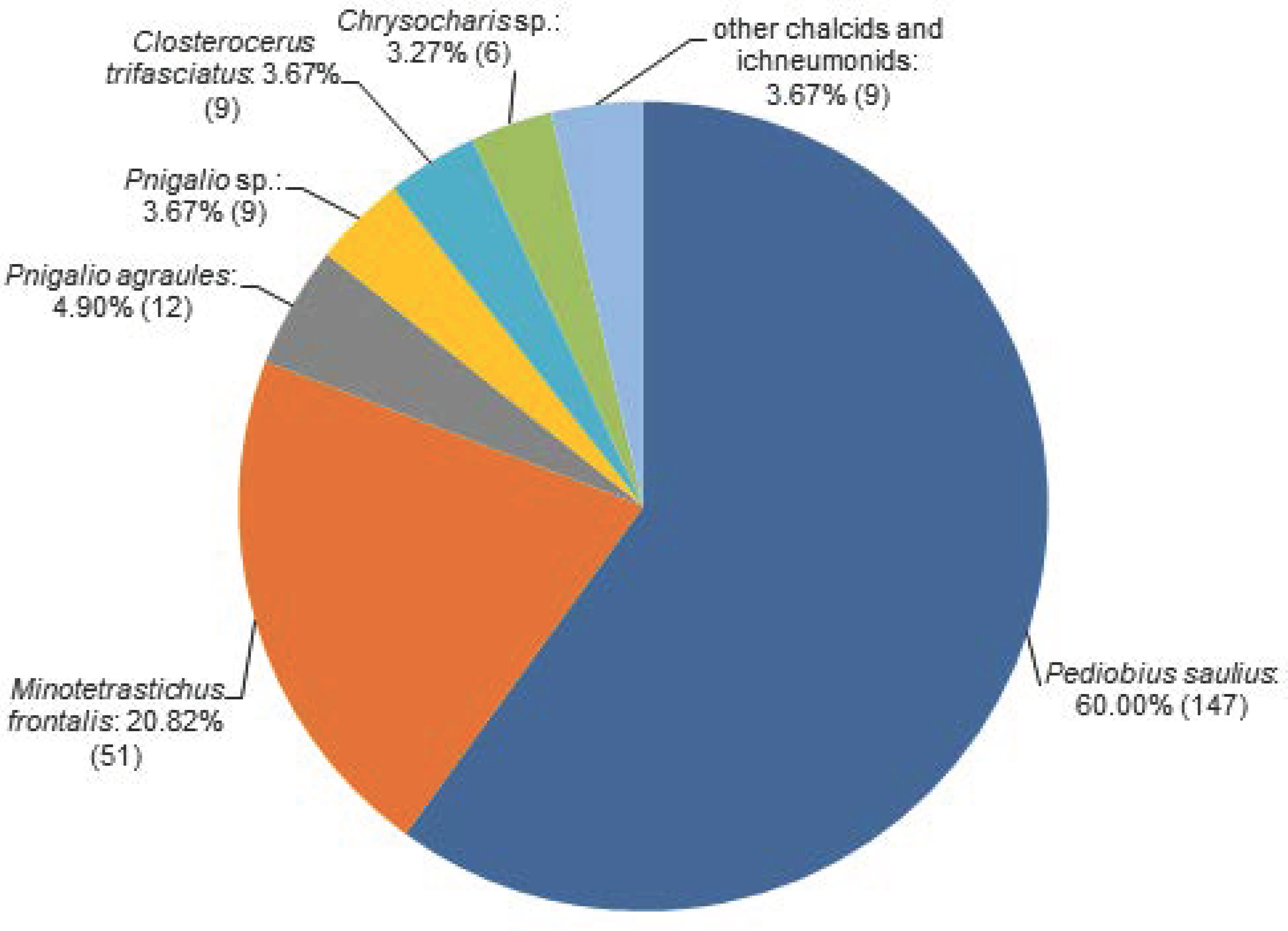
3.3. Parasitoids of C. ohridella
4. Discussion
4.1. Parasitism and Other Mortality Rates
4.2. Parasitoids of C. ohridella
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site 1 | Year | Sampling Date | Parasitism 2 | Other Mortality 2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L1 and L2 | L3 | L4 | SP | P | L1 and L2 | L3 | L4 | SP | P | |||
| A | 2006 | 27 July | 0 | 0 | 0 | 2.9 | 24.1 | 19.6 | 2.9 | 0 | 0 | 10.3 |
| 2007 | 20 June | 1.0 | 0 | 0 | 0.6 | 8.7 | 30.8 | 2.2 | 0.4 | 0 | 10.7 | |
| 2008 | 2 July | 0 | 0 | 0.3 | 0 | 0.8 | 12.2 | 1.2 | 0.8 | 0 | 1.2 | |
| 2009 | 2 July | 0 | 0.2 | 0 | 0 | 2.6 | 9.0 | 0.5 | 0 | 0 | 6.9 | |
| 2010 | 2 July | 0.2 | 0 | 0 | 0.9 | 10.8 | 13.4 | 0 | 0 | 1.8 | 3.1 | |
| B | 2006 | 15 July | 0 | 0.8 | 0.4 | 2.2 | 4.8 | 26.0 | 1.6 | 0.8 | 1.7 | 32.7 |
| 2007 | 17 June | 0 | 0.3 | 0.3 | 3.0 | 7.4 | 28.2 | 4.3 | 0.9 | 1.5 | 27.1 | |
| 2008 | 29 June | 0.2 | 0.5 | 1.0 | 6.4 | 1.8 | 19.8 | 3.2 | 2.7 | 1.7 | 0 | |
| 2009 | 27 June | 0.2 | 0 | 0.7 | 3.8 | 6.1 | 21.0 | 1.7 | 0.3 | 0 | 5.1 | |
| 2010 | 26 June | 0 | 0 | 0 | 0 | 4.9 | 6.0 | 0 | 0 | 0 | 3.7 | |
| C | 2006 | 26 July | 0 | 0 | 0 | 2.9 | 2.9 | 13.2 | 4.1 | 0 | 0 | 52.9 |
| 2007 | 29 June | 0.2 | 0 | 0 | 0.3 | 9.2 | 22.0 | 4.3 | 0.6 | 0 | 13.4 | |
| 2008 | 10 July | 0 | 0 | 0 | 0.3 | 5.1 | 8.6 | 0.4 | 0.7 | 0 | 4.6 | |
| 2009 | 9 July | 0.2 | 0 | 0.3 | 0.4 | 6.6 | 12.6 | 1.9 | 0.3 | 0 | 9.4 | |
| 2010 | 13 July | 0.2 | 0 | 0 | 0.9 | 2.1 | 13.6 | 0.5 | 0 | 0 | 1.1 | |
| D | 2006 | 17 July | 0 | 0.4 | 0.4 | 3.8 | 11.1 | 34.2 | 3.7 | 1.2 | 0 | 19.4 |
| 2007 | 20 June | 1.2 | 0.6 | 0.6 | 4.5 | 17.9 | 25.6 | 4.2 | 1.0 | 0.7 | 20.8 | |
| 2008 | 1 July | 0.2 | 0.2 | 0.6 | 0.7 | 5.4 | 18.2 | 5.2 | 3.9 | 0 | 17.8 | |
| 2009 | 2 July | 0.2 | 0 | 0 | 0.4 | 4.2 | 22.2 | 4.1 | 0 | 0 | 3.6 | |
| 2010 | 2 July | 0.4 | 0 | 0.7 | 1.0 | 3.6 | 15.6 | 2.6 | 0 | 0 | 2.2 | |
| E | 2006 | 10 July | 0 | 0.7 | 1.1 | 5.3 | 3.9 | 26.6 | 1.1 | 0.8 | 1.6 | 36.2 |
| 2007 | 12 June | 0.4 | 0.4 | 0 | 1.8 | 15.6 | 29.4 | 5.8 | 1..9 | 0 | 12.2 | |
| 2008 | 25 June | 0 | 0 | 0.3 | 0.4 | 1.7 | 17.6 | 1.5 | 0.3 | 1.2 | 11.7 | |
| 2009 | 25 June | 0.2 | 0.8 | 0.3 | 0 | 6.9 | 1.4 | 4.3 | 0.3 | 0 | 6.9 | |
| 2010 | 25 June | 0 | 0.2 | 0.4 | 1.5 | 5.1 | 12.4 | 1.7 | 0 | 0 | 3.8 | |
| F | 2006 | 25 July | 0 | 0 | 0 | 0.7 | 10.3 | 26.6 | 1.9 | 0 | 0 | 39 |
| 2007 | 27 June | 0.2 | 0 | 0 | 1.7 | 13.4 | 14 | 2.2 | 2.2 | 0 | 15.6 | |
| 2008 | 7 July | 0 | 0 | 0.3 | 0 | 2.2 | 12.6 | 1.2 | 0 | 0 | 3.9 | |
| 2009 | 8 July | 0.2 | 0 | 0.4 | 1.2 | 2.4 | 28 | 3.1 | 0 | 0 | 4.8 | |
| 2010 | 7 July | 0.6 | 0.3 | 0 | 0.5 | 18.1 | 21 | 0.6 | 0 | 0 | 5.8 | |

References
- Simova-Tosic, D.; Filev, S. Contribution to the Horse Chestnut Miner. Zast. Bilja 1985, 36, 235–239. [Google Scholar]
- Grabenweger, G.; Grill, R. On the Place of Origin of Cameraria Ohridella Deschka & Dimic (Lepidoptera: Gracillariidae). Beiträge Zur Entomofaunist. 2000, 1, 9–17. [Google Scholar]
- Valade, R.; Kenis, M.; Hernandez-Lopez, A.; Augustin, S.; Mari Mena, N.; Magnoux, E.; Rougerie, R.; Lakatos, F.; Roques, A.; Lopez-Vaamonde, C. Mitochondrial and Microsatellite DNA Markers Reveal a Balkan Origin for the Highly Invasive Horse-Chestnut Leaf Miner Cameraria Ohridella (Lepidoptera, Gracillariidae). Mol. Ecol. 2009, 18, 3458–3470. [Google Scholar] [CrossRef]
- Thalmann, C.; Freise, J.; Heitland, W.; Bacher, S. Effects of Defoliation by Horse Chestnut Leafminer (Cameraria Ohridella) on Reproduction in Aesculus hippocastanum. Trees-Struct. Funct. 2003, 17, 383–388. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.A.; Alhamd, O.; Iszkuło, G.; Dering, M.; Mukassabi, T.A. Biological Flora of the British Isles: Aesculus hippocastanum. J. Ecol. 2019, 107, 992–1030. [Google Scholar] [CrossRef] [Green Version]
- Liška, J. Verbreitung der Rosskastanienminiermotte in Tschechien; Forstschutz-Aktuell: Wien, Austria, 1997; Volume 21. [Google Scholar]
- Šefrová, H.; Laštůvka, Z. Dispersal of the Horse-Chestnut Leafminer, Cameraria Ohridella Deschka & Dimic, 1986, in Europe: Its Course, Ways and Causes (Lepidoptera: Gracillariidae). Entomol. Z. 2001, 111, 194–198. [Google Scholar]
- Cebeci, H.; Acer, S. The Occurrence of Some Lepidopterous Species on the Horse Chestnut (Aesculus hippocastanum L.) at Istanbul-Belgrad Forest in Turkey. Acta Agric. Slov. 2007, 89, 95–102. [Google Scholar] [CrossRef]
- Laštůvka, Z.; Liška, J.; Vávra, J.; Elsner, V.; Laštůvka, A.; Marek, J.; Dufek, T.; Dvořák, M.; Kopeček, F.; Petrů, M.; et al. Faunistic Records from the Czech Republic. Klapalekiana 1994, 18, 197–206. [Google Scholar]
- Wittenberger, G. Die Roßkastanien-Miniermotte in Nordböhmen nebst einigen Hinweisen zum Vorkommen in Schlesien (Polen). Nassau. Ver. Für Nat. 1998, 98, 75–78. [Google Scholar]
- D’Costa, L.; Koricheva, J.; Straw, N.; Simmonds, M.S.J. Oviposition Patterns and Larval Damage by the Invasive Horse-Chestnut Leaf Miner Cameraria Ohridella on Different Species of Aesculus: Oviposition Patterns and Larval Damage. Ecol. Entomol. 2013, 38, 456–462. [Google Scholar] [CrossRef]
- Skuhravý, V. Zur Kenntnis der Blattminen-Motte Cameraria ohridella Desch. & Dim. (Lep., Lithocolletidae) an Aesculus hippocastanum L. in der Tschenchischen Republik. Anz. Schadl. Pflanzenschutz Umweltschutz 1998, 71, 81–84. [Google Scholar] [CrossRef]
- Péré, C.; Augustin, S.; Turlings, T.C.J.; Kenis, M. The Invasive Alien Leaf Miner Cameraria Ohridella and the Native Tree Acer Pseudoplatanus: A Fatal Attraction? Agric. For. Entomol. 2010, 12, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Kopacka, M.; Zemek, R. Spatial Variability in the Level of Infestation of the Leaves of Horse Chestnut by the Horse Chestnut Leaf Miner, Cameraria Ohridella (Lepidoptera: Gracillariidae) and in the Number of Adult Moths and Parasitoids Emerging from Leaf Litter in an Urban Environment. Eur. J. Entomol. 2017, 114, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Kopačka, M.; Nachman, G.; Zemek, R. Seasonal Changes and the Interaction between the Horse Chestnut Leaf Miner Cameraria ohridella and Horse Chestnut Leaf Blotch Disease Caused by Guignardia aesculi. Forests 2021, 12, 952. [Google Scholar] [CrossRef]
- Nardini, A.; Raimondo, F.; Scimone, M.; Salleo, S. Impact of the Leaf Miner Cameraria Ohridella on Whole-Plant Photosynthetic Productivity of Aesculus hippocastanum: Insights from a Model. Trees 2004, 18, 714–721. [Google Scholar] [CrossRef]
- Salleo, S.; Nardini, A.; Raimondo, F.; Lo Gullo, M.A.; Pace, F.; Giacomich, P. Effects of Defoliation Caused by the Leaf Miner Cameraria Ohridella on Wood Production and Efficiency in Aesculus hippocastanum Growing in North-Eastern Italy. Trees 2003, 17, 367–375. [Google Scholar] [CrossRef]
- Freise, J.F.; Heitland, W. The Waipuna Hot Foam System–a Chance for Cameraria Ohridella Control? Nachrichtenbl. Deut. Pflanzenschutz 2003, 55, 242–244. [Google Scholar]
- Heitland, W.; Kopelke, J.-P.; Freise, J. Die Roßkastanien-Miniermotte–19 Jahre Forschung Und Noch Keine Lösung in Sicht. Nat. Und Mus. 2003, 133, 221–231. [Google Scholar]
- Kehrli, P.; Bacher, S. Date of Leaf Litter Removal to Prevent Emergence of Cameraria Ohridella in the Following Spring. Entomol. Exp. Appl. 2003, 107, 159–162. [Google Scholar] [CrossRef]
- Kehrli, P.; Lehmann, M.; Bacher, S. Mass-Emergence Devices: A Biocontrol Technique for Conservation and Augmentation of Parasitoids. Biol. Control 2005, 32, 191–199. [Google Scholar] [CrossRef]
- Pavela, R.; Bárnet, M. Systemic Applications of Neem in the Control of Cameraria Ohridella, a Pest of Horse Chestnut (Aesculus hippocastanum). Phytoparasitica 2005, 33, 49–56. [Google Scholar] [CrossRef]
- Zemek, R.; Prenerová, E.; Volter, L.; Weyda, F.; Skuhravý, V. Perspectives for the Biological Control of Cameraria ohridella. Commun. Agric. Appl. Biol. Sci. 2007, 72, 521–526. [Google Scholar] [PubMed]
- Prenerová, E.; Zemek, R.; Volter, L.; Weyda, F. Strain of Entomopathogenic Fungus Isaria Fumosorosea CCM 8367 (CCEFO.011.PFR) and the Method for Controlling Insect and Mite Pests. EPO Patent No. EP2313488, 29 April 2015. [Google Scholar]
- Barta, M. In Planta Bioassay on the Effects of Endophytic Beauveria Strains against Larvae of Horse-Chestnut Leaf Miner (Cameraria Ohridella). Biol. Control 2018, 121, 88–98. [Google Scholar] [CrossRef]
- Richter, D.; Sermann, H.; Jackel, B.; Buttner, C.B. Pathogenicity of Entomopathogenic Fungi on Hibernating Pupae of Cameraria Ohridella Deschka & Dimic 1986 (Lepidoptera, Gracillariidae). Part 1: Pathogenicity against the Naked Pupa. Commun. Agric. Appl. Biol. Sci. 2007, 72, 399–410. [Google Scholar] [PubMed]
- Richter, D.; Sermann, H.; Jackel, B.; Buttner, C. Pathogenicity of Entomopathogenic Fungi on Hibernating Pupae of Cameraria Ohridella Deschka & Dimic 1986 (Lepidoptera, Gracillariidae). Part 2: Efficacy of Entomopathogenic Fungi against Pupa in Her Pupal Cell. Commun. Agric. Appl. Biol. Sci. 2007, 72, 411–421. [Google Scholar]
- Kalmus, M.; Sermann, H.; Buettner, C. Efficacy of Entomopathogenic Fungi against Eggs and Larvae of the Horse Chestnut Leafminer Cameraria ohridella. J. Plant Dis. Prot. 2007, 114, 92. [Google Scholar]
- Nedveckytė, I.; Pečiulytė, D.; Būda, V. Fungi Associated with Horse-Chestnut Leaf Miner Moth Cameraria Ohridella Mortality. Forests 2021, 12, 58. [Google Scholar] [CrossRef]
- Robert, S.; Petra, C.; Juraj, M.; Marek, B. Natural Prevalence of Entomopathogenic Fungi in Hibernating Pupae of Cameraria Ohridella (Lepidoptera: Gracillariidae) and Virulence of Selected Isolates. Plant Protect. Sci. 2016, 52, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Matuska-Lyzwa, J.; Kaca, W.; Zarnowiec, P. Biological Activity of Wild Isolates of Entomopathogenic Nematodes to Horse-Chestnut Leaf Miner (Cameraria Ohridella). Pol. J. Environ. Stud. 2015, 24, 1181–1184. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Agassiz, D.J.L.; Nash, D.R.; Lawton, J.H. The Recruitment of Parasitoid Species to Two Invading Herbivores. J. Anim. Ecol. 1995, 64, 393. [Google Scholar] [CrossRef]
- Kirichenko, N.; Augustin, S.; Kenis, M. Invasive Leafminers on Woody Plants: A Global Review of Pathways, Impact, and Management. J. Pest Sci. 2019, 92, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Grabenweger, G.; Kehrli, P.; Schlick-Steiner, B.; Steiner, F.; Stolz, M.; Bacher, S. Predator Complex of the Horse Chestnut Leafminer Cameraria Ohridella: Identification and Impact Assessment. J. Appl. Entomol. 2005, 129, 353–362. [Google Scholar] [CrossRef]
- Zemek, R.; Prenerova, E.; Weyda, F. The First Record of Entomopathogenic Fungus Paecilomyces Fumosoroseus (Deuteromycota: Hyphomycetes) on the Hibernating Pupae of Cameraria Ohridella (Lepidoptera: Gracillariidae). Entomol. Res. 2007, 37, A135–A136. [Google Scholar]
- Grabenweger, G.; Kehrli, P.; Zweimüller, I.; Augustin, S.; Avtzis, N.; Bacher, S.; Freise, J.; Girardoz, S.; Guichard, S.; Heitland, W.; et al. Temporal and Spatial Variations in the Parasitoid Complex of the Horse Chestnut Leafminer during Its Invasion of Europe. Biol. Invasions 2010, 12, 2797–2813. [Google Scholar] [CrossRef] [Green Version]
- Lupi, D. A 3 Year Field Survey of the Natural Enemies of the Horse-Chestnut Leaf Miner Cameraria Ohridella in Lombardy, Italy. Biocontrol 2005, 50, 113–126. [Google Scholar] [CrossRef]
- Girardoz, S.; Kenis, M.; Quicke, D.L.J. Recruitment of Native Parasitoids by an Exotic Leaf Miner, Cameraria Ohridella: Host-Parasitoid Synchronization and Influence of the Environment. Agric. Forest Entomol. 2006, 8, 49–56. [Google Scholar] [CrossRef]
- Tóth, P.; Lukáš, J. Parasitic Ichneumonoidea on the Horse Chestnut Leaf Miner, Cameraria Ohridella (Lepidoptera: Gracillariidae) in Slovakia. J. Pest Sci. 2005, 78, 151–154. [Google Scholar] [CrossRef]
- Grabenweger, G.; Avtzis, N.; Girardoz, S.; Hrasovec, B.; Tomov, R.; Kenis, M. Parasitism of Cameraria Ohridella (Lepidoptera, Gracillariidae) in Natural and Artificial Horse-Chestnut Stands in the Balkans. Agric. Forest Entomol. 2005, 7, 291–296. [Google Scholar] [CrossRef]
- Freise, J.F.; Heitland, W.; Tosevski, I. Parasitism of the Horse Chestnut Leaf Miner, Cameraria Ohridella Deschka and Dimic (Lep., Gracillariidae), in Serbia and Macedonia. Anz Schadl. 2002, 75, 152–157. [Google Scholar] [CrossRef]
- Girardoz, S.; Volter, L.; Tomov, R.; Quicke, D.L.J.; Kenis, M. Variations in Parasitism in Sympatric Populations of Three Invasive Leaf Miners. J. Appl. Entomol. 2007, 131, 603–612. [Google Scholar] [CrossRef]
- Grabenweger, G.; Lethmayer, C. Occurrence and Phenology of Parasitic Chalcidoidea on the Horse Chestnut Leafminer, Cameraria Ohridella Deschka & Dimic (Lep., Gracillariidae). J. Appl. Entomol. 1999, 123, 257–260. [Google Scholar] [CrossRef]
- Grabenweger, G. Parasitism of Different Larval Stages of Cameraria ohridella. BioControl 2003, 48, 671–684. [Google Scholar] [CrossRef]
- Marchesini, E.; Pasini, M.; Galbero, G. Parassitoidi Della Minatrice Fogliare Degli Ippocastani. L’informatore Agrar. 2002, 27, 75–77. [Google Scholar]
- Stojanović, A.; Marković, C. Parasitoid Complex of Cameraria Ohridella (Lepidoptera: Gracillariidae) in Serbia. Phytoparasitica 2004, 32, 132–140. [Google Scholar] [CrossRef]
- Volter, L.; Kenis, M. Parasitoid Complex and Parasitism Rates of the Horse Chestnut Leafminer, Cameraria Ohridella (Lepidoptera: Gracillariidae) in the Czech Republic, Slovakia and Slovenia. Eur. J. Entomol. 2006, 103, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Van Driesche, R.G. Meaning of “Percent Parasitism” in Studies of Insect Parasitoids. Environ. Entomol. 1983, 12, 1611–1622. [Google Scholar] [CrossRef]
- Freise, J.F.; Heitland, W. Parasitierungsraten: Ermittlung Und Interpretation Am Fallbeispiel Der Roßkastanien-Miniermotte, Cameraria Ohridella, Einer Invasiven Und Faunenfremden Schmetterlingsart in Europa. Mitt. Dtsch. Ges. Allg. Angew. Ent. 2004, 14, 347–350. [Google Scholar]
- Moreth, L.; Baur, H.; Schönitzer, K.; Diller, E. Zum Parasitoid-Komplex Der Roßkastanien-Miniermotte in Bayern (Cameraria Ohridella, Gracillariidae, Lithocolletinae). Mitt. Dtsch. Ges. Allg. Angew. Ent. 2000, 12, 489–492. [Google Scholar]
- Stolz, M. Untersuchungen Zur Befallsregulierung Der Kastanienminiermotte Durch Natürliche Gegenspieler. Der Förderungsdienst 2000, 48, 193–195. [Google Scholar]
- Nováková, P.; Holuša, J.; Horák, J. The Role of Geography and Host Abundance in the Distribution of Parasitoids of an Alien Pest. PeerJ 2016, 4, e1592. [Google Scholar] [CrossRef] [Green Version]
- Lomolino, M.V.; Riddle, B.R.; Brown, J.H.; Brown, J.H. Biogeography, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2006; ISBN 978-0-87893-062-3. [Google Scholar]
- Myśkow, E.; Sokołowska, K.; Słupianek, A.; Gryc, V. Description of Intra-Annual Changes in Cambial Activity and Differentiation of Secondary Conductive Tissues of Aesculus hippocastanum Trees Affected by the Leaf Miner Cameraria ohridella. Forests 2021, 12, 1537. [Google Scholar] [CrossRef]
- Gilbert, M.; Gregoire, J.-C. Visual, Semi-Quantitative Assessments Allow Accurate Estimates of Leafminer Population Densities: An Example Comparing Image Processing and Visual Evaluation of Damage by the Horse Chestnut Leafminer Cameraria Ohridella (Lep., Gracillariidae). J. Appl. Entomol. 2003, 127, 354–359. [Google Scholar] [CrossRef]
- Grabenweger, G.; Stolz, M.; Jeziorny, C. A Key to the Parasitoids of Cameraria Ohridella (Lep., Gracillariidae); CD-ROM v. 1.0; Verlag Naturhistorisches Museum: Vienna, Austria, 2003. [Google Scholar]
- SAS Institute. SAS/STAT 14.3: User’s Guide; SAS Institute: Cary, NC, USA, 2017. [Google Scholar]
- Liang, K.-Y.; Zeger, S.L. Longitudinal Data Analysis Using Generalized Linear Models. Biometrika 1986, 73, 13–22. [Google Scholar] [CrossRef]
- Siegel, S.; Castellan, N.J. Nonparametric Statistics for the Behavioral Sciences, 2nd ed.; McGraw-Hill: Boston, MA, USA, 2003; ISBN 978-0-07-057357-4. [Google Scholar]
- Cebeci, H.; Markovic, C.; Grabenweger, G.; Ayberk, H.; Dobrosavljevic, J.; Goltas, M.; Stojanovic, A. Preliminary Notes on Pupal Parasitism Rates of the Horse Chestnut Leafminer, Cameraria Ohridella (Lepidoptera: Gracillariidae) in Belgrade and Istanbul. Fresenius Environ. Bull. 2018, 27, 7122–7124. [Google Scholar]
- Baraniak, E.; Walczak, U.; Tryjanowski, P.; Zduniak, P. Effect of Distance between Host Trees and Leaf Litter Removal on Population Density of Cameraria Ohridella Deschka & Dimic, 1986 (Lepidoptera, Gracillariidae)-Pest of Chestnut (Aesculus Sp.) Trees. Pol. J. Ecol. 2004, 52, 569–574. [Google Scholar]
- Klug, T.; Meyhöfer, R.; Kreye, M.; Hommes, M. Native Parasitoids and Their Potential to Control the Invasive Leafminer, Cameraria Ohridella DESCH. & DIM. (Lep.: Gracillariidae). Bull. Entomol. Res. 2008, 98, 379–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balázs, K.; Thuróczy, C. Über Den Parasitoidkomplex von Cameraria Ohridella Deschka et Dimi ü 1986, (Lepidop-Tera, Lithocolletidae). Entomol. Basil. 2000, 22, 269–277. [Google Scholar]
- Balázs, K.; Pál, B. Parasitoidender Roßkastanien-Miniermotte (Cameraria Ohridella Deschka & Dimić, 1986) in Nordwest-Ungarn. Acta Agron. Ovar. 2001, 43, 35–48. [Google Scholar]
- Čapek, M. Parasitoids of the Horse-Chestnut Leafminer. In Horse-Chestnut Leafminer (Cameraria ohridella Deschka & Dimić, 1986); Laštůvka, Z., Ed.; Veronica: Brno, Czech Republic, 1999; pp. 1–12. [Google Scholar]
- Rämert, B.; Kenis, M.; Kärnestam, E.; Nyström, M.; Rännbäck, L.-M. Host Plant Suitability, Population Dynamics and Parasitoids of the Horse Chestnut Leafminer Cameraria Ohridella (Lepidoptera: Gracillariidae) in Southern Sweden. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2011, 61, 480–486. [Google Scholar] [CrossRef]
- Noyes, J. Interactive Catalogue of World Chalcidoidea. CD-ROM; Taxapad: Vancouver, BC, Canada, 2002. [Google Scholar]
- Gebiola, M.; Lopez-Vaamonde, C.; Nappo, A.G.; Bernardo, U. Did the Parasitoid Pnigalio Mediterraneus (Hymenoptera: Eulophidae) Track the Invasion of the Horse Chestnut Leafminer? Biol. Invasions 2014, 16, 843–857. [Google Scholar] [CrossRef]




| Label | City | Local Name | Geographical Coordinates | Altitude(m) | Greenery | ||
|---|---|---|---|---|---|---|---|
| Type | Area 1 | Care 2 | |||||
| A | Praha | Obora Hvězda | 50°4′57″ N, 14°19′54″ E | 369 | city park | 88 | 2 |
| B | Plzeň | Lochotínský park | 49°45′37″ N, 13°21′49″ E | 310 | city park | 36 | 1 |
| C | Liberec | Jablonecká street | 50°46′5″ N, 15°4′4″ E | 382 | group of trees | 2 | 1 |
| D | České Budějovice | Sady | 48°58′38″ N, 14°28′34″ E | 386 | city park | 3 | 2 |
| E | Brno | Park Špilberk | 49°11′40″ N, 16°36′12″ E | 240 | city park | 21 | 1 |
| F | Rožnov pod Radhoštěm | Hradisko | 49°26′53″ N, 18°7′9″ E | 463 | alley | 166 | 0 |
| Dependent Variable | Parameter | Estimate | Standard Error | Z | p |
|---|---|---|---|---|---|
| Parasitism rate | Intercept | 440.340 | 136.875 | 3.22 | 0.001 |
| Year | −0.215 | 0.069 | −3.13 | 0.002 | |
| Altitude | 0.001 | 0.001 | 1.41 | 0.157 | |
| Latitude | −0.331 | 0.052 | −6.32 | <0.001 | |
| Greenery area | −0.002 | 0.002 | −1.00 | 0.319 | |
| Developmental stage | 1.165 | 0.091 | 12.81 | <0.001 | |
| C. ohridella population density | −0.009 | 0.007 | −1.15 | 0.248 | |
| Care (mowing, raking) intensity | −0.157 | 0.047 | −3.31 | 0.001 | |
| Other mortality rate | Intercept | 605.940 | 73.252 | 8.27 | <0.001 |
| Year | −0.295 | 0.037 | −8.06 | <0.001 | |
| Altitude | 0.001 | 0.000 | 1.65 | 0.099 | |
| Latitude | −0.315 | 0.019 | −16.80 | <0.001 | |
| Greenery area | −0.001 | 0.001 | −2.36 | 0.019 | |
| Developmental stage | −0.515 | 0.053 | −9.82 | <0.001 | |
| C. ohridella population density | 0.001 | 0.002 | 0.44 | 0.662 | |
| Care (mowing, raking) intensity | −0.059 | 0.015 | −4.06 | <0.001 |
| Superfamily | Family | Species | Sampling Site 1 | Host Stage 2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | F | L1 and L2 | L3 | L4 | SP | P | |||
| Chalcidoidea | Eulophidae | Cirrospilus vittatus (Walker) | 1 | 1 | |||||||||
| Closterocerus trifasciatus (Westwood) | 1 | 4 | 2 | 2 | 1 | 2 | 6 | ||||||
| Chrysocharis sp. | 1 | 2 | 4 | 1 | 1 | 4 | 3 | ||||||
| Minotetrastichus frontalis (Nees) | 3 | 6 | 1 | 19 | 10 | 12 | 1 | 22 | 28 | ||||
| Pediobius saulius (Walker) | 11 | 28 | 23 | 26 | 19 | 40 | 1 | 146 | |||||
| Pnigalio agraules (Walker) | 7 | 2 | 2 | 1 | 1 | 3 | 5 | 3 | |||||
| Pnigalio sp. | 3 | 6 | 1 | 2 | 5 | 1 | |||||||
| Eupelmidae | Eupelmus urozonus (Dalman) | 1 | 1 | ||||||||||
| Pteromalidae | Pteromalus sp. | 3 | 1 | 2 | |||||||||
| Ichneumonoidea | Ichneumonidae | Itoplectis alternans (Gravenhorst) | 1 | 1 | |||||||||
| Scambus annulatus (Kiss) | 2 | 1 | 3 | ||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volter, L.; Prenerová, E.; Weyda, F.; Zemek, R. Changes in the Parasitism Rate and Parasitoid Community Structure of the Horse Chestnut Leafminer, Cameraria ohridella (Lepidoptera: Gracillariidae), in the Czech Republic. Forests 2022, 13, 885. https://doi.org/10.3390/f13060885
Volter L, Prenerová E, Weyda F, Zemek R. Changes in the Parasitism Rate and Parasitoid Community Structure of the Horse Chestnut Leafminer, Cameraria ohridella (Lepidoptera: Gracillariidae), in the Czech Republic. Forests. 2022; 13(6):885. https://doi.org/10.3390/f13060885
Chicago/Turabian StyleVolter, Lubomír, Eva Prenerová, František Weyda, and Rostislav Zemek. 2022. "Changes in the Parasitism Rate and Parasitoid Community Structure of the Horse Chestnut Leafminer, Cameraria ohridella (Lepidoptera: Gracillariidae), in the Czech Republic" Forests 13, no. 6: 885. https://doi.org/10.3390/f13060885
APA StyleVolter, L., Prenerová, E., Weyda, F., & Zemek, R. (2022). Changes in the Parasitism Rate and Parasitoid Community Structure of the Horse Chestnut Leafminer, Cameraria ohridella (Lepidoptera: Gracillariidae), in the Czech Republic. Forests, 13(6), 885. https://doi.org/10.3390/f13060885