Abstract
Camellia oleifera (Abel) is an economic tree species and one of the four largest oil plants in the world. The leaf and twig responses and plasticity indices of C. oleifera were investigated under four light regimes in Pinus massoniana understory plantations, namely, 100% light intensity (CK), 75% of CK (HL), 50% of CK (ML), and 30% of CK (LL). The morphological characteristics, biomass allocation, and physiological characteristics of C. oleifera leaves and twigs under different light regimes, as well as their plasticity indexes, were comprehensively evaluated. The results showed that leaf area, and specific leaf area, leaf total carbon, total nitrogen, total phosphorus and chlorophyll contents, and photosynthesis increased, which indicates that plants have the strongest adaptability under HL. No fruit appeared in twigs under LL and ML. The plastic morphological traits were greater than the biomass allocation and physiological traits. The plasticity of palisade/sponge tissue thickness and lower epidermis thickness were the lowest. In conclusion, C. oleifera have differences in sensitivity and regulation mechanism according to their differences in leaf morphological characteristics, biomass allocation physiological indicators, and response to light regimes. C. oleifera plants showed obvious phenotypic inhibition under CK, while they can adjust their strategies for using light energy to maintain their own growth and development under HL. The wide range of light adaptation and strong plasticity of C. oleifera may be two important reasons for its existence in heterogeneous habitats, but it needs at least 75% light regimes to complete its normal growth development and fruit setting. The study provides insights into the optimum light regimes for the improvement of the quality and efficiency of C. oleifera in P. massoniana understory plantations.
1. Introduction
Camellia oleifera (Abel) is an important woody oil plant with high economic value in South China, where it is distributed in the hilly and mountainous areas of Guizhou Province, at an altitude of 800–1000 m [1]. Camellia oil is rich in octadecenoic acid, which is good for human health because it has an unsaturated fatty acid content of 80.34–86.18% [2]. Therefore, Camellia oil is known as the “Oriental Olive Oil”. C. oleifera is a light-loving plant and, therefore, requires adequate light for flowering and fruit setting [3]. A weak light will postpone the differentiation of treetops and flower buds and lead to the formation of slender branches, delaying the accumulation of seed oil [4]. Thus, low yield could be one of the main restricting factors during the course of the C. oleifera industry [5].
Since 2000, the project of returning farmland to forests has been widely carried out in Guizhou Province, and the province has focused on the development of the C. oleifera industry. The local government has made relevant policies, such as “the Guizhou Province C. oleifera Industry Development Plan (2019–2025)”, in which 600 km2 of new high-yield C. oleifera forests were to be created in 2019 and 2020 [6], which greatly promoted the expansion of the C. oleifera planting area. At that time, a large number of camellia trees were planted under Pinus massoniana plantations. However, with the gradual development of P. massoniana individuals, the intensity and duration of sunlight in the understory plantations gradually decreased, which remarkably influenced C. oleifera growth. C. oleifera plants have experienced excessive vegetative growth, and produce flowers but do not bear fruits, and, therefore, fruit dropping cannot be found in the area. These abnormal phenomena dampened the enthusiasm of forest farmers planting C. oleifera. A large area of the C. oleifera plantation has been unmanaged or even abandoned. Hence, studying the causes of the degradation of C. oleifera forests and restoring their utilization value are among of the most important problems that people in Guizhou Province should be giving their attention to [7,8].
Light regimes in the understory substantially affect the growth and development of understory plants, especially shrub plants [9]. The leaf, as an important organ for sensing and utilizing light energy, has many physiological functions, such as photosynthesis, water evaporation, and respiration metabolism [10]. Therefore, leaf response to a light regime is also very sensitive [2]. Guo et al. [11] found that the medium light regime was the best habitat for the endangered Sinopodophyllum hexandrum, based on the highest values of LA and Pnmax. Liu et al. [12] reported that Schima lanceolata is better adapted to low light conditions than Cunninghamia lanceolata because of its enlarged leaf area and increased carbohydrate reserves, which allow the plant to cope with low light stress. Valladares et al. [13] also proved that low photon flux density thins the leaves and loosens the spongy tissue to increase air permeability among tropical rainforest shrubs. The adaptation of plant leaves to different light regimes is diverse. However, the responses of C. oleifera (leaf morphology and physiological traits) to the change in light regimes are still unknown [14].
Light regimes remarkably affect the growth and survival of understory plants in a forest community [15]. Thus, light systems lead plants to gradually develop adaptability, called phenotypic plasticity, under different light intensities [16]. Phenotypic plasticity depends on the interaction between plants and the environment, which makes plants behave differently under various habitat conditions [17]. During this process, light regime plays an important role in plant growth and development. The phenotypic plasticity of plants adapts to different light environments through changes in morphology, structure, element contents, and other traits, and maximizes the use of resources to survive in unfavorable habitats [18].
According to Stotz et al. [19], the phenotypic plasticity of plants could be divided into four categories, as follows: leaf morphology, physiology, size, and performance. Actually, the plasticity level of plants determines the ability of plants to respond to environmental changes [20]. Usually, the morphological parameters of plant leaves present considerable changes, such as a thinner leaf blade and an expanded leaf area under low light intensity [21]. Moreover, plant leaf area, specific leaf area, chlorophyll a, chlorophyll b, and total chlorophyll content decreased with the increase of light intensity [22,23]. Under strong light regimes, more mesophyll cells with thick palisade tissue developed. [24]. A study was undertaken in order to explore whether C. oleifera would adopt a trade-off strategy between twig and leaf traits in under-forest environments with different light intensities to improve the utilization of light resources or enhance the abilities of survival and defense. Corner’s rules [25] also illustrated that the relationship between twigs and leaves can explain the adaptation strategy of plants to the environment, and the economic values of flowers, fruits, leaves, and other components on twigs are a factor that cannot be ignored for understory plants. Wright et al. [26] proposed the concept of the “leaf economic spectrum” (LES) to explain that ecological strategies and the adaptive and responsive functions of plants should be regulated through the trade-off mechanism of resource compensation, allocation, and balance. Thus, the diversified light intensity has remarkable influences on leaf structure, physiological and chemical traits, photosynthetic function, and twig size.
The preliminary investigation revealed that a large number of C. oleifera individuals are growing under P. massoniana plantations, and C. oleifera is also the dominant shrub species in Southeast Guizhou Province, China. According to our observation, the leaf morphology and physiological traits, fruits, and twig size of C. oleifera in the understory show a marked difference because of the different light intensity. In addition, the fruit yield of C. oleifera plants in the study area shows low or no fruit yield, because of the lack of effective management measures. Therefore, we selected mature (10 years old) C. oleifera plants under four light regimes (CK: 100% light intensity located in open ground; HL: about 75% of CK located at the edge of P. massoniana plantations; ML: about 50% of CK located in the gap of P. massoniana plantations; LL: about 30% of CK located in the shade of P. massoniana plantations) as the research object to explore their differences in growth and development. Our aims were as follows: (a) to explore the leaf morphology, leaf physiology, twig size, and performance of C. oleifera under different light intensities, and to calculate their plasticity indices, and (b) find out the significant indexes which affected C. oleifera growth and development with the change in light intensity. The results of this study will further improve our understanding of the adaptation of C. oleifera under different light regimes, which is valuable for formulating reasonable management measures for C. oleifera.
2. Materials and Methods
2.1. Study Site
The study was carried out in Qingyun Town (109°11′ E, 25°83′ N; 997 m altitude), Congjiang County, Southeast Guizhou Province China, (Figure 1). The research site has a total area of 8.5 ha and an altitude of 354–997 m. It belongs to the southern edge of the mid-subtropical humid monsoon climate zone, with an annual mean temperature of 18.5 °C and annual mean relative humidity of 80%. The annual average sunlight hours are 1248.1 h. The annual precipitation reaches 1185.9 mm, which is concentrated from April to August every year, and accounts for about 71% of the annual precipitation. The soil type is acid yellow soil, with minor variation in soil nutrient content (total carbon [TC]: 26.01–27.18 mg·g−1; total nitrogen [TN]: 1.05–1.15 mg·g−1; total phosphorus [TP]: 0.65–0.69 mg·g−1) and the pH value (4.3–5.6).
Figure 1.
Satellite image of Qingyun Town, Congjiang County, Guizhou Province.
According to prior soil sampling analysis, the different areas of the study site have no remarkable differences in soil TC, TN, TP, and stoichiometric ratios. The site is a pure 21 year old P. massoniana plantation with many C. oleifera plants in the shrub layer. Ten year old C. oleifera “Xiangxi No. 02” grafted seedlings (1.63–1.82 m height, and 12.50–13.81 mm diameter at breast height), which were only irrigated, pruned, and managed in the first three years were selected. C. oleifera (Abel.) blossoms in mid-October, and its fruits are harvested in late October of the following year. In the study site, the main canopy trees are the P. massoniana, and the main shrub species are C. oleifera, Viburnum dilatatum (Thunb.), Loropia chinense var. rubrum, and Vaccinium bracteatum (Thunb.). The main herbs are Rottboellia cochinchinensis (Lour.), and Cibotium barometz (Lour.).
The field experiment was conducted from July 25 to July 31, 2020. The four light regimes were determined according to the average daily light intensity from 8:00 to 18:00, as measured using a 3415FQ quantum sensor (Spectrum Technologies, Inc. Aurora, IL, USA). Referring to the method of Xu [27], the sensor was set in the middle part of the canopy top layer of each C. oleifera observation point and used to measure the light intensity of photosynthetically active radiation (PAR, 0–2000 μmol m−2·s−1), and to use the average light intensity of six observation points per day to represent the light intensity of C. oleifera under different light regimes. The light regimes were CK (100% light intensity, 1302.0 µmol m−2·s−1), HL (75% light intensity, 976.5 µmol m−2·s−1), ML (50% light intensity, 651.0 µmol m−2·s−1), and LL (30% light intensity, 390.6 µmol m−2·s−1). No remarkable differences in air temperature and humidity were observed between the four light environments. However, significant differences (p < 0.001) in light intensity were noted between the four light regimes, in the following order: CK > HL > ML > LL. Under each light regime, five C. oleifera individuals with similar growth conditions were selected as the subjects. Meanwhile, soil samples in a 0–20 cm layer near the five subjects were collected to determine their physicochemical characteristics.
2.2. Morphological Parameters and Biomass
Five functional leaves (5 branches) were selected from the labeled individuals to measure the morphological parameters and biomass. Leaf length (LL), leaf width (LW), leaf area (LA), and perimeter (LP) were measured using an AM305 leaf area meter (AM350, ADC, UK), and leaf thickness (LT) was determined using digital vernier caliper (EMKC150, Elecall, Shanghai, China). Fresh weight (FW) was measured using an electronic balance (JT1003B, Jinnuo, Ningbo, China), and samples were then placed into an oven (DHG-9070B, Huarun, Changzhou, China). The leaf samples were dried to a constant weight at 60 °C to determine dry weight (DW). Leaf succulence (LS) was calculated by the ratio of the leaf FW and DW, and specific leaf area (SLA) was calculated by the ratio of LA and leaf dry mass (FD) [28].
2.3. Anatomical Structure and Stomatal Structure
Under four light regimes, five plants of C. oleifera were selected for each treatment, and five mature leaves with the same light direction and time were selected for each plant. Immediately after picking, FAA was added. Whole function leaves were selected from the labeled individuals and used in making paraffin sections [29]. The steps are as follows. A leaf was cut into a 4 mm × 4 mm section along the main vein at the base 1/3 of the leaf, and fixed with 70% formalin–acetic acid–70% ethyl alcohol (=5:5:90 v/v/v) immediately after picking. A syringe was used to pump air for 15–30 min to eliminate the interference of air bubbles, and the fixed time was not less than 24 h. The sections were prepared using alcohol (70%, 85%, 95%, 100%, 100%, and 100%) and xylene gradient dehydration, paraffin immersion, and embedding. The leaf was sliced into 8–10 μm thick slices, dyed with safranin–fast green double dye, and sealed with neutral gum to make the slices permanent. The slices were photographed using an optical microscope (DM2500, Leica, Germany). Image J software (Wayne Rasband NIH, http://imagej.nih.gov/ij/index.html (accessed on 24 December 2021)) was used to measure the upper cuticle thickness (UCT), lower cuticle thickness (LCT), upper epidermis thickness (UET), lower epidermis thickness (LET), palisade tissue thickness (PT), sponge tissue thickness (ST), and PT/ST ratio. Twenty fields of view were selected, and the average value for each treatment was calculated.
A leaf piece (2 mm × 2 mm) was collected from the middle of the fresh specimen at one-third from the leaf base. Then, we applied the nail polish evenly to the leaves and let them dry naturally 3 to 8 min. The stoma was measured using the nail polish imprinting method [30] with an optical microscope (DM2500, Leica, Germany). Permanent slices of the leaf’s lower epidermis were prepared according to the standardized method. Images were captured using Leica Application Suit X 3.0.2 (LAS X, Leica, Germany), which were used to determine stoma length, width, area, circumference, and density using Image J software (Wayne Rasband NIH, http://imagej.nih.gov/ij/index.html (accessed on 24 December 2021)).
2.4. Chlorophyll Content
Under different light regimes, 30 functional leaves with the same light direction, time and growth trend were measured for each treatment, and the average value was taken under each treatment. Chlorophyll a and b, chlorophyll a/b, and total chlorophyll were quantified from fresh leaves using acetone and 95% alcohol (= 1:2 v/v). The leaf samples were cut into 2 mm2 pieces. Fresh leaves (0.1 g) were extracted using 10 mL extraction, and chlorophyll was extracted with acetone, as follows: 95% alcohol = 1:2 in the dark for 24 h. Each treatment was repeated three times, and 10 mL of the extract was added to each tube and shaken regularly. The extract was analyzed at 645 and 663 nm using a spectrophotometer (UV-5500, Shimadzu, Kyoto, Japan). Chlorophyll content was calculated using the following equations proposed by Lichtenthaler [31]:
where A663 and A645 are the absorbances at the wavelengths of 663 and 645 nm, respectively, V is the volume of the extract, and W is the FW of the leaves.
Chlorophyll a content (mg g−1 Chl a) = (12.7 A663 − 2.69 A645) × V/(1000 × W)
Chlorophyll b content (mg g−1 Chl b) = (22.7 A645 − 4.68 A663) × V/(1000 × W)
Total chlorophyll content (mg g−1 Chl T) = Chl a + Chl b
2.5. Soil and Leaf Analysis
Soil samples (100 g) were drilled from the surface with 5 cm of soil in diameter, and the collected soil was placed into envelopes for preservation and marked for the determination of soil nutrient content. Then, 30–50 whole functional leaves were selected from the labeled plant individuals, and the collected leaves were stored in envelopes and labeled. The nutrient contents of the leaves were then determined.
The soil samples collected from the 0–20 cm soil layer were air-dried naturally at room temperature and cleaned to remove rocks, roots, and other debris. The samples were pulverized and passed through an 80-mesh sieve (0.18 mm), weighed, bagged, and sealed for the determination of TC, TN, and TP contents. Fresh functional leaves selected from labeled C. oleifera individuals were dried at 80 °C to constant weight, crushed, passed through an 80-mesh sieve (0.18 mm), weighed, bagged, and sealed for the determination of TC, TN, and TP contents. The TC and TN contents were determined with total organic carbon analysis and the Kjeldahl method [32], respectively. The TP content was determined with the Mo–Sb colorimetric assay method by heating and digesting with concentrated sulfuric acid and a hydrogen peroxide–molybdenum–antimony ratio in a Shimadzu UV2450 spectrophotometer. All the samples were made in three replicates, and the result was expressed in mean ± standard deviation.
2.6. Photosynthetic Parameters
Three to five whole functional leaves from the labeled individual’s outer canopy were selected as the measurement objects. Their photosynthesis-light response curves were measured between 9:00 and 11:00 h in sunny days with a Li-6400 photosynthesis instrument (Li-Cor Inc., Lincoln, NE, USA). During the measurement process, the light intensity gradient in the chamber with a red/blue light source (6400-02B) [33] was set to 1800, 1600, 1400, 1200, 1000, 800, 600, 400, 300, 200, 100, 50, and 0 μmol·m−2·s−1, and the temperature and relative humidity (RH) in the chamber were uniformly set as 25 °C and 60%, respectively. The instrument automatically records photosynthetic parameters. Maximum net photosynthetic rate (Pmax), light saturation point (LSP), light compensation point (LCP), and apparent quantum yield (AQY) were calculated using Thornley’s nonrectangular hyperbola model (NRH) [34], and the equation is as follows:
where PN is the net photosynthetic rate, PAR is photosynthetically active radiation, apparent quantum yield (α) is calculated as the slope of regression line, and α, PAR, PN, Pmax, and dark respiration rate (Rd) values are as described above. The LCP and LSP of C. oleifera were the PAR values when the values of PN are zero and maximum value, respectively, while k is the convexity.
2.7. Twigs Parameters
Four representative one year old twigs were selected from the east, west, south, and north of the labeled individuals’ outer canopy to measure their parameters under different light regimes. First, the length (TL), diameter (TD), number of leaves (LN), and number of fruits (FN) of each twig were measured or counted, and fruit diameter (FD) and fruit longitudinal diameter (FLD) were measured with a digital vernier caliper (EMKC150, Elecall, Shanghai, China). Finally, all the measured twigs and fruits were classified, placed in plastic bags, and brought to the laboratory in order to measure their biomass.
2.8. Plasticity Index (PI) Calculation
The PI of a parameter was calculated according to Valladares’ method [13], using the following equation:
PI = (maximum mean − minimum mean)/maximum mean
The PI ranges from 0 to 1, in which a larger value indicates, greater phenotypic plasticity.
2.9. Statistical Analysis
The data of the experimental parameters were analyzed using ANOVA in the SPSS 26.0 statistical software (SPSS Inc., Chicago, IL, USA), followed by Duncan’s test (p < 0.05). The R language package (version 4.1.1, http://www.R-project.org/ (accessed on 26 December 2021) R Development Core Team 2011) and the ggplot 2 package (Version 3.3.2, http://ggplot2.org (accessed on 26 December 2021), https://github.com/hadley/ggplot2 (accessed on 26 December 2021)) were used to conduct the principal component analysis (PCA) and plot the graph, respectively. All the other figures were plotted using Origin 2018 (Origin Lab Corporation, Northampton, MA, USA). All the data were presented as mean ± standard deviation.
3. Results
3.1. Leaf Morphology and Biomass
The morphological parameters of C. oleifera leaves changed significantly under different light regimes (p < 0.05, Table 1). As the light regimes decreased, the leaves gradually became larger. The average LL of C. oleifera increased under ML and HL, and the LW increased. Under LL, the difference between LL and LW was significant (p < 0.05, Table 1). The LA, LC, and SLA increased with the decrease of light regimes. The LA, LC, and SLA with light increased by 31.04%, 11.10%, and 37.48%, respectively, compared with that under LL. The LA, LC, and SLA increased by 18.02%, 7.81%, and 4.90%, respectively, compared with HL. The LB and LT of C. oleifera were the highest under HL, but no significant difference in LB was noted (p > 0.05). The plasticity index of SLA and LA was higher, but the other indexes were lower. The average PI values of leaf morphological parameters were 0.17.

Table 1.
Leaf morphological characteristics of C. oleifera under different light regimes.
3.2. Anatomical Structure of Leaves
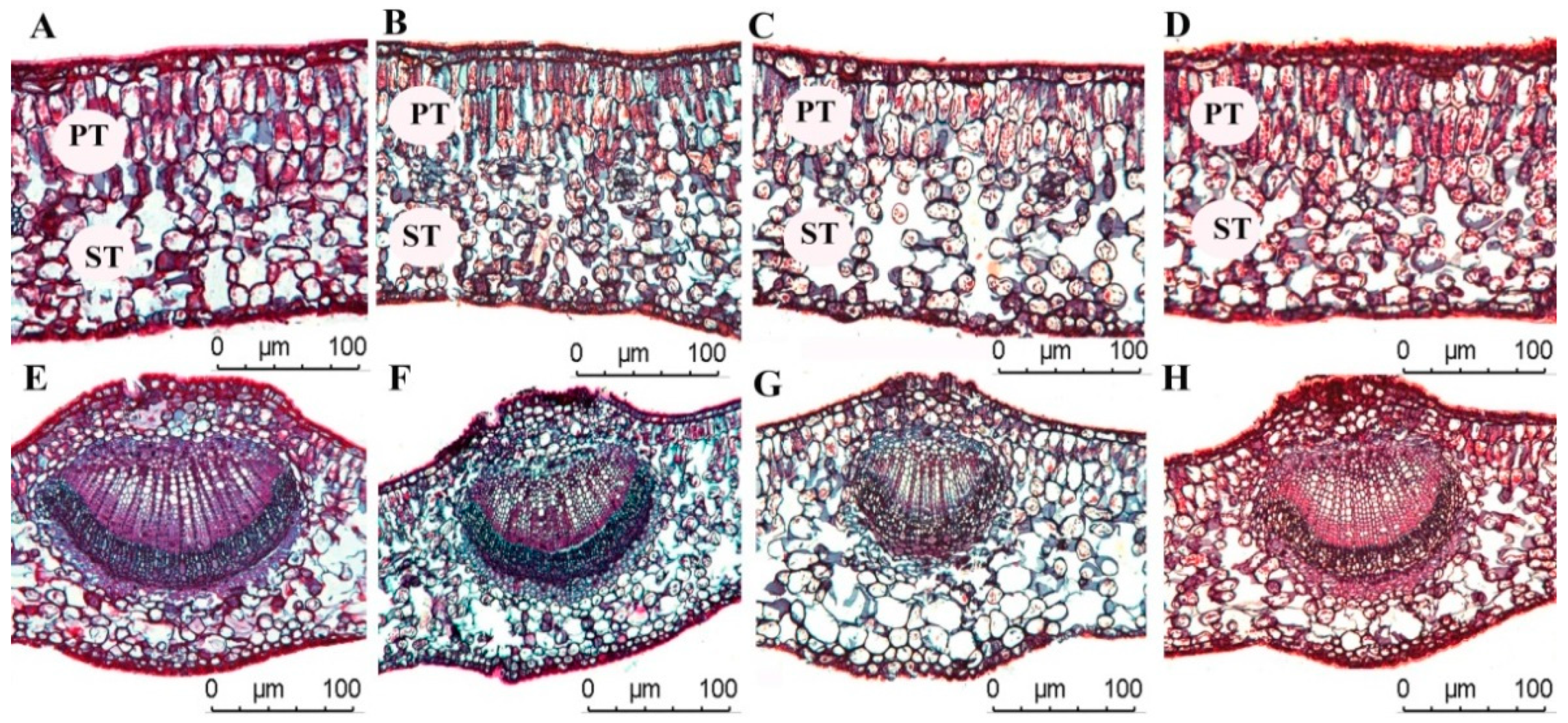
A one-way ANOVA showed (Table 2) that UCT and LCT increased alongside the increase of light regimes (p < 0.05), and no significant difference in UCT was found between CK and ML in UCT (p > 0.05). No significant difference in LCT was found among CK, HL, ML, and LL (p > 0.05), and the UCT and LCT of C. oleifera leaves were 3.52 ± 0.88–5.41 ± 0.71 and 2.67 ± 0.52–3.24 ± 0.41 μm, respectively. The HL and LL light regimes were significantly lower than other light regimes. The UET and LET of C. oleifera leaves increased with the increase of light regimes, and the difference between the light regimes was significant (p < 0.05). The variation ranges of UET and LET were 14.08 ± 1.81–22.29 ± 1.53, and 12.65 ± 0.94–15.72 ± 0.71 μm, respectively. The PT and ST in C. oleifera leaves increased with the increase in light regimes. No remarkable differences in PT/ST were found between CK and HL in PT/ST (p > 0.05), and PT/ST was lesser under ML than in LL (p < 0.05). The PI of UET parameters was higher than those of other leaf anatomical parameters, and the average PI of leaf anatomical parameters was 0.24 (Table 2 and Figure 2).
Table 2.
Anatomical characteristic parameters of C. oleifera leaves under different light regimes.
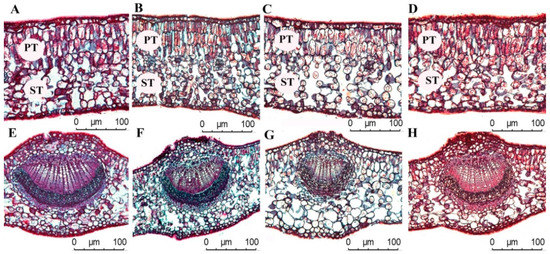
Figure 2.
Anatomical characteristics of C. oleifera leaves under different light regimes. Note that CK: 100% light regime (A,E); HL: 75% light regime (B,F); ML: 50% light regime(C,G); LL: 30% light regime (D,H).
3.3. Leaf Stomata Structure
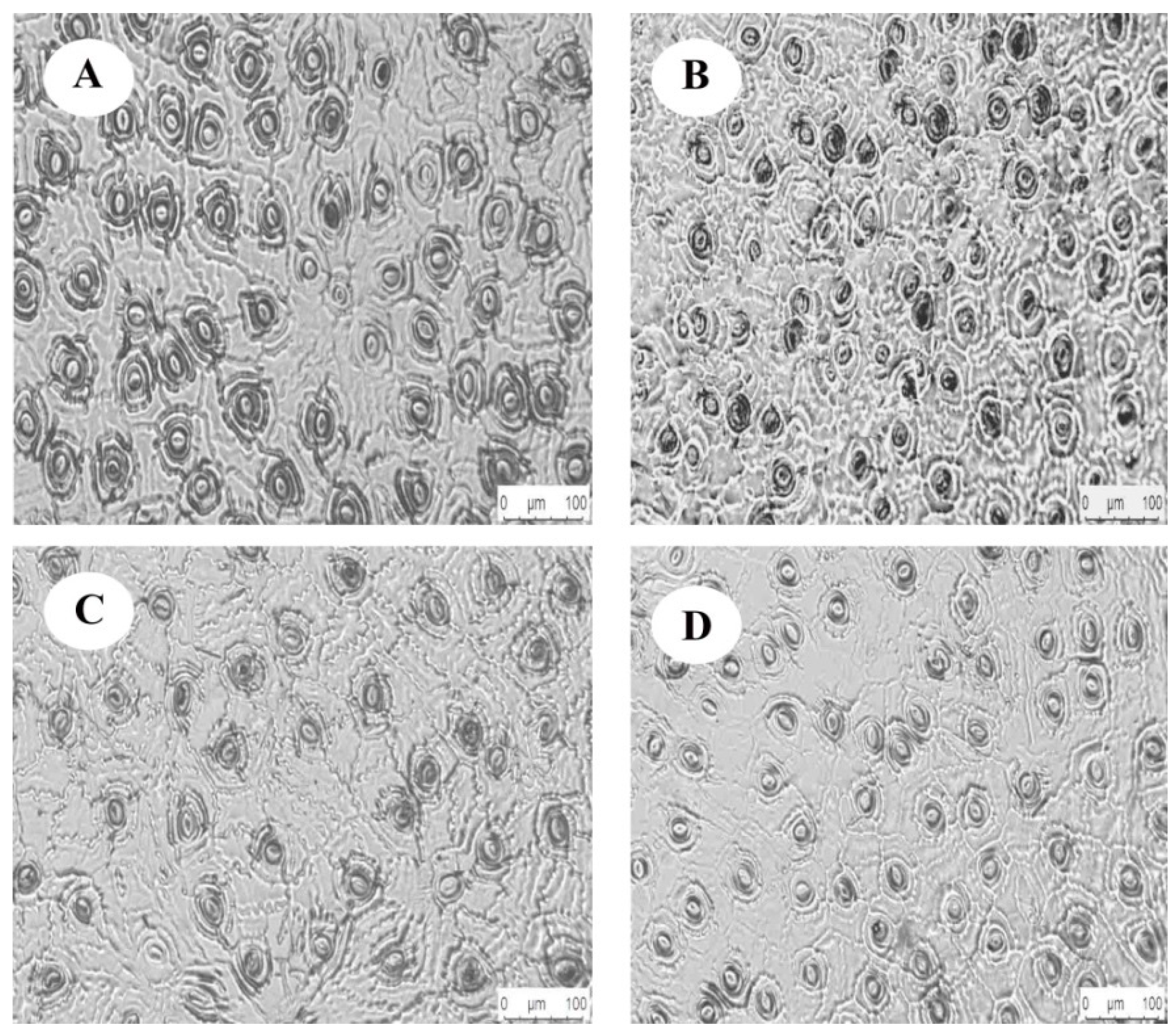
The one-way ANOVA showed (Table 3) that light regimes had a significant effect on the HAS and the VAS (p < 0.05), and no significant differences with HAS and VAS were observed between HL and LL (p > 0.05). Figure 3 showed that the HAS and VAS in the leaves increased with the increase in light regimes. The HAS and VAS of C. oleifera leaves vary from 18.44 ± 1.05–23.48 ± 1.23, 14.31 ± 0.97–17.23 ± 1.89 μm, respectively, and were significantly lower under HL and LL than under other light regimes. Significant differences in stomatal density (SD) and stomatal area (SA) were found in the C. oleifera leaves under the four light regimes (p < 0.05). The SD under HL was significantly higher than in other light regimes (p < 0.05), as was SA under HL. The lower light regime was significantly lower than other light regimes (p < 0.05), and the variation range was 124.6 ± 4.8–193.4 ±7.86 and 141.62 ± 20.21–203.61 ± 8.62 μm, respectively. The SP had a variation range of 48.13 ± 2.62—57.15 ± 2.11 μm and had no significant difference between CK and HL (p > 0.05). The SP under LL was the smallest and was significantly lower than the SP in the other light regimes (Figure 3, p < 0.05).
Table 3.
Stomatal structure characteristic parameters of C. oleifera leaves under different light regimes.
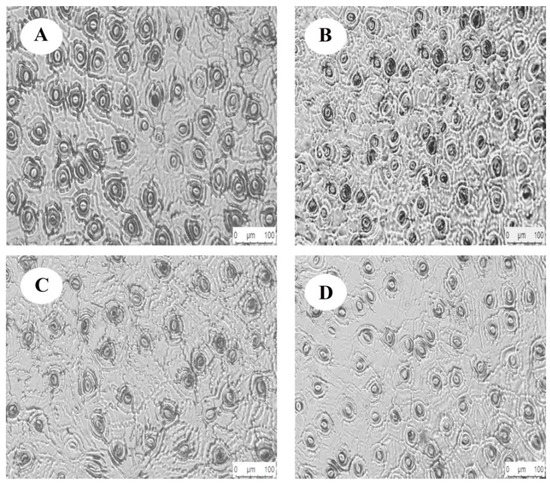
Figure 3.
Stomatal structure of C. oleifera leaves under different light regimes. Note: CK: 100% light regime (A); HL: 75% light regime (B); ML: 50% light regime (C); LL: 30% light regime (D).
The HAS, VAS, SD, SP, and SC were significantly different among the different light regimes (p < 0.05). The values of HAS in the four regimes were in the following order: ML > CK > LL > HL. The VAS values were in the following order: CK > ML > LL > HL, while the SD order was HL > CK > LL > ML. The values of SP were in the following order: ML > CK > HL > LL. The PI of SD was higher than those of the other stomatal parameters, and the average PI of the stomatal parameters was 0.24.
3.4. Chlorophyll Content
Figure 4 showed that the contents of chlorophyll, chlorophyll b, and total chlorophyll contents in C. oleifera gradually decreased with the increase in light regimes under the forest. Their variation ranges were 1.248 ± 0.11–1.657 ± 0.08, 0.351 ± 0.05–0.637 ± 0.07, and 2.153 ± 0.07–2.153 ± 0.07 mg·g–1, respectively. Chlorophyll content under LL was significantly higher than those of other light regimes (p < 0.05), whereas the chlorophyll contents of HL and ML had no significant difference (p > 0.05). Chlorophyll a/b decreased with the increase in light regimes (p > 0.05). The variation range was 2.6 ± 0.07–3.56 ± 0.06 mg·g–1, and no significant differences in chlorophyll a/b were found between ML and LL (p > 0.05).
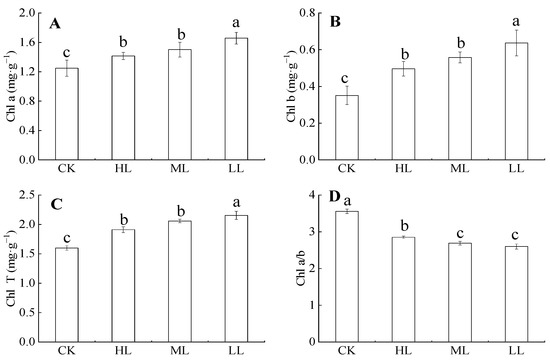
Figure 4.
The content of chlorophyll in C. oleifera under different light intensity treatments. Note: Parameters include Chl a (Chlorophyll a content; A), Chl b (Chlorophyll b content; B), Chl T (total chlorophyll content; C), and Chl a/b (Chlorophyll a/b content; D) of C. oleifera under different light regimes. Different lowercase letters (a, b, c) indicate significant differences between light regimes (p < 0.05). Regime acronyms are defined as CK: 100% light regime; HL: 75% light regime; ML: 50% light regime; LL: 30% light regime.
3.5. Leaf Nutrient Content
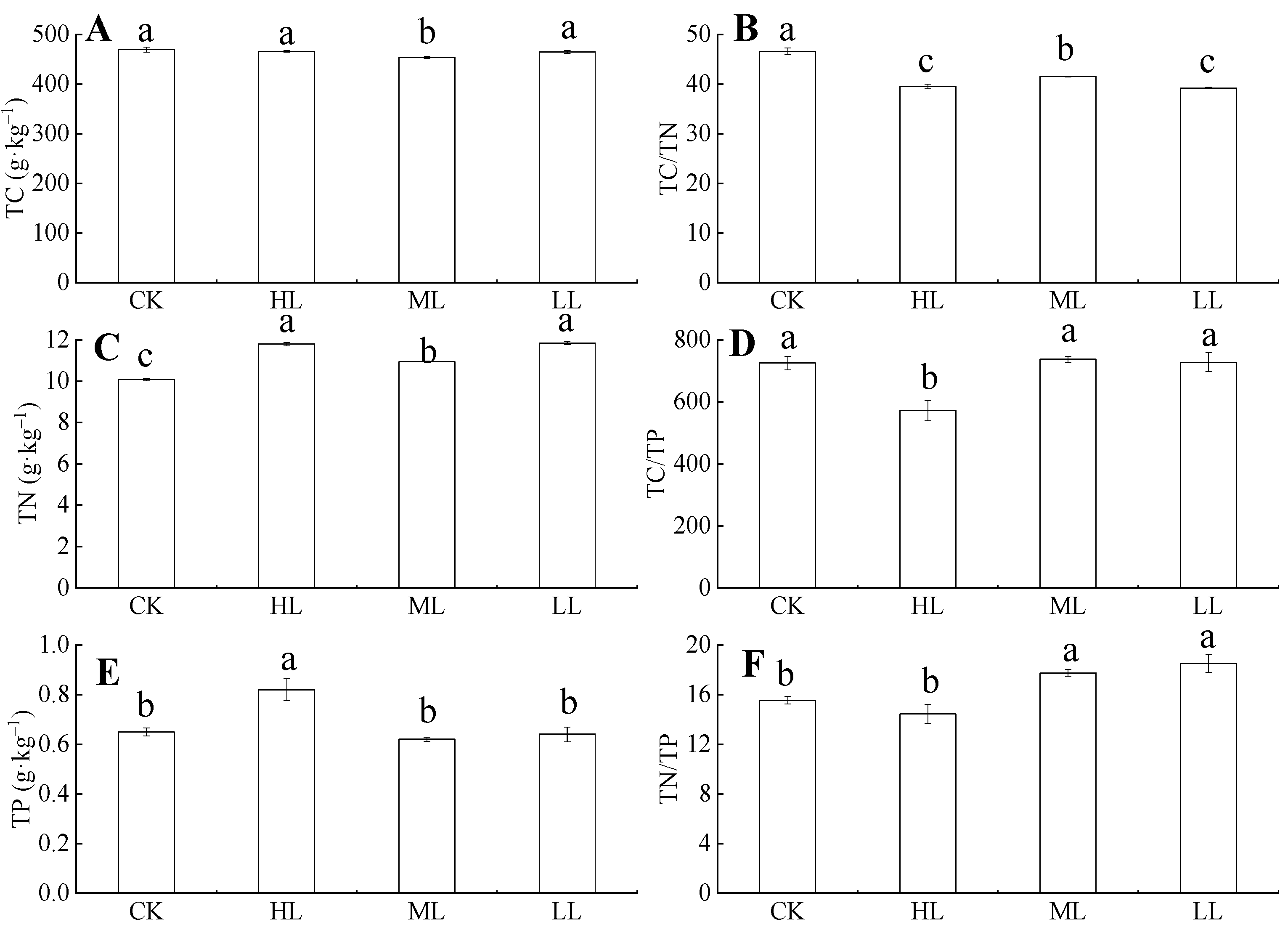
Figure 5 showed that, under ML, the total carbon (TC) content of C. oleifera leaves decreased, and the variation ranges were 469.98 ± 4.98–466.08 ± 1.65 g·kg–1 and 454 ± 2.16–465.06 ± 2.50 g·kg–1. The ML samples had significantly lower TC than the other light regimes (p < 0.05), whereas the TC contents under CK, HL, and LL had no significant difference (p > 0.05). The TC content did not change significantly with the increase in light regimes (p > 0.05), and the variation range was 454 ± 2.16–469.98 ± 4.98 g·kg–1. Total nitrogen (TN) contents in each of the leaves from the different light regimes were significantly different (p < 0.05), and the variation ranges were 10.08 ± 0.06–11.79 ± 0.08 g·kg–1, 10.93 ± 0.05–11.84 ± 0.07 g·kg–1. Total phosphorus (TP) content in leaves initially decreased and then increased with the increase in light regimes. The ranges were 0.92 ± 0.05–1.33 ± 0.13 g·kg–1 and 0.29 ± 0.03–0.71 ± 0.06 g·kg–1, respectively, and, under HL, the TP was significantly higher than other light regimes (p < 0.05). The TP content in C. oleifera tends to decrease with the increase of light regimes, but the difference was not significant (p > 0.05). However, under HL, TP is significantly higher than under other light regimes (p < 0.05), and the variation range is 0.62 ± 0.008–0.82 ± 0.04 g·kg–1. The TN and TP contents had significant differences among the different light regimes. The TN contents were ordered as follows: HL > LL > ML > CK, whereas TP contents were ordered as follows: HL > CK > LL > ML.
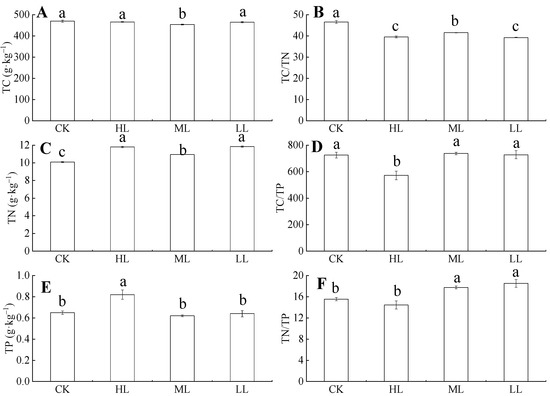
Figure 5.
The TC, TN, TP contents and their ratios in C. oleifera leaves under different light regimes. Note: Parameters include TC (A), TC: TN (B), TN content (C), TC: TP (D), TP (E), and TC: TP (F) of C. oleifera under different light regimes. Different lowercase (a, b, c) letters represent significant differences between light regimes (p < 0.05). Regime acronyms are defined as CK: 100% light regime; HL: 75% light regime; ML: 50% light regime; LL: 30% light regime.
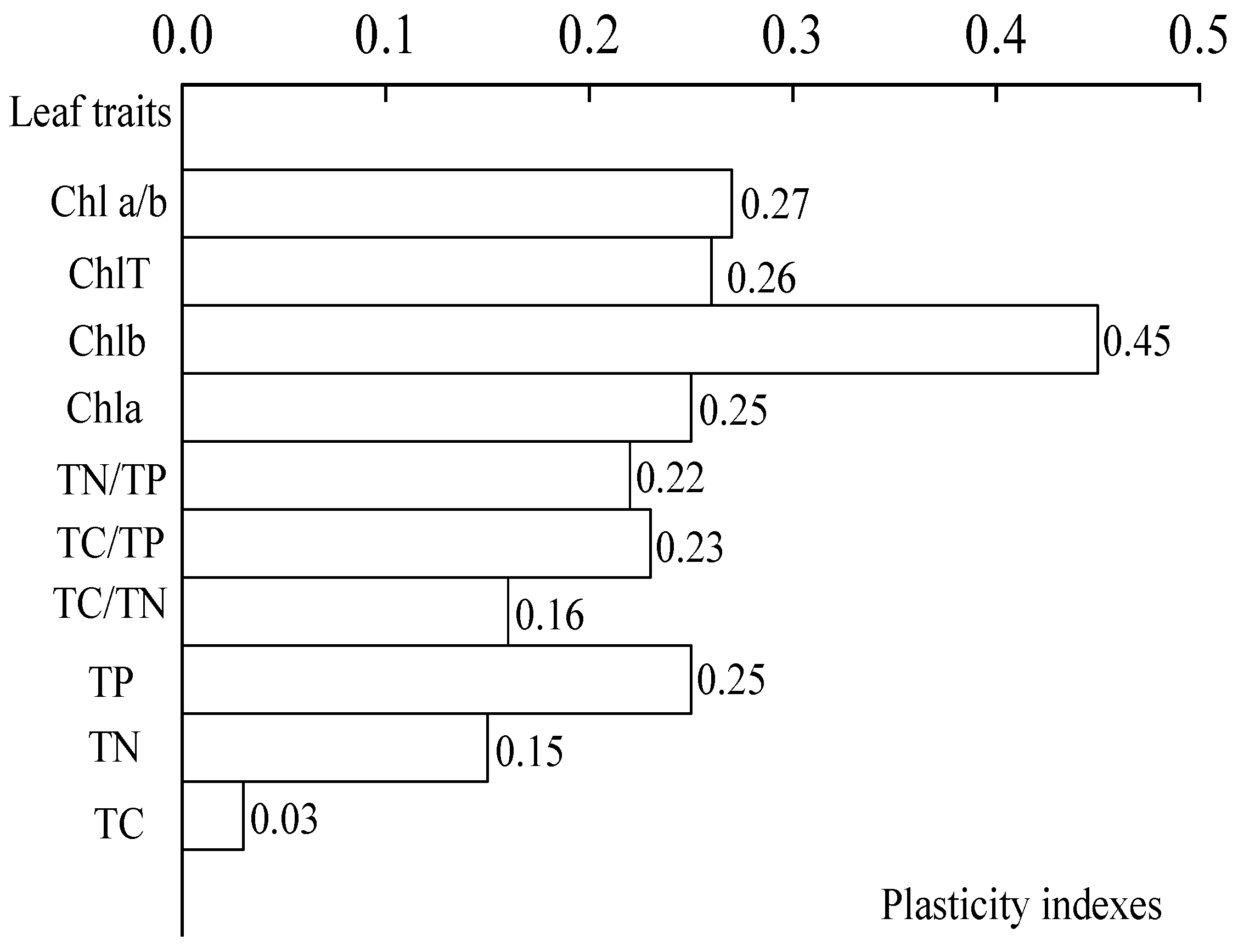
A one-way ANOVA (Figure 5) showed that the light regimes had a significant effect on TC/TN, TC/TP, and TN/TP ratios (p < 0.05). Figure 5 showed that the TC/TN ratio in leaves tends to decrease with the increase of light under the four light regimes, and the variation range of TC/TN ratio was 39.26 ± 0.12–46.62 ± 0.69. The TC/TN ratio in CK was significantly higher than those in HL, ML, and LL (p < 0.05). The TC/TP ratio of C. oleifera leaves initially decreased and then increased, and the variation ranges were 571.29 ± 32.59–571.28 ± 32.59 and 737.38 ± 9.68–727.74 ± 30.32. The TC/TP ratios of the four light regimes were different. The TN/TP ratio in the leaves first decreased and then increased with the increase in light regimes, and the light regime with the lowest TN/TP ratio was HL. The variation ranges were 15.55 ± 0.30–14.45 ± 0.76 and 17.75 ± 0.26–18.53 ± 0.73. The results showed that TP content had the highest PI (0.25). The average PI of the nutrient content indexes was 0.17 (Figure 6).
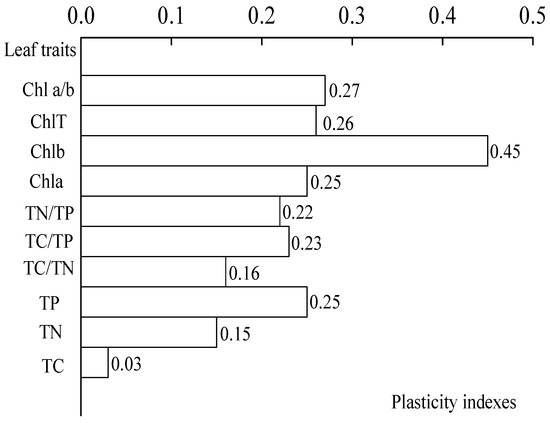
Figure 6.
Plasticity indexes of different characteristics.
The plasticity of the physiological parameters of C. oleifera leaves had the following variation trend (Figure 6). The PI of chlorophyll b content was the highest, and the PI of total chlorophyll was the lowest. Second, the TN content was the lowest PI, and the TP content had the highest PI among the nutrient content indexes. Overall, the average PI of the chlorophyll content indexes was 0.31. In addition, the overall mean PI of the physiological indicators was 0.27.
3.6. Photosynthetic Parameters
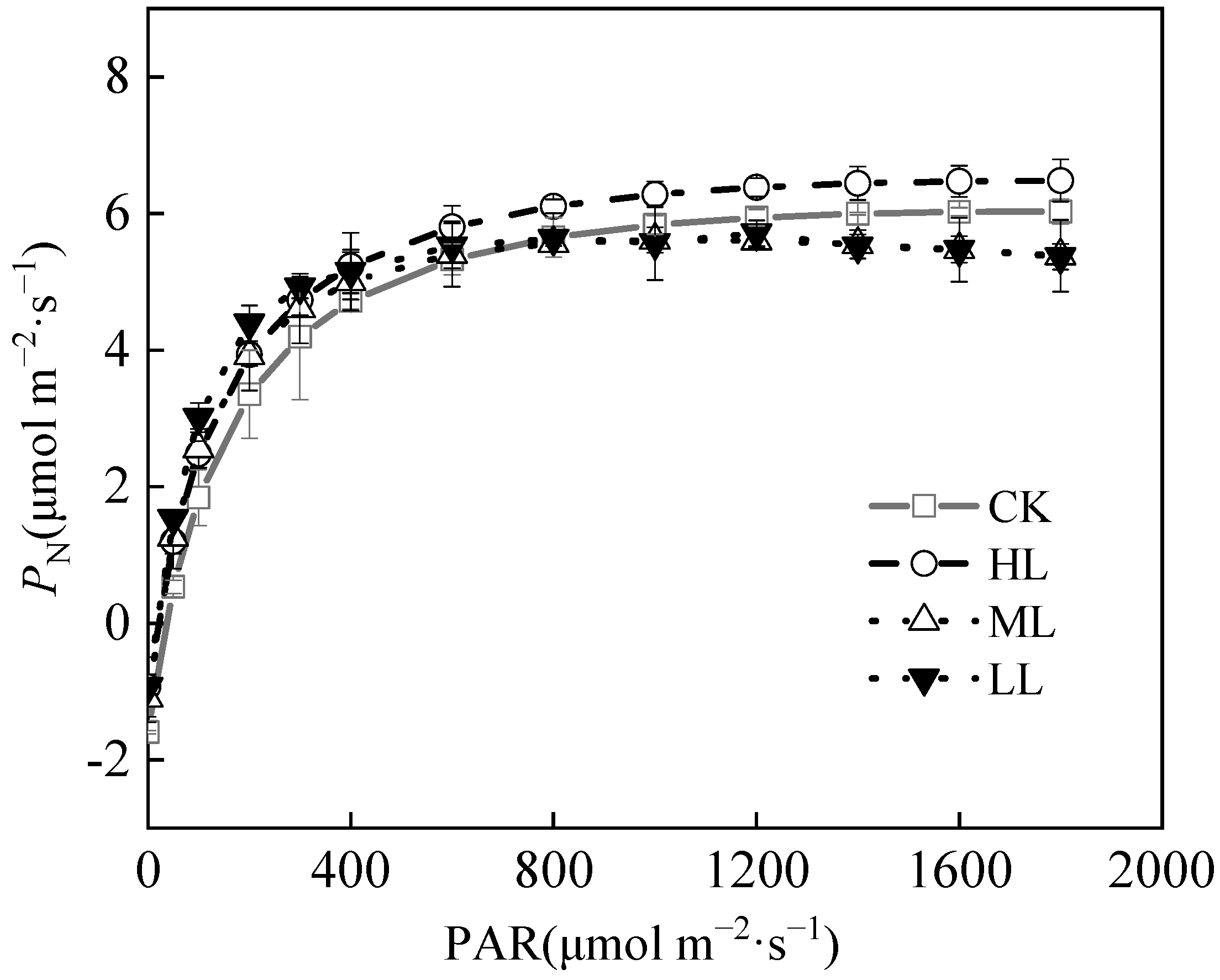
The Pmax of C. oleifera leaves were as shown in Figure 7 and Table 4, and the Pmax of C. oleifera leaves were 6.03 ± 1.02, 6.48 ± 1.34, 5.61 ± 1.18, and 5.66 ± 1.23 μmol m−2·s−1, respectively, under the four light regimes. The Pmax of C. oleifera leaves under HL was higher than those in the other light regimes. The LSP and LCP of C. oleifera leaves reflect the utilization of plant light regimes. The LSP of C. oleifera leaves under the four light regimes were 1753.41 ± 65.26, 1830.12 ± 43.18, 1041.52 ± 43.62, and 932.38 ± 31.54 μmol m−2·s−1, respectively. The LSP showed the highest performance. The LCP values of C. oleifera leaves in the four light regimes were between 34.76 ± 2.13, 18.47 ± 1.25, 19.12 ± 1.06, and 14.07 ± 2.28 μmol m−2·s−1, respectively, while LSP was significantly lower under HL (Table 4, p < 0.05). Compared with CK, the LCP of C. oleifera leaves decreased by 40.6% and 46.82% under ML and LL, respectively. The LCP of C. oleifera leaves reflects the utilization of weak light by plants under different light regimes. Compared with CK, the LCP of C. oleifera had remarkable downward trend with the decrease in light intensity, which indicates the adaptability of C. oleifera plants to different light regimes. The AQY reflects the ability of plants to utilize weak light. The AQY decreased gradually with the increase in light regimes. The AQY of C. oleifera leaves under four light regimes were 0.0558 ± 0.003, 0.0566 ± 0.007, 0.0662 ± 0.005, and 0.0762 ± 0.002, respectively. With the decrease of light regimes, the AQY of C. oleifera leaves was 0.0566 μmol m−2·s−1, which was 1.43% higher than in CK. The result indicates that the ability to use light energy is strong. The Rd of C. oleifera leaves decreased gradually with the decrease in light regimes (p < 0.05) (Table 4; Figure 7). The Rd ranges of C. oleifera leaves under four light regimes were 1.59 ± 0.23, 0.93 ± 0.11, 1.10 ± 0.14, and 0.97 ± 0.25 μmol m−2·s−1, respectively, and CK and ML had significantly higher Rd than the other light regimes (p < 0.05). The PI of LCP (0.60) was slightly higher than the other indexes. The average PI of the photosynthetic parameters was 0.38.
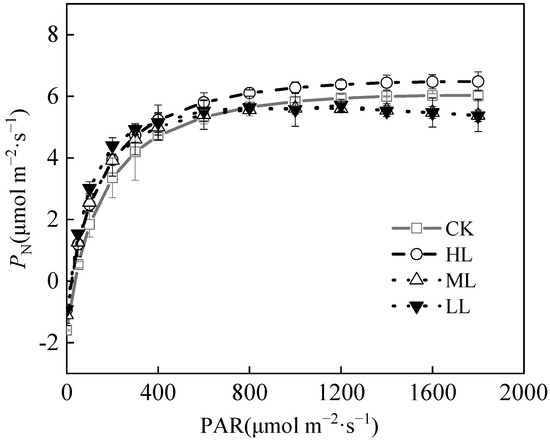
Figure 7.
Photosynthetic curve parameters of C. oleifera leaves under different light regimes.
Table 4.
Photosynthetic parameters of C. oleifera leaves under different light regimes.
3.7. Effects of Light Regimes on Twig Morphological Characteristics
The TD, TL, and LN had significant differences under the four light regimes (p < 0.05). The TD, TL, and LN under CK and HL were significantly higher than those under ML and LL (Table 5; Figure 8; p < 0.05). No twig characteristics were recorded under ML and LL. Fruits appeared under CK and ML, among which the FD, FLD, and FN under HL were larger than those in CK. The average PI of twig plasticity parameters was 0.26.
Table 5.
Morphology and biomass of C. oleifera twigs under different light regimes.

Figure 8.
Twig size of C. oleifera under different light regimes. Note: CK: 100% light regime; HL: 75% light regime; ML: 50% light regime; LL: 30% light regime.
3.8. Phenotypic Plasticity of Leaf Traits in Different Light Regimes
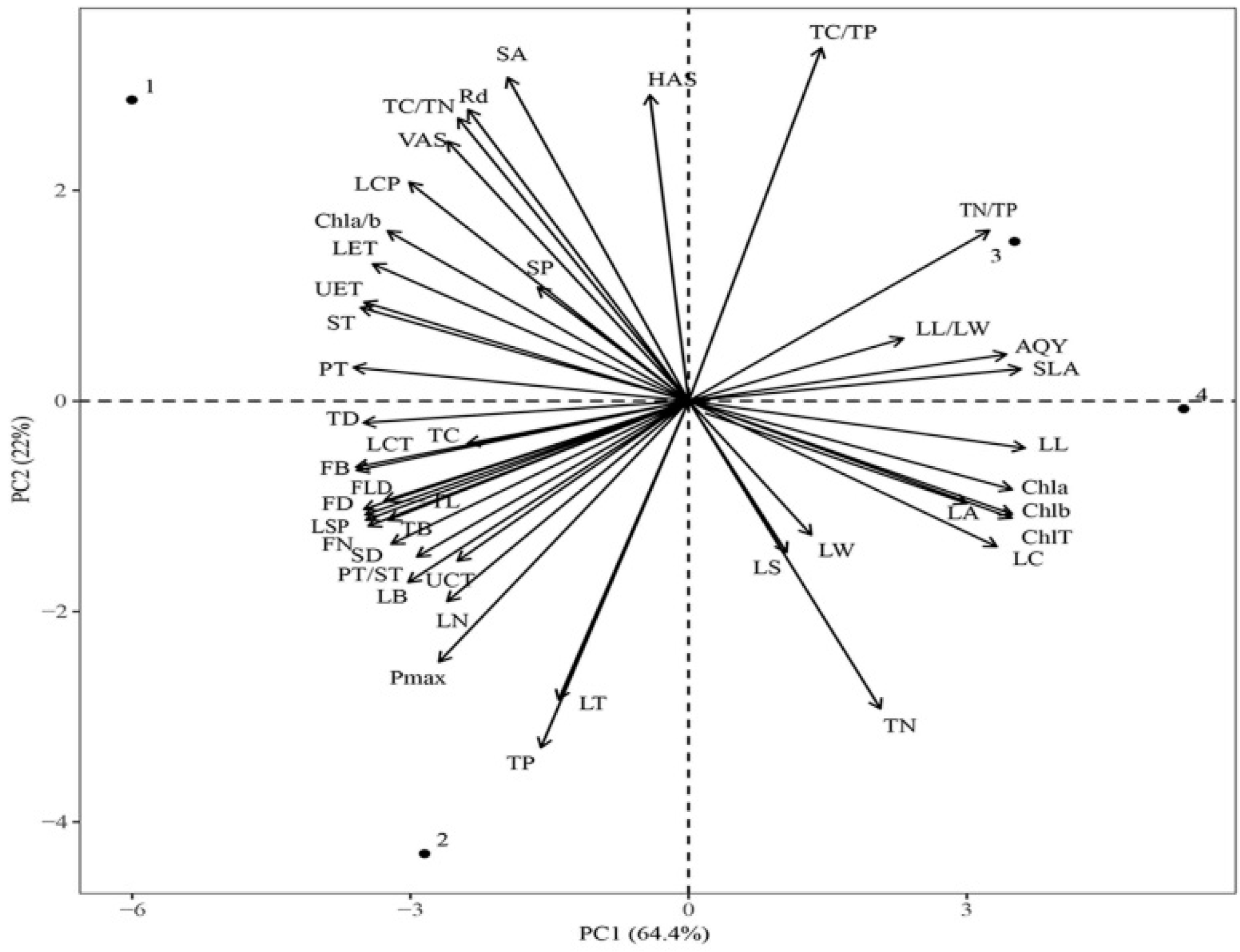
The results showed that the first two principal components could explain 86.4% of the total phenotypic variation under the four light regimes, and PC1 could explain 64.4% of the total variance. The SLA, LL, and chlorophyll content contributed remarkably to PC1. The PC2 could explain 22% of the total variance, and Pmax, TP, TN, LT, and TC/TP contributed substantially to PC2 (Figure 9). The functional traits under CK were significantly different from those under LL and ML, whereas the traits under LL and ML were similar. Leaf traits, such as Pmax and TC content, were significantly different between CK and HL, and SLA and TC/TP were significantly different between LL and ML.
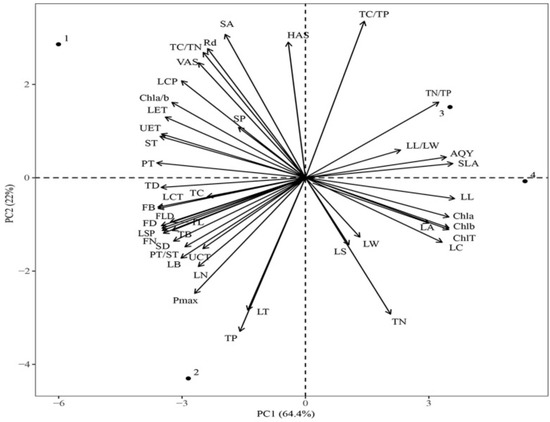
Figure 9.
Principal component analysis (PCA) of functional traits of C. oleifera under different light regimes. Note: Parameters include CK: 100% light regime (1); HL: 75% light regime(2); ML: 50% light regime(3); LL: 30% light regime(4), leaf length (LL), leaf width (LW), leaf length/leaf width (LL/LW), leaf area and circumference (LA and LC), specific leaf area (SLA), and leaf thickness (LT); upper cuticle thickness (UCT), lower cuticle thickness (LCT), upper epidermis thickness (UET), lower epidermis thickness (LET), palisade tissue thickness (PT), spongy tissue thickness (ST), and palisade tissue thickness/spongy tissue thickness (PT/ST); horizontal axes of stomata (HAS), vertical axes of stomata (VAS), stomatal density (SD), stomatal area (SA), and stomatal perimeter (SP); chlorophyll a content (Chl a), chlorophyll b content (Chl b), total chlorophyll content (Chl T), chlorophyll a/b content (Chl a/b), TC, TC/TN, TN, TC/TP, TP, and TC/TP; light saturation point (LSP), light compensation point (LCP), net photosynthetic rate (Pmax), apparent quantum yield (AQY), and dark respiration rate (Rd), twig diameter (TD), twig length (TL), leaf number (LN), twig biomass (TB), fruit diameter (FD), fruit longitudinal diameter (FLD), fruit number (FN), and fruit biomass (FB).
4. Discussion
4.1. Effect of Different Light Regimes on the Leaf Characteristics of C. oleifera
Our findings showed that plants increase photosynthesis by increasing LL with a decrease in light intensity. In this study, LL increased with the decrease in light intensity. This finding was consistent with Han et al.’s research [35]. Under the four light regimes, the SLA of C. oleifera increased considerably with the decrease in light intensity. The results indicated that, as the light intensity decreases, the Camellia oleifera leaves gradually become larger [36]. Moreover, this adaptation is a photoprotection strategy for plants to increase excess light energy captured by leaves. Leaf biomass was the highest under HL, followed by CK (Table 1). In low light regimes, plants usually adapt and optimize their ability to capture light energy and leaf efficiency by increasing the proportion of leaf biomass. Other researchers found similar characteristics in plant response to light [37]. The LS of C. oleifera was higher under ML and HL than under the CK and LL light regimes. The higher LS indicates a higher the relative water content and a higher photosynthetic efficiency. In general, the higher the degree of succulence of plant leaves, and the higher the relative water content, the smaller the water saturation deficit value and the higher photosynthetic efficiency.
4.2. Anatomical Structure and Photosynthetic Function
The light energy utilization of plant leaves depends on their anatomical structure, because the structure of leaf tissues affects the path of light through reflection and refraction. We found that PT, ST, UET, LET, UCT, and LCT decreased with the decrease in overall light intensity. In previous reports, PT and ST increased, the leaves became thicker, and PT developed under CK [38,39]. In the present study, under LL and ML (Table 2), that is, under insufficient light regimes, LT decreased, and PT and ST increased, promoting the leaf to absorb more light to reduce the effect of insufficient light and improving the light energy utilization rate of the leaf. Under sufficient light, LT (PT and ST) increased, which protected the leaves from the damage of strong light [39]. There were many layers of palisade cells in the leaves under CK, and they were more elongated and had increased LT compared with the leaves under low light (Figure 2). In addition, the PT/ST ratio is an important indicator of plant ecological adaptation and an important expression of PN. Under HL, the PT/ST ratio was relatively large, while PN value reached the highest. The HL had no significant difference with CK in terms of PT/ST ratio. This finding suggested that this light intensity and the more developed PT effectively promote photosynthesis. The anatomical structure of C. oleifera leaves was closely related to its photosynthetic characteristics. The LSP and LCP reflect the light demand of plants. The LSP reflects the ability of plants to use strong light. A higher LSP value indicates a higher photosynthetic potential. In this study, the LSP of C. oleifera leaves was higher under HL, which indicated that C. oleifera is suitable for growing in a sunny environment. The LCP was 34.76 μmol m−2·s−1 under CK, and it decreased with the decrease in light intensity. The AQY reflects the light energy conversion efficiency in photosynthesis and the utilization capacity of plants to low light (Table 4, Figure 7). The AQY is an important indicator of plants’ ability to utilize weak light. Under weak light regimes, the apparent quantum efficiency of C. oleifera was higher, indicating that the ability to utilize weak light was stronger. The AQY of C. oleifera decreased under the conditions of ML and LL, indicating that C. oleifera absorbs and converts light energy with more pigment−protein complexes. This indicates that the C. oleifera has a strong ability to utilize weak light, which may be an adaptation of C. oleifera to the external environment.
4.3. Stomatal Structure
The stomata is one of the vital factors that affects plant photosynthesis and transpiration [40,41,42]. Stomata are mostly distributed in the lower epidermis of leaves, and this distribution pattern can not only promote the gas exchange between plants and the external environment, but also assists in retaining water. Under LL and ML (Table 3; Figure 3), plants obtain a lower energy for photosynthesis and evaporation, which requires fewer stomata for gas exchange and, thus, fewer veins to transport water. Under CK light regimes, the light is too strong, meaning that the leaf area of the plant is reduced, the leaves become thicker, and the stomatal density is increased; in order to dissipate the excess excitation energy, the content of chlorophyll decreases, and the chlorophyll a and chlorophyll b ratio (Chla/Chl b) increases [43].
4.4. Chlorophyll and Nutrient Contents
Chlorophyll content indicates the ability of a plant to absorb, transport, and utilize light energy. In this study, the contents of chlorophyll a, chlorophyll b, and total chlorophyll of C. oleifera increased with the decrease in light intensity. The result means that increasing chlorophyll a and chlorophyll b can cause a plant to fix and store more light energy [44]. With the changes in light regimes, plants will adjust their own resources to optimize the chlorophyll and nutrient allocation and adapt themselves to the new environment. Different light intensity affected the carbon and nitrogen metabolism in plant leaves [45]. In low light regimes, plants allocate more carbon to their leaves to gain more light energy [46]. In the photosynthetic mesophyll tissue of leaves, chlorophyll is the irreplaceable photosynthetic pigment of plants that absorbs and transmits the light quantum. Chlorophyll content increased with the decrease in optical quantum density, but the chlorophyll a/b value decreased with the decrease in the optical quantum density. Low chlorophyll a/b value can improve the absorption of far-red light by plants [47]. Therefore, plants with a lower chlorophyll a/b value and higher chlorophyll content under low light also have higher photosynthetic activity (Figure 4).
In this study, plants obtained more light energy under HL. Carbon, nitrogen, and phosphorus are important elements for plant growth and development. Carbon is important to plant photosynthesis and the dry matter accumulation of structural elements, and nitrogen and phosphorus are necessary for the storage of plant materials, such as protein and nucleic acids. These are components of plant photosynthesis related-enzymes that participate in various metabolic activities in plants [48]. The results of this study showed that the TC content of plants was higher under CK and HL, which indicates that C. oleifera has a strong carbon reserve capacity and nutrient utilization strategy. Thus, it shows that the accumulation of C elements is the highest. This is because C. oleifera is an evergreen plant, and the leaves are renewed slowly and have a long life cycle, so that the fixed C elements accumulate in the leaves, which can significantly improve the ability of plant leaves to synthesize organic substances, reflecting that differential growth is slow. It is a “slow growing” type of plant. The contents of TC, TN, and TP (Figure 5) were the lowest under ML. Under the four light regimes, TN content was lower than China nitrogen content (18.6 mg·g−1). The TP content was lower than the average TP content of plants in China (1.21 mg·g−1) [49]. The values of TC, TN, and TP in C. oleifera leaves under different light regimes were different. The result indicates that diversified light regimes affect the utilization efficiency of understory resources and the adaptation strategies of the understory light environment, which are also reflected in the nutrient characteristics of leaf TC, TN, and TP. The TC/TN ratios of C. oleifera plants under four light regimes were higher than the global average value (22.5). Wright et al. [26,50] showed that when the N/P ratio is less than 14, plants are limited by nitrogen. When the N/P ratio is 14–16, plants were limited by nitrogen and phosphorus. When the TN/TP ratio is greater than 16, plants are restricted by phosphorus. In this study area, the TC/TN of C. oleifera leaves is the greater than 16 under LL and ML, which indicates that plants are restricted by TP. Under CK and HL, TN/TP was 14–16, and plant growth are limited by TN and TP nutrient. The TC/TN of C. oleifera leaves in this study area was greater than 16 under LL and ML, which indicates that the plant was restricted by TP. The TN/TP ratio of the plants was 14–16 under CK and HL, which indicates that plant growth was limited by TN and TP. Nitrogen is the main nutrient element necessary for chlorophyll biosynthesis, and phosphorus also participates in the transformation of photosynthesis intermediates and energy transfer, so it has a great influence on photosynthesis. The energy required for nitrogen and phosphorus absorption and transport is ultimately provided by photosynthesis. Under LL and ML, phosphorus stress affects the activity of enzymes in photosynthesis and the regeneration of RuBP, and the demand for photosynthetic products is weakened. The contents of TC, TN, and TP were higher under HL, which indicates that C. oleifera leaves focused on storage metabolism and nutrient storage, which promoted the growth of twigs and leaves, flowering and fruiting, photosynthesis, and nitrogen metabolism, which is also beneficial to fruit growth.
4.5. Twigs Size
According to the LES theory [50] and “Corner’s rules”, plant species with higher SLA and leaf N and P contents have a higher photosynthesis rate and quicker growth. Therefore, C. oleifera belongs to the “slow investment-return” type. Our finding is that C. oleifera twigs have different adaptive strategies to changes in light regimes [51]. C. oleifera branches and fruits were also better developed under CK and HL (Table 5; Figure 8). Obviously, under LL and ML, the twigs of C. oleifera was thin, the leaves were found on the twigs, and the number of fruits was close to zero. Therefore, our results conform to Corner’s rules [52], and reflect plant adaptation strategies to different light regimes from the perspectives of the response mechanism of twigs and leaf functional traits, and the trade-off relationship [53,54].
The results of Ma [55] showed that under the condition of 60% shading, ‘Cenruan 3’ and ‘Gan 190’ could normally carry out flower bud differentiation and fruit setting, but with ‘Gan 190’ fruit setting was inhibited, while ‘Xiang 190’ was also inhibited. The number of flower buds and fruit set in Lin 86’ was significantly reduced; under 75% and 90% shading, the flower bud differentiation and fruit set of different varieties were significantly inhibited. The results of He [56] showed that Cenruan No. 3, Cenruan No. 22, Cenruan No. 24, Changlin No. 166, and Changlin No. 4 had moderate shade tolerance, and that Changlin No. 4 had the worst shade tolerance. Therefore, the current main cultivars of C. oleifera have a wide range of light energy utilization efficiency and can adapt well to strong light and weak light environments, but their potential net photosynthetic rate is low. Identifying and screening excellent C. oleifera varieties with high photosynthetic rate is an important direction for C. oleifera breeding with high light efficiency.
4.6. Evaluation of the Plasticity of C. oleifera Leaves under Different Light Regimes
We found that the external morphology, leaf anatomy, stomatal structure, photosynthetic parameters, chlorophyll content, nutrient content, and twigs size plasticity parameters of C. oleifera had different degrees of response to different light regimes (Figure 9). Among them, the factors were ranked as follows: the average plasticity index of photosynthetic parameters of C. oleifera leaves (0.38) > the average plasticity index of chlorophyll (0.31) > the average plasticity index of twigs size (0.26) > the average anatomical structure (0.24) and the leaf stomatal plasticity index (0.24) > the average plasticity index of the nutrient content (0.17) and leaf morphology index mean plasticity index (0.17). This indicates that the growth process of C. oleifera does not completely depend on the growth environment and other indicators of leaf morphological changes that adapt to changes in light, while the plasticity of photosynthetic parameters plays an important role in adapting to changes, further indicating that C. oleifera with higher light energy utilization efficiency is selected during the growth process, to promote growth under different habitats [57,58].
In this study, certain differences in external morphology, leaf anatomical structure, stomatal structure, photosynthetic parameters, chlorophyll content, nutrient content and twigs size were focused upon in C. oleifera leaves under four different light regimes. The results showed that leaves had greater thickness and increased chlorophyll content under HL [59]. In addition, more light energy was captured and fixed, and more biomass is available for photosynthesis in leaves under HL. Moreover, HL leaves had higher SLA, LB, and Pmax. In comparison, the twigs bore little fruit under LL and ML. In conclusion, the PT, ST, LA, SD, and chlorophyll contents of C. oleifera leaves changed under different light regimes. The HL was the most suitable regime for the growth of C. oleifera and has a high carbon assimilation ability and strong photosynthetic potential [60].
5. Conclusions
In this paper, five PI of C oleifera leaves under four light regimes, namely morphology, physiology, function, and twig size, were studied. The different response and adaptation strategies of C. oleifera to changing light regimes were preliminarily clarified. C. oleifera grew better and showed higher leaf biomass, twig biomass, and fruit biomass under HL. C oleifera increases its leaf area for photosynthesis by changing its own morphology, and invests more photosynthates in leaf parts, to enhance its light capture ability, light competition ability, and survival suitability. Therefore, C. oleifera requires strong light regimes for growth. The regulation of leaf physiological indexes (especially photosynthetic pigments) plays a key role in the adaptation of C. oleifera to different light regimes [61,62]. However, low light regimes (LL and ML) were not conducive to the fruit development of C. oleifera. Therefore insufficient light may be the main limiting factor for the regeneration and reproductive growth of C oleifera understory.
Author Contributions
Y.Z. performed date curation, formal analysis, investigation, initial draft Q.G. planned and supervised the project, conceived and designed the experiments, funding acquisition, writing-censor-editing; S.L., J.P. and S.Y. performed investigation C.G. and G.W. conducted experimental guidance; Y.G. performed supervision, writing-censor-editing. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by Technological Projects of Guizhou Province, China ([2018]5261), China Scholarship Council ([2021]15), Platform Talent Program of Guizhou Province, China ([2019]5643), and the Construction Project for Firs-class Ecology Discipline in Guizhou (GNYL [2017] 007).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tu, J.; Chen, J.; Zhou, J.; Ai, W.; Chen, L. Plantation quality assessment of Camellia oleifera in mid-subtropical China. Soil Tillage Res. 2019, 186, 249–258. [Google Scholar] [CrossRef]
- Li, N.N.; Yang, Y.P.; Ye, J.H.; Liu, J.L.; Zheng, X.P.; Liang, Y.R. Effects of sunlight on gene expression and chemical composition of light-sensitive albino tea plant. Plant Growth Regul. 2016, 78, 253–262. [Google Scholar] [CrossRef]
- Gao, C.; Yuan, D.Y.; Wang, B.F.; Yang, Y.; Liu, D.M.; Han, Z.Q. A cytological study of anther and pollen development in Camellia oleifera. GMR 2015, 14, 8755–8765. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Zhang, Y.; Su, S.; Yang, S.; Wang, X. Effects of tree shape on the microclimate and fruit quality parameters of Camellia oleifera Abel. Forests 2019, 10, 563. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Carriquí, M. Photosynthesis and photosynthetic efficiencies along the terrestrial plant’s phylogeny: Lessons for improving crop photosynthesis. Plant J. 2020, 101, 964–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.L.; Huang, M.Y.; Chen, C.W.; Weng, J.H. Light induction of nonphotochemical quenching, CO2 fixation, and photoinhibition in woody and fern species adapted to different light regimes. Photosynthetica 2014, 52, 272–280. [Google Scholar] [CrossRef]
- Gao, C.; Yang, R.; Yuan, D.Y. Structural characteristics of the mature embryosac of Camellia oleifera. Nord. J. Bot. 2017, 36, 1–13. [Google Scholar]
- Wei, H.L.; Gao, C.; Qiu, J.; Long, L.; Wang, B.; Yang, L.; Hu, Y. Flowering Biological Characteristics of Camellia weiningensis Y.K. Li. J. Am. Soc. Hortic. Sci. 2021, 56, 1331–1339. [Google Scholar] [CrossRef]
- Yu, Z.C.; Luo, Y.N.; Shi, W.Y.; Lin, W.; Peng, C.L. Adaptability variations and differences in photoprotection of young leaves of two tree species of subtropical forest to the light environment. Photosynthetica 2021, 9, 116–126. [Google Scholar] [CrossRef]
- Liu, C.; Jin, G.; Liu, Z. Importance of organ age in driving intraspecific trait variation and coordination for three evergreen coniferous species. Ecol. Indic. 2020, 121, 107099. [Google Scholar] [CrossRef]
- Guo, Q.Q.; Li, H.E.; Yang, R. Leaf traits and photosynthetic characteristics of endangered Sinopodophyllum hexandrum (Royle) Ying under different light regimes in Southeastern Tibet Plateau. Photosynthetica 2019, 57, 548–555. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Huang, Z.; Wang, Z.; Chen, Y.; Tigabu, M. Responses of leaf morphology, NSCs contents and C: N: P stoichiometry of Cunninghamia lanceolate and Schima superba to shading. BMC Plant Biol. 2021, 20, 354. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Wright, S.J.; Lasso, E.; Kitajima, K.; Pearcy, R.W. Plastic phenotypic response to light of 16 congeneric shrubs from a panamanian rainforest. Ecology 2000, 81, 1925–1936. [Google Scholar] [CrossRef]
- Wen, Y.; Su, S.C.; Jia, T.T.; Wang, X.N. Allocation of photoassimilates in bud and fruit from different leaf nodes of Camellia oleifera. J. Am. Soc. Hortic. Sci. 2021, 56, 1–9. [Google Scholar] [CrossRef]
- Cheng, X.; Ping, T.; Li, Z.; Wang, T.; Epstein, H.E. Effects of environmental factors on plant functional traits across different plant life forms in a temperate forest ecosystem. New For. 2021, 53, 125–142. [Google Scholar] [CrossRef]
- Liu, C.X.; He, Z.L.; Chen, Y.Z.; Tang, W.; Ma, L.; Chen, L.S. Codon usage patterns analysis of Camellia oleifera based on transcriptome data. J. Biobased Mater. Bioenergy 2021, 15, 347–352. [Google Scholar] [CrossRef]
- Wang, D.; Huang, X.L.; Chen, J.; Li, L.X.; Liu, J.M. Plasticity of leaf traits of Juglans regia L. f. luodianense Liu et Xu seedlings under different light conditions in Karst habitats. Forests 2021, 12, 81. [Google Scholar]
- Mlinaric, S.; Pfeiffer, T.; Krstin, L.; Maronic, D.; Varga, M. Adaptation of Amorpha fruticosa to different habitats is enabled by photosynthetic apparatus plasticity. Photosynthetica 2021, 59, 137–147. [Google Scholar] [CrossRef]
- Stotz, G.C.; Salgado, L.C.; Escobedo, V.M.; Valladares, F.; Gianoli, E. Global trends in phenotypic plasticity of plants. Ecol. Lett. 2021, 24, 2267–2281. [Google Scholar] [CrossRef]
- Blondeel, H.; Perring, M.P.; Depauw, L.; Lombaerde, E.D.; Landuyt, D.; Frenne, P.D.; Verheyen, K. Light and warming drive forest understory community development in different environments. Glob. Change Biol. 2020, 26, 1681–1696. [Google Scholar] [CrossRef]
- Marenco, R.A.; Camargo, M.; Antezana-Vera, S.A.; Oliveira, M.F. Leaf trait plasticity in six forest tree species of central Amazonia. Photosynthetica 2017, 55, 679–688. [Google Scholar] [CrossRef]
- Tsialtas, J.T.; Maslaris, N. Leaf allometry and prediction of specific leaf area (SLA) in a sugar beet (Beta vulgaris L.) cultivar. Photosynthetica 2008, 46, 351–355. [Google Scholar] [CrossRef]
- Kholmanskiy, A.; Zaytseva, N. Dependence of chlorophyll content in leaves from light regime, electromagnetic fields and plant species. JOJ Hortic. Arboric. 2020, 3, 555602. [Google Scholar] [CrossRef]
- Antonielli, M.; Pasqualini, S.; Batini, P.; Ederli, L.; Massacci, A.; Loreto, F. Physiological and anatomical characterisation of Phragmites australis leaves. Aquat. Bot. 2002, 72, 55–66. [Google Scholar] [CrossRef]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef] [Green Version]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Xu, C.Y. Response and Adaptation of Tilia Seedlings and Young Trees to Light; Beijing Forestry University: Beijing, China, 1999; pp. 20–21. [Google Scholar]
- Gonzalez-Paleo, L.; Ravetta, D.A. Relationship between photosynthetic rate, water use and leaf structure in desert annual and perennial forbs differing in their growth. Photosynthetica 2018, 56, 1177–1187. [Google Scholar] [CrossRef]
- Fischer, A.H.; Jacobson, K.A.; Rose, J.; Zeller, R. Paraffin embedding tissue samples for sectioning. CSH Protoc. 2008, 3, 4989. [Google Scholar] [CrossRef]
- Horiguchi, G.; Fujikura, U.; Ferjani, A.; Ishikawa, N.; Tsukaya, H. Large-scale histological analysis of leaf mutants using two simple leaf observation methods: Identification of novel genetic pathways governing the size and shape of leaves. Plant J. Cell Mol. Biol. 2010, 48, 638–644. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzym. 1987, 148, 350–382. [Google Scholar]
- Bao, S. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Silva, E.N.; Ferreira-Silva, S.L.; de Vasconcelos Fontenele, A.; Ribeiro, R.V.; Viégas, R.A.; Silveira, J.A.G. Photosynthetic changes and protective mechanisms against oxidative damage subjected to isolated and combined drought and heat stresses in Jatropha curcas plants. J. Plant Physiol. 2010, 167, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Thornley, J.H.M. Mathematical Models in Plant Physiology; Academic Press: London, UK, 1976. [Google Scholar]
- Han, C.J.; Wang, Q.; Zhang, H.B.; Wang, S.H.; Song, H.D.; Hao, J.M.; Dong, H.Z. Light shading improves the yield and quality of seed in oil-seed peony (Paeonia ostii Feng Dan). J. Integr. Agric. 2018, 17, 1631–1640. [Google Scholar] [CrossRef]
- An, H.; Shangguan, Z.P. Specific leaf area, leaf nitrogen content, and photosynthetic acclimation of Trifolium repens L. seedlings grown at different irradiances and nitrogen concentrations. Photosynthetica 2008, 46, 143–147. [Google Scholar] [CrossRef]
- Walcroft, A.; Le Roux, X.; Diaz-Espejo, A.; Dones, N.; Sinoquet, H. Effects of crown development on leaf irradiance, leaf morphology and photosynthetic capacity in a peach tree. Tree Physiol. 2002, 22, 929–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Feng, S.L.; Chen, T.; Zhou, L.J.; Yuan, M.; Liao, J.Q.; Huang, Y.; Yang, H.Y.; Yang, R.W.; Ding, C.B. Quality assessment of Camellia oleifera oil cultivated in Southwest China. Separations 2021, 8, 144. [Google Scholar] [CrossRef]
- Lin, X.Y.; Peng, Q.F.; Lü, H.F.; Du, Y.Q.; Tang, B.Y. Leaf anatomy of Camellia sect. Oleifera and sect. Paracamellia (Theaceae) with reference to their taxonomic significance. J. Syst. Evol. 2008, 46, 183–193. [Google Scholar]
- Zhai, J.; Li, Y.; Han, Z.; Li, Z. Morphological, structural and physiological differences in heteromorphic leaves of Euphrates poplar during development stages and at crown scales. Plant Biol. 2020, 22, 366–375. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Feng, Q.; Si, J.H.; Chang, Z.Q.; Cheng, K.L.; Cao, G.C. Relationships of photosynthesis and transpiration of Populus euphratica with their affecting factors. J. Arid. Land Resour. Environ. 2012, 26, 155–159. [Google Scholar]
- Luo, Q.; Xu, C.; Zheng, T.; Ma, Y.; Li, Y.; Zuo, Z. Leaf morphological and photosynthetic differences among four chemotypes of Cinnamomum camphora in different seasons. Ind. Crop. Prod. 2021, 169, 113651. [Google Scholar] [CrossRef]
- Zhang, Z.; Tao, S.; Zhou, B.; Zhang, X.; Zhao, Z. Plant stomatal conductance determined transpiration and photosynthesis both contribute to the enhanced negative air ion (NAI). Ecol. Indic. 2021, 130, 108114. [Google Scholar] [CrossRef]
- Yan, Z.; Huang, L.; Wei, X.; Zhou, H.; Xun, C. Physiological, morphological, and anatomical changes in Rhododendron agastum in response to shading. Plant Growth Regul. 2016, 81, 23–30. [Google Scholar]
- Stefan, H.; Urs, F. Nitrogen metabolism and remobilization during senescence. J. Exp. Bot. 2002, 53, 927–937. [Google Scholar]
- Zhang, X.R.; Tan, X.F.; Wang, R.Q.; Xu, N.N.; Guo, W.H. Effects of soil moisture and light intensity on ecophysiological characteristics of Amorpha fruticosa seedlings. J. For. Res. 2013, 24, 293–300. [Google Scholar] [CrossRef]
- Walters, R.G.; Horton, P. Acclimation of Arabidopsis thaliana to the light environment: Changes in photosynthetic function. Planta 1995, 197, 306–317. [Google Scholar] [CrossRef]
- Laurel, C.; Walls, R.L.; Justin, E.; Gandolfo, M.A.; Stevenson, D.W.; Barry, S.; Justin, P.; Balaji, A.; Mungall, C.J.; Stefan, R. The plant ontology as a tool for comparative plant anatomy and genomic analyses. Plant Cell Physiol. 2012, 2, 2. [Google Scholar]
- Han, W.X.; Fang, J.Y.; Guo, D.L. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef]
- Wright, J.P.; Sutton-Grier, A.; Stevens, C. Does the leaf economic spectrum hold within local species pools across varying environmental conditions? Funct. Ecol. 2012, 26, 1380–1398. [Google Scholar] [CrossRef]
- Díaz, S.; Kattge, J.; Cornelissen, J.; Wright, I.J.; Gorné, L.D. The global spectrum of plant form and function. Nature 2015, 529, 167–183. [Google Scholar] [CrossRef]
- Sun, S.; Jin, D.; Shi, P. The Leaf Size-Twig size spectrum of temperate woody species along an altitudinal gradient: An invariant allometric scaling relationship. Ann. Bot. 2006, 97, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Cai, H. Allometric models for leaf area and leaf mass predictions across different growing seasons of elm tree (Ulmus japonica). J. For. Res. 2017, 28, 975–982. [Google Scholar] [CrossRef]
- Reich, P.B.; Cornelissen, H. The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- MA, J.L. Research on Physiological Properties of Low-Light Tolerance in Oil-Tea Camellia Plantation; Central South University of Forestry and Technology: Changsha, China, 2012; pp. 18–26. [Google Scholar]
- He, X.Y. Effect of Low Light Stress on Photosynthetic and Growth Properties in Near-Mature Oil-Tea Camellia Plantation; Central South University of Forestry and Technology: Changsha, China, 2012; pp. 56–58. [Google Scholar]
- Kusi, J.; Karsai, I. Plastic leaf morphology in three species of Quercus: The more exposed leaves are smaller, more lobated and denser. Plant Species Biol. 2020, 35, 24–37. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Viña, A.; Ciganda, V.; Rundquist, D.C.; Arkebauer, T.J. Remote estimation of canopy chlorophyll content in crops. Geophys. Res. Lett. 2005, 32, L08403. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Shabala, L.; Zhou, M.; Brodribb, T.J.; Corkrey, R.; Shabala, S. Factors determining stomatal and non-stomatal (residual) transpiration and their contribution towards salinity tolerance in contrasting barley genotypes. Environ. Exp. Bot. 2018, 153, 10–20. [Google Scholar] [CrossRef]
- Hörtensteiner, S. Chlorophyll breakdown in higher plants and algae. Cell Mol. Life Sci. 1999, 56, 330. [Google Scholar] [CrossRef]
- Urbas, P.; Zobel, K. Adaptive and inevitable morphological plasticity of three herbaceous species in a multi-species community: Field experiment with manipulated nutrients and light. Acta Oecol. 2000, 21, 139–147. [Google Scholar] [CrossRef]
- Shipley, B.; Lechowicz, M.J.; Wright, I.; Reich, P.B. Fundamental trade-offs generating the worldwide leaf economics spectrum. Ecology 2006, 87, 535–541. [Google Scholar] [CrossRef]
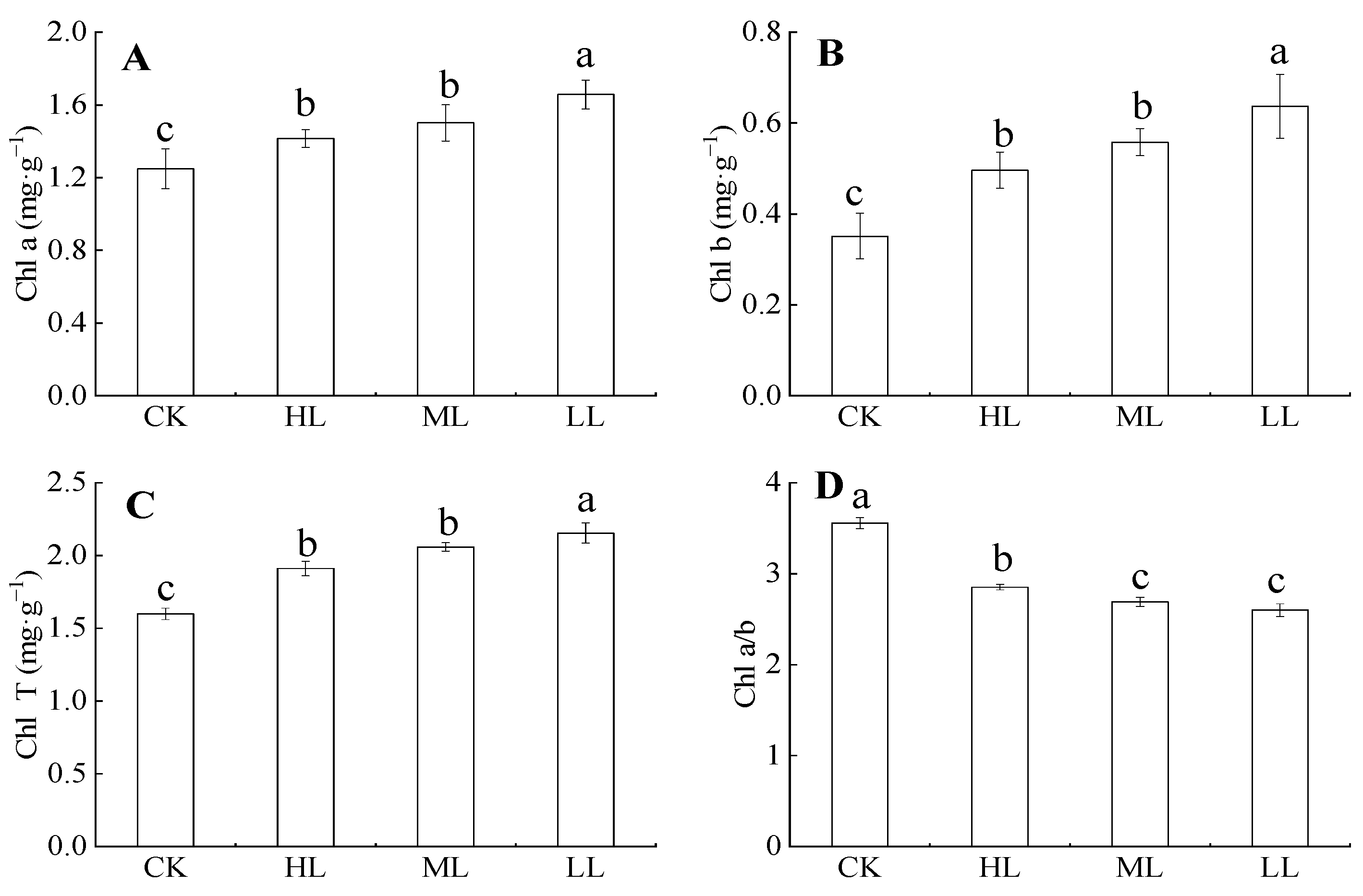

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).