
Disentangling the Interspecific and Intraspecific Variation in Functional Traits of Desert Plant Communities under Different Moisture Gradients

Abstract
:1. Introduction
2. Materials and Methods
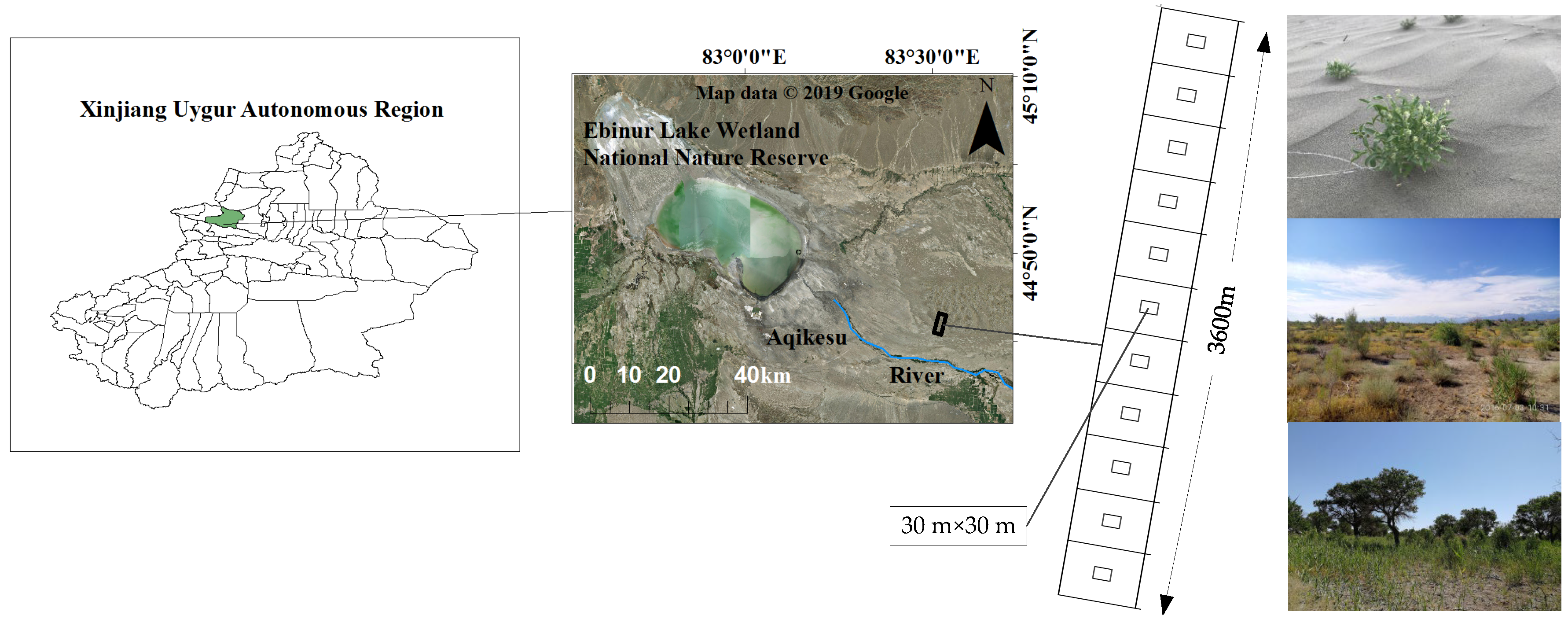
2.1. Overview of Experimental Area
2.2. Research Methods
2.2.1. Sample Setting
2.2.2. Collection of Soil Samples
2.2.3. Collection of Plant Samples
2.3. Data Processing and Analysis
2.3.1. Division of Different Soil Moisture Gradients
2.3.2. Calculation of CWM
2.3.3. Data Analysis
3. Results and Analysis
3.1. Difference Analysis of Soil Factors and Plant Functional Traits under Different Water Gradients
3.2. Inter- and Intraspecific Variation in Plant Traits and Their Differences According to Water Gradient
3.3. Relationship between Inter- and Intraspecific Variation in Plant Functional Traits and Soil Physicochemical Factors
3.4. Contribution of Inter- and Intraspecific Variation to Community Weighting
4. Discussion
4.1. Analysis of Inter- and Intraspecific Variation in Plant Functional Traits and Response to Community Construction
4.2. Effects of Soil Environment on Inter- and Intraspecific Variation in Community Functional Traits
4.3. Contribution and Importance of Inter- and Intraspecific Variation to Community Weighting
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xiong, M.H. The Association and Differentiation of Plant Functional Traits in Tropical Cloud Forest in Hainan. Master’s Thesis, Hainan University, Seaport, China, 2015. [Google Scholar]
- Guo, A.; Zuo, X.; Zhang, S.; Hu, Y.; Yue, P.; Lv, P.; Li, X.; Zhao, S.; Yu, Q. Contrasting effects of plant inter- and intraspecific variation on community trait responses to nitrogen addition and drought in typical and meadow steppes. BMC Plant Biol. 2022, 22, 1471–2229. [Google Scholar] [CrossRef]
- Li, Y.Z. Intraspecific Trait Variability at Multiple Spatial Scales and Community Assembly in a Subalpine Meadow. Master’s Thesis, Lanzhou University, Lanzhou, China, 2013. [Google Scholar]
- Ackerly, D.D.; Cornwell, W.K. A trait-based approach to community assembly: Partitioning of species trait values into within-and among-community components. Ecol. Lett. 2007, 10, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Albert, C.H.; Thuiller, W.; Yoccoz, N.G.; Soudant, A.; Boucher, F.; Saccone, P.; Lavorel, S. Intraspecific functional variability: Extent, structure and sources of variation. J. Ecol. 2010, 98, 604–613. [Google Scholar] [CrossRef]
- Albert, C.H.; Thuiller, W.; Yoccoz, N.G.; Douzet, R.; Aubert, S.; Lavorel, S. A multi-trait approach reveals the structure and the relative importance of intra- vs. interspecific variability in plant traits. Funct. Ecol. 2010, 24, 1192–1201. [Google Scholar] [CrossRef]
- Jung, V.; Albert, C.H.; Violle, C.; Kunstler, G.; Loucougaray, G.; Spiegelberger, T. Intraspecific trait variability mediates the response of subalpine grassland communities to extreme drought events. J. Ecol. 2013, 102, 45–53. [Google Scholar] [CrossRef]
- Plourde, B.T.; Boukili, V.K.; Chazdon, R.L. Radial changes in wood specific gravity of tropical trees: Interand intraspecific variation during secondary succession. Funct. Ecol. 2015, 29, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Jung, V.; Violle, C.; Mondy, C.; Hoffmann, L.; Muller, S.D. Intraspecific variability and trait-based community assembly. J. Ecol. 2010, 98, 1134–1140. [Google Scholar] [CrossRef]
- Paine, C.; Baraloto, C.; Chave, J.; Hérault, B. Functional traits of individual trees reveal ecological constraints on community assembly in tropical rain forests. Oikos 2011, 120, 720–727. [Google Scholar] [CrossRef]
- Siefert, A. Incorporating intraspecific variation in tests of trait-based community assembly. Oecologia 2012, 170, 767–775. [Google Scholar] [CrossRef]
- Cianciaruso, M.V.; Batalha, M.A. Short-term community dynamics in seasonal and hyperseasonal cerrados. Braz. J. Biol. 2009, 69, 631–637. [Google Scholar] [CrossRef] [Green Version]
- De Bello, F.; Lavorel, S.; Albert, C.H.; Thuiller, W.; Grigulis, K.; Dolezal, J.; Janeček, Š.; Lepš, J. Quantifying the relevance of intraspecific trait variability for functional diversity. Methods Ecol. Evol. 2011, 2, 163–174. [Google Scholar] [CrossRef]
- Clark, J.S.; Bell, D.M.; Hersh, M.H.; Kwit, M.C.; Moran, E.; Salk, C.; Stine, A.; Valle, D.; Zhu, K. Individual-scale variation, species-scale differences: Inference needed to understand diversity. Ecol. Lett. 2011, 14, 1273–1287. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, D.C.; Joshi, C.; van Bodegom, P.M.; Bastow, Z.A.; Fulé, P.Z. A predictive model of community assembly that incorporates intraspecific trait variation. Ecol. Lett. 2012, 15, 1291–1299. [Google Scholar] [CrossRef]
- Hulshof, C.M.; Violle, C.; Spasojevic, M.J.; McGill, B.; Damschen, E.; Harrison, S.; Enquist, B.J. Intra-specific and inter-specific variation in specific leaf area reveal the importance of abiotic and biotic drivers of species diversity across elevation and latitude. J. Veg. Sci. 2013, 24, 921–931. [Google Scholar] [CrossRef]
- Benavides, R.; Carvalho, B.; Bastias, C.C.; López-Quiroga, D.; Mas, A.; Cavers, S.; Gray, A.; Albet, A.; Alía, R.; Ambrosio, O.; et al. The GenTree Leaf Collection: Inter- and intraspecific leaf variation in seven forest tree species in Europe. Glob. Ecol. Biogeogr. 2021, 30, 590–597. [Google Scholar] [CrossRef]
- Violle, C.; Enquist, B.J.; McGill, B.J.; Jiang, L.; Albert, C.H.; Hulshof, C.; Jung, V.; Messier, J. The return of the variance: Intraspecific variability in community ecology. Trends Ecol. Evol. 2012, 27, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Q.; Huang, Y.T.; Ding, Y.; Zang, R.G. Interspecific and intraspecific variations in plant functional traits in subtropical evergreen and deciduous broad-leaved mixed forests. Biodiversity 2016, 24, 262–270. [Google Scholar]
- Jackson, B.G.; Peltzer, D.A.; Wardle, D.A. The within-species leaf economic spectrum does not predict leaf litter decomposability at either the within-species or whole community levels. J. Ecol. 2013, 101, 1409–1419. [Google Scholar] [CrossRef] [Green Version]
- Asplund, J.; Wardle, D.A. Within-species variability is the main driver of community-level responses of traits of epiphytes across a long-term chronosequence. Funct. Ecol. 2014, 28, 1513–1522. [Google Scholar] [CrossRef] [Green Version]
- Laforest-Lapointe, I.; Martínez-Vilalta, J.; Retana, J. Intraspecific variability in functional traits matters: Case study of Scots pine. Oecologia 2014, 175, 1337–1348. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Yao, Y.P.; Yao, Y.P.; Jiang, Y.; Liang, S.C.; Li, Y.J.; Liang, H.H.; Zhao, Q.N.; Huang, Y.B.; Ling, C.J. Interspecific and intraspecific variations in plant functional traits of Cyclobalanopsis glauca community in karst rocky mountain of Guilin. Acta Ecol. Sin. 2021, 41, 8237–8245. [Google Scholar]
- Li, Y.Z. Study on Intraspecific Variation and Community Construction Mechanism of Subalpine Meadow at Different Spatial Scales. Master’s Thesis, Lanzhou University, Lanzhou, China, 2013. [Google Scholar]
- Yan, B.G. Study on Plant Community Combination Mechanism of Alpine Forest—Grass Transition Zone in Western Sichuan. Master’s Thesis, Sichuan Agricultural University, Chengdu, China, 2010. [Google Scholar]
- Price, J.N.; Gazol, A.; Tamme, R.; Hiiesalu, I.; Pärtel, M. The functional assembly of experimental grasslands in relation to fertility and resource heterogeneity. Funct. Ecol. 2014, 28, 509–519. [Google Scholar] [CrossRef]
- Pausas, J.G.; Austin, M.P. Patterns of plant species richness in relation to different environments: An appraisal. J. Veg. Sci. 2001, 12, 153–166. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, C.Z.; Yao, W.X.; Wang, J.L.; Zhang, W.T. Response of the relationship between the transpiration rate of reed and leaf traits in Zhangye wetland to soil moisture. J. Ecol. 2018, 37, 1095–1101. [Google Scholar]
- Reich, P.B. The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Wilson, P.J.; Thompson, K.; Hodgson, J.G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytol. 1999, 143, 155–162. [Google Scholar] [CrossRef]
- Moles, A.T.; Perkins, S.E.; Laffan, S.W.; Flores-Moreno, H.; Awasthy, M.; Tindall, M.L.; Sack, L.; Pitman, A.; Kattge, J.; Aarssen, L.W.; et al. Which is a better predictor of plant traits: Temperature or precipitation? J. Veg. Sci. 2004, 25, 1167–1180. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, Y.; Wei, W.W. Changes of leaf traits of typical shrub and grass along climate gradient in northwest arid region. Ecol. J. 2021, 40, 3769–3777. [Google Scholar]
- Chalmandrier, L.; Münkemüller, T.; Colace, M.P.; Renaud, J.; Aubert, S.; Carlson, B.Z.; Clément, J.-C.; Legay, N.; Pellet, G.; Saillard, A.; et al. Spatial scale and intraspecific trait variability mediate assembly rules in alpine grasslands. J. Ecol. 2017, 105, 277–287. [Google Scholar] [CrossRef] [Green Version]
- Niu, K.; Zhang, S.; Lechowicz, M.J. Harsh environmental regimes increase the functional significance of intraspecific variation in plant communities. Funct. Ecol. 2020, 34, 1666–1677. [Google Scholar] [CrossRef]
- Wang, X.; Yang, L.; Zhao, Q.; Zhang, Q.D. Responses of grassland community functional traits to soil moisture in typical small watersheds of the Loess Plateau. Ecology 2020, 40, 2691–2697. [Google Scholar]
- Wang, H.F. Study on Plant Diversity and Ecosystem Function in Ebinur Lake Basin. Ph.D. Thesis, Xinjiang University, Urumqi, China, 2020. [Google Scholar]
- Xinjiang Comprehensive Investigation Team; Chinese Academy of Sciences. Vegetation and Its Utilization in Xinjiang; Science Press: Beijing, China, 1978. [Google Scholar]
- Yang, X.D.; Lv, G.H.; Tian, Y.H.; Yang, J.; Zhang, X.M. The ecological grouping of plants in Xinjiang Ebinur Lake Wetland Nature Reserve. Ecol. J. 2009, 28, 2489–2494. [Google Scholar]
- Bao, S.D. Soil Agrochemical Analysis, 3rd ed.; China Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Guan, S.Y. Soil Enzyme and Its Research Method; Agricultural Press: Beijing, China, 1986. [Google Scholar]
- Zhang, X.N.; Yang, X.D.; Lv, G.H. Diversity pattern of desert plants under water-salt gradient and its relationship with soil environment. Ecology 2016, 36, 3206–3215. [Google Scholar]
- Lepš, J.; de Bello, F.; Šmilauer, P.; Doležal, J. Community trait response to environment: Disentanglig species turnover vs. intraspecific trait variability effects. Ecography 2011, 4, 856–863. [Google Scholar] [CrossRef]
- Taiyun, W.; Viliam, S. R Package ‘Corrplot’: Visualization of a Correlation Matrix (Version 0.92). 2021. Available online: https://github.com/taiyun/corrplot (accessed on 28 March 2022).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-7. 2020. Available online: https://CRAN.Rproject.org/package=vegan (accessed on 28 March 2022).
- Berg, M.P.; Ellers, J. Trait plasticity in species interactions: A driving force of community dynamics. Evol. Ecol. 2010, 24, 617–629. [Google Scholar] [CrossRef] [Green Version]
- Lichstein, J.W.; Dushoff, J.; Levin, S.A.; Pacala, S.W. Intraspecific Variation and Species Coexistence. Am. Nat. 2007, 170, 807–818. [Google Scholar] [CrossRef]
- Bolnick, D.I.; Amarasekare, P.; Araújo, M.S.; Bürger, R.; Levine, J.M.; Novak, M.; Rudolf, V.H.; Schreiber, S.J.; Urban, M.C.; Vasseur, D.A. Why intraspecific trait variation matters in community ecology. Trends Ecol. Evol. 2011, 26, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef] [Green Version]
- Xun, Y.H.; Di, X.Y.; Jin, G.Z. Vertical variation and economic strategy of leaf traits of main tree species in typical broad-leaved Korean pine forest. Plant Ecol. J. 2020, 44, 730–741. [Google Scholar] [CrossRef]
- Ma, H.Y.; Yang, X.D.; Lv, G.H.; He, X.M.; Zhang, X.N.; Wang, X.Y.; Li, Y. Water Sources in Dominant Species of Desert in Ebinur Lake Wetland Nature Reserve, Xinjiang. Ecol. J. 2017, 37, 829–840. [Google Scholar]
- Lajoie, V.M. Understanding context dependence in the contribution of intraspecific variation to community trait-environment matching. Ecology 2015, 96, 2912–2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, P.B.; Wright, I.J.; Cavender-Bares, J.; Craine, J.M.; Oleksyn, J.; Westoby, M.; Walters, M.B. The Evolution of Plant Functional Variation: Traits, Spectra, and Strategies. Int. J. Plant Sci. 2003, 164, 143–164. [Google Scholar] [CrossRef]
- Li, Y.Q.; Wang, Z.H. Ecological function, geographical distribution and genesis of plant leaf morphology. J. Plant Ecol. 2021, 45, 1154–1172. [Google Scholar] [CrossRef]
- Li, J.X.; Xu, W.T.; Xiong, G.M.; Wang, Y.; Zhang, C.M.; Lu, Z.J.; Li, Y.L.; Xie, Z.Q. Contents of nitrogen and phosphorus in leaves of dominant woody shrubs in southern China and their influencing factors. J. Plant Ecol. 2017, 41, 31–42. [Google Scholar]
- He, D. Variation of Plant Functional Traits and Community Construction. Ph.D. Thesis, Sun Yat-sen University, Guangzhou, China, 2016. [Google Scholar]
- Zhang, R.Y.; Li, Y.P.; Ni, Y.L.; Gui, X.; Lian, J.; Ye, W. Intraspecific variation of leaf functional traits of subtropical evergreen broad-leaved forest in Dinghu Mountain along the vertical level of community. Biodiversity 2019, 27, 1279–1290. [Google Scholar]
- Messier, J.; McGill, B.J.; Lechowicz, M.J. How do traits vary across ecological scales? A case for trait-based ecology: How do traits vary across ecological scales? Ecol. Lett. 2010, 13, 838–848. [Google Scholar] [CrossRef] [PubMed]
- Garnier, E.; Laurent, G.; Bellmann, A.; Debain, S.; Berthelier, P.; Ducout, B.; Roumet, C.; Navas, M.-L. Consistency of Species Ranking Based on Functional Leaf Traits. New Phytol. 2001, 152, 69–83. [Google Scholar] [CrossRef]
- Hulshof, C.M.; Swenson, N.G. Variation in leaf functional trait values within and across individuals and species: An example from a Costa Rican dry forest. Funct. Ecol. 2010, 24, 217–223. [Google Scholar] [CrossRef]
- Bao, L.; Liu, Y.H. Comparison of leaf functional traits among different forest communities in Dongling Mountain. Ecology 2009, 29, 3692–3703. [Google Scholar]
- Gratani, L.; Meneghini, M.; Pesoli, P.; Crescente, M.F. Structural and functional plasticity of Quercus ilex seedlings of different provenances in Italy. Trees 2003, 17, 515–521. [Google Scholar] [CrossRef]
- Lü, X.-T.; Hu, Y.-Y.; Zhang, H.-Y.; Wei, H.-W.; Hou, S.-L.; Yang, G.-J.; Liu, Z.-Y.; Wang, X.-B. Intraspecific variation drives community-level stoichiometric responses to nitrogen and water enrichment in a temperate steppe. Plant Soil 2018, 423, 307–315. [Google Scholar] [CrossRef]
- Sun, X.Y. Effects of Intraspecific Variation on Individual Growth of Woody Plants and Responses to Interspecific Relationships. Master’s Thesis, East China Normal University, Shanghai, China, 2016. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Factors | Method |
|---|---|
| SWC | Drying and weighing method |
| SSC | Residue method |
| pH | Acidimeter |
| SOC | Potassium dichromate dilution heating method |
| TN | Kjeldahl method |
| TP | Molybdenum antimony colorimetric method |
| Moisture | Number of Plot | Mean/% | Stand Deviation | Min/% | Max/% |
|---|---|---|---|---|---|
| High moisture gradient | 26 | 7.10 a | 4.72 | 3.58 | 21.21 |
| Low moisture gradient | 26 | 1.85 b | 1.09 | 0.29 | 3.51 |
| Functional Trait | Interspecific Variation | Intraspecific Variation | ||
|---|---|---|---|---|
| Low Moisture | High Moisture | Low Moisture | High Moisture | |
| LL | 9.047 ± 21.016 a | 12.215 ± 25.628 b | −7.767 ± 21.151 a | −9.033 ± 25.639 b |
| DBH/BD | 2.107 ± 2.677 a | 2.449 ± 2.633 a | −1.704 ± 2.684 a | −1.644 ± 2.726 a |
| H | 1.388 ±1.278 a | 1.590 ± 1.253 a | −1.074 ± 1.294 a | −1.192 ± 1.276 a |
| LW | 1.448 ± 1.916 a | 1.661 ± 2.13 a | −1.238 ± 1.927 a | −1.245 ± 2.156 a |
| LT | 0.315 ± 0.591 a | 0.387 ± 0.642 b | −0.235 ± 0.591 a | −0.273 ± 0.644 b |
| Soil Factor | High Moisture Gradient | Low Moisture Gradient | ||||
|---|---|---|---|---|---|---|
| Explanation % | Contribution % | p | Explanation % | Contribution % | p | |
| TN | 6.4 | 32.4 | 0.188 | 9.1 | 32.8 | 0.134 |
| TP | 8.3 | 42 | 0.122 | 1.3 | 4.6 | 0.588 |
| pH | 1.9 | 9.5 | 0.51 | 0.4 | 1.6 | 0.858 |
| SOC | 1.1 | 5.7 | 0.622 | 9.3 | 33.5 | 0.088 |
| SWC | 0.9 | 4.6 | 0.686 | 3.1 | 11.3 | 0.352 |
| SSC | 1.1 | 5.8 | 0.644 | 4.5 | 16.3 | 0.244 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.; Wang, H.; Cai, Y.; Yang, Q.; Chen, C.; Lv, G. Disentangling the Interspecific and Intraspecific Variation in Functional Traits of Desert Plant Communities under Different Moisture Gradients. Forests 2022, 13, 1088. https://doi.org/10.3390/f13071088
Sun L, Wang H, Cai Y, Yang Q, Chen C, Lv G. Disentangling the Interspecific and Intraspecific Variation in Functional Traits of Desert Plant Communities under Different Moisture Gradients. Forests. 2022; 13(7):1088. https://doi.org/10.3390/f13071088
Chicago/Turabian StyleSun, Li, Hengfang Wang, Yan Cai, Qi Yang, Caijin Chen, and Guanghui Lv. 2022. "Disentangling the Interspecific and Intraspecific Variation in Functional Traits of Desert Plant Communities under Different Moisture Gradients" Forests 13, no. 7: 1088. https://doi.org/10.3390/f13071088