
An Integrative Approach to Study How Driving Factors Control Biomass Carbon Density for Natural Mountain Forests, in China’s Loess Plateau

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Region
2.2. Data Collection and Preprocessing
2.2.1. Forest Inventory Data
2.2.2. Climate Data
2.3. Data Analyses
2.3.1. Statistical Analyses
2.3.2. Spatial Hot and Cold Spots Analyses
2.3.3. Multi-Group SEM
3. Results
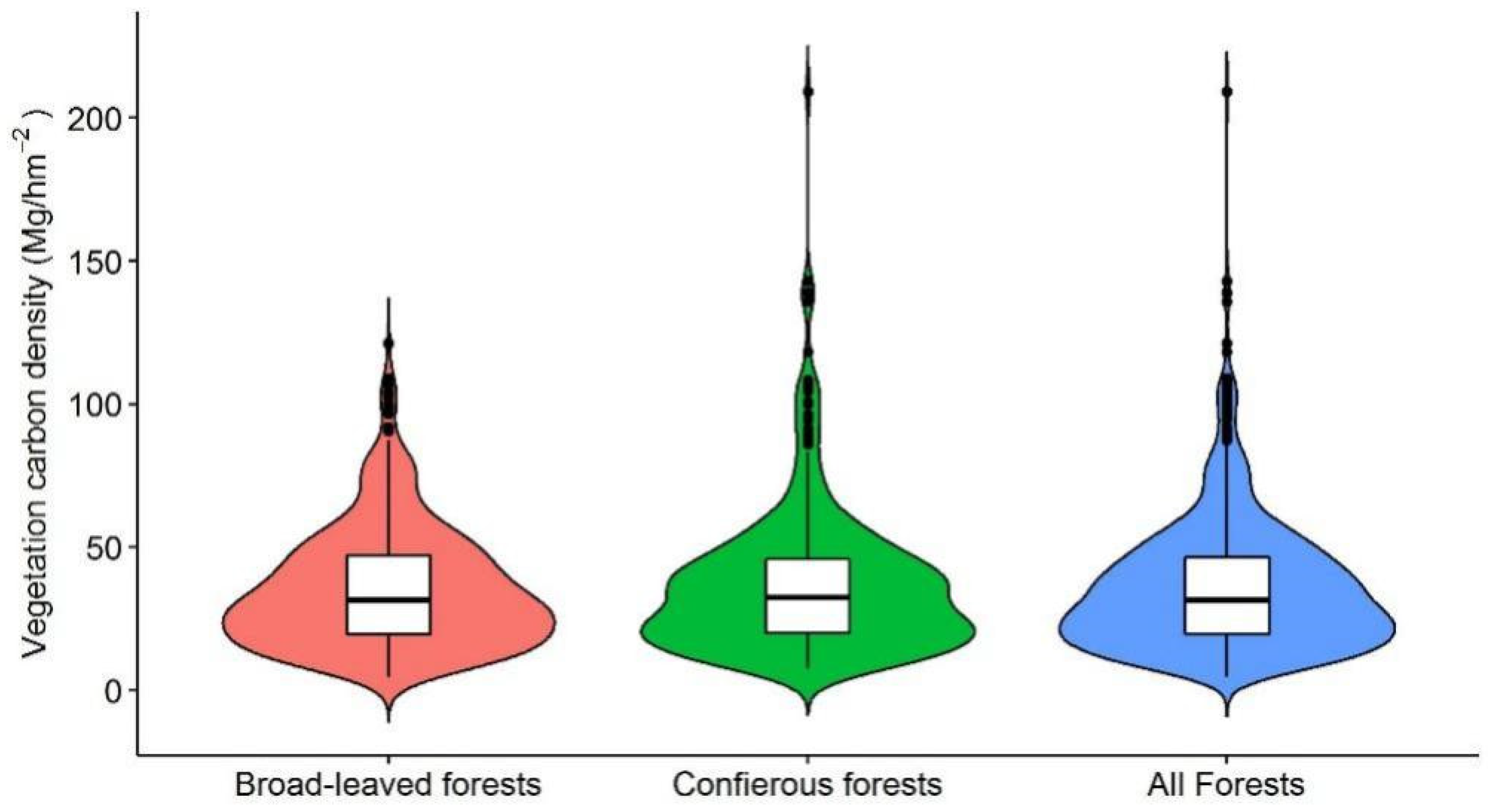
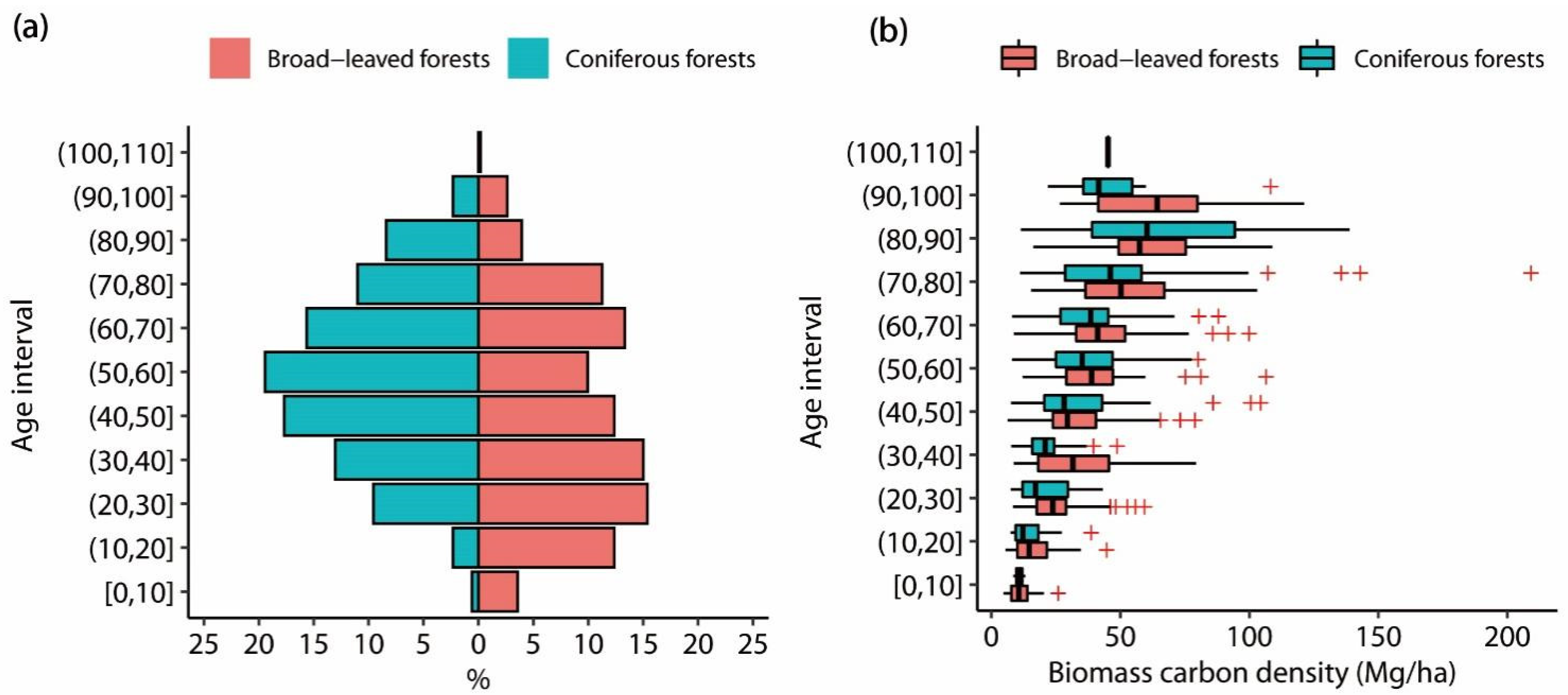
3.1. Biomass Carbon Density
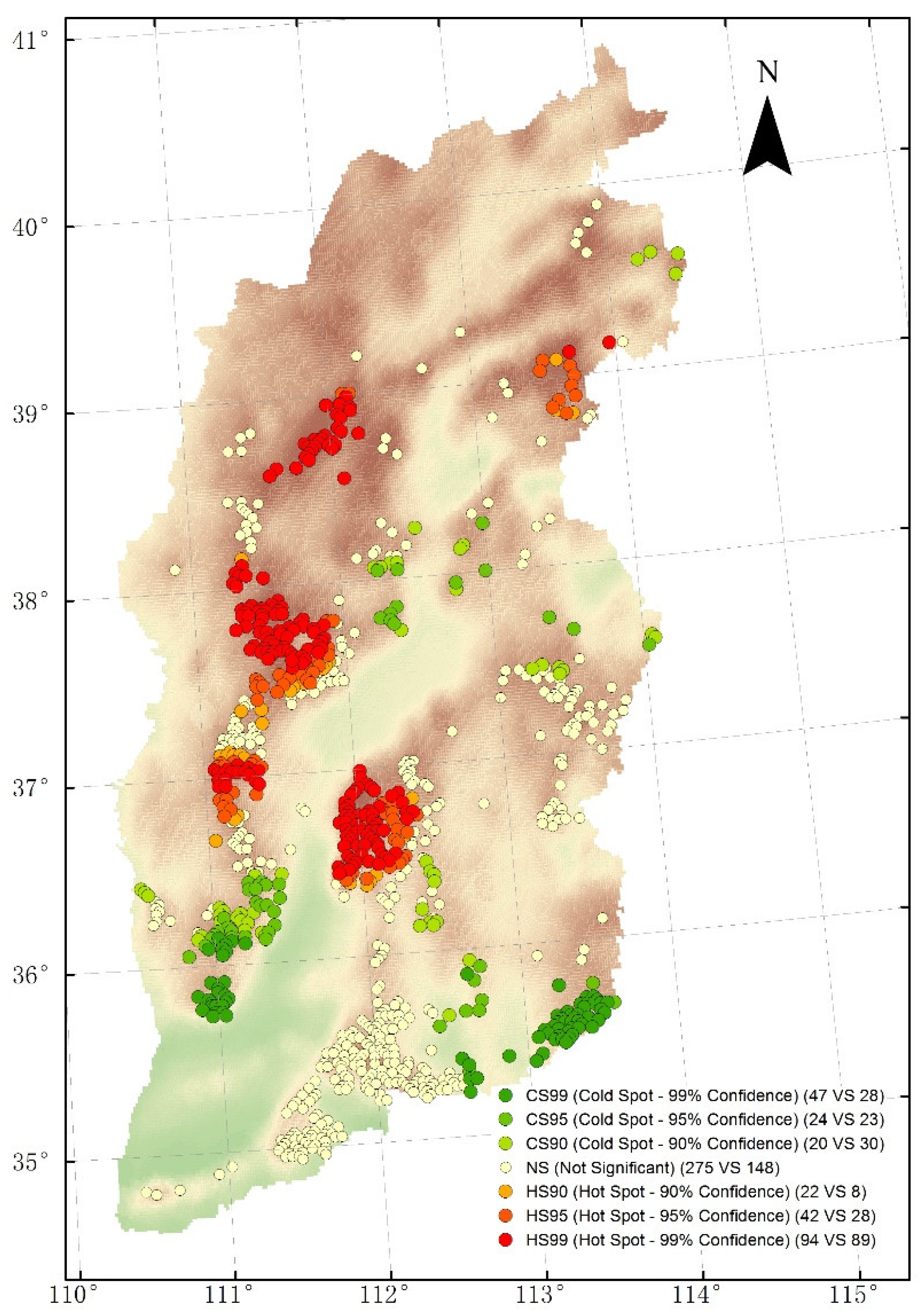
3.2. Spatial Hot Spots and Cold Spots Analyses for Biomass Carbon Densities
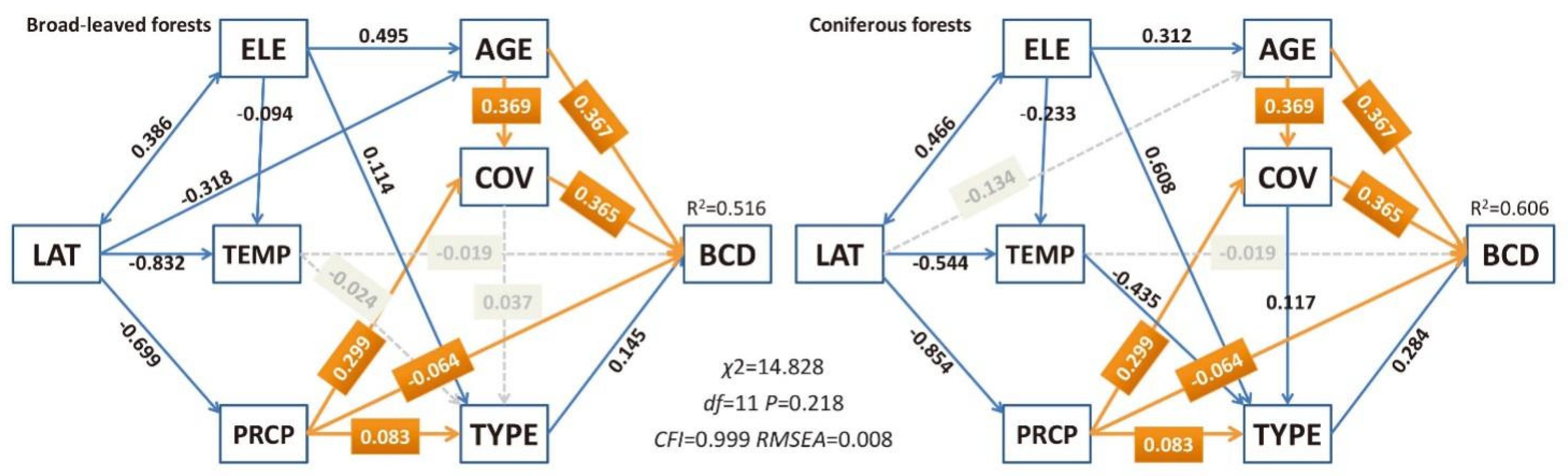
3.3. Influences of Various Factors on Biomass Carbon Density of Natural Mountain Forests
4. Discussion
4.1. Spatial Patterns of Biomass Carbon Density
4.2. The Relationships between Stand Factors and Biomass Carbon Densities
4.3. The Effects of Geographical Factors on Biomass Carbon Density
4.4. The Effects of Climatic Factors on Biomass Carbon Density
4.5. Integrative Framework of Biomass Carbon Density for the Mountainous Forests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. IPCC Climate Change 2014: Synthesis Report; Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014.
- FAO. FAO Global Forest Resources Assessment. 2020. Available online: https://www.fao.org/forest-resources-assessment/en/ (accessed on 15 July 2022).
- Chave, J.; Condit, R.; Lao, S.; Caspersen, J.P.; Foster, R.B.; Hubbell, S.P. Spatial and Temporal Variation of Biomass in a Tropical Forest: Results from a Large Census Plot in Panama. J. Ecol. 2003, 13, 240–252. [Google Scholar] [CrossRef]
- Slik, J.W.F.; Aiba, S.-I.; Brearley, F.Q.; Cannon, C.H.; Forshed, O.; Kitayama, K.; Nagamasu, H.; Nilus, R.; Payne, J.; Paoli, G.; et al. Environmental Correlates of Tree Biomass, Basal Area, Wood Specific Gravity and Stem Density Gradients in Borneo’s Tropical Forests. Glob. Ecol. Biogeogr. 2010, 19, 50–60. [Google Scholar] [CrossRef]
- Xu, M.; Ji, H.; Zhuang, S. Carbon Stock of Moso Bamboo (Phyllostachys Pubescens) Forests along a Latitude Gradient in the Subtropical Region of China. PLoS ONE 2018, 13, e0193024. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Shi, Y.; Fang, H.; Zhou, G.; Xu, X.; Zhou, Y.; Tao, J.; Ji, B.; Xu, J.; Li, C. Vegetation Carbon Stocks Driven by Canopy Density and Forest Age in Subtropical Forest Ecosystems. Sci. Total. Environ. 2018, 631–632, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Li, M.; Chen, H.; Lewis, B.J.; Yu, D.; Zhou, L.; Zhou, W.; Fang, X.; Zhao, W.; Dai, L. Variation in Carbon Storage and Its Distribution by Stand Age and Forest Type in Boreal and Temperate Forests in Northeastern China. PLoS ONE 2013, 8, e72201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.F.; Butt, N.; International Union of Forestry Research Organizations (Eds.) Forests in Sustainable Mountain Development: A State of Knowledge Report for 2000; IUFRO Research Series; CABI Pub.: Wallingford, UK; International Union of Forestry Research Organizations: New York, NY, USA, 2000; ISBN 978-085-199-446-8. [Google Scholar]
- Li, S.; Hu, S.; Tang, X. Dynamic Change of Forest Carbon Stocks; Science Press: Beijing, China, 2013; ISBN 978-7-03-037127-0. [Google Scholar]
- Investigation and Planning Institute of the Ministry of forestry of China. The Mountain Forests in China; China Forestry Publishing House: Beijing, China, 1981.
- Chang, R.; Wang, G.; Fei, R.; Yang, Y.; Luo, J.; Fan, J. Altitudinal Change in Distribution of Soil Carbon and Nitrogen in Tibetan Montane Forests. Soil Sci. Soc. Am. J. 2015, 79, 1455–1469. [Google Scholar] [CrossRef]
- Zald, H.S.J.; Spies, T.A.; Seidl, R.; Pabst, R.J.; Olsen, K.A.; Steel, E.A. Complex Mountain Terrain and Disturbance History Drive Variation in Forest Aboveground Live Carbon Density in the Western Oregon Cascades, USA. For. Ecol. Manag. 2016, 366, 193–207. [Google Scholar] [CrossRef] [Green Version]
- Fu, W.; Fu, Z.; Ge, H.; Ji, B.; Jiang, P.; Li, Y.; Wu, J.; Zhao, K. Spatial Variation of Biomass Carbon Density in a Subtropical Region of Southeastern China. Forests 2015, 6, 1966–1981. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Chao, L.; Wu, C.; Hong, W.; Hong, T.; Hu, X. Spatial Analysis of Carbon Storage Density of Mid-Subtropical Forests Using Geostatistics: A Case Study in Jiangle County, Southeast China. Acta Geochim. 2017, 37, 90–101. [Google Scholar] [CrossRef]
- Ren, H.; Chen, H.; Li, L.; Li, P.; Hou, C.; Wan, H.; Zhang, Q.; Zhang, P. Spatial and Temporal Patterns of Carbon Storage from 1992 to 2002 in Forest Ecosystems in Guangdong, Southern China. Plant Soil 2013, 363, 123–138. [Google Scholar] [CrossRef]
- Dai, W.; Fu, W.; Jiang, P.; Zhao, K.; Li, Y.; Tao, J. Spatial Pattern of Carbon Stocks in Forest Ecosystems of a Typical Subtropical Region of Southeastern China. For. Ecol. Manag. 2018, 409, 288–297. [Google Scholar] [CrossRef]
- Gairola, S.; Sharma, C.M.; Ghildiyal, S.K.; Suyal, S. Live Tree Biomass and Carbon Variation along an Altitudinal Gradient in Moist Temperate Valley Slopes of the Garhwal Himalaya (India). Curr. Sci. India 2011, 100, 1862–1870. [Google Scholar]
- Liu, N.; Nan, H. Carbon Stocks of Three Secondary Coniferous Forests along an Altitudinal Gradient on Loess Plateau in Inland China. PLoS ONE 2018, 13, e0196927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisk, M.C.; Schmidt, S.K.; Seastedt, T.R. Topographic Patterns of Above- and Belowground Production and Nitrogen Cycling in Alpine Tundra. Ecology 1998, 79, 2253–2266. [Google Scholar] [CrossRef]
- Sanaei, A.; Chahouki, M.A.Z.; Ali, A.; Jafari, M.; Azarnivand, H. Abiotic and Biotic Drivers of Aboveground Biomass in Semi-Steppe Rangelands. Sci. Total Environ. 2018, 615, 895–905. [Google Scholar] [CrossRef]
- Wen, D.; He, N. Forest Carbon Storage along the North-South Transect of Eastern China: Spatial Patterns, Allocation, and Influencing Factors. Ecol. Indic. 2016, 61, 960–967. [Google Scholar] [CrossRef]
- Fan, Y.Q.; Zhou, G.M.; Shi, Y.J.; Dong, D.J.; Zhou, Y.F. Relationship of Slope Aspect and Position on Biomass and Carbon Storage in a Phyllostachys Edulis Stand. J. Zhejiang A F Univ. 2012, 29, 321–327. [Google Scholar]
- Durán, S.M.; Sánchez-Azofeifa, G.A.; Rios, R.S.; Gianoli, E. The Relative Importance of Climate, Stand Variables and Liana Abundance for Carbon Storage in Tropical Forests: Climate, Stand Variables and Lianas Control Forest Carbon. Glob. Ecol. Biogeogr. 2015, 24, 939–949. [Google Scholar] [CrossRef]
- Stegen, J.C.; Swenson, N.G.; Enquist, B.J.; White, E.P.; Phillips, O.L.; Jørgensen, P.M.; Weiser, M.D.; Monteagudo Mendoza, A.; Núñez Vargas, P. Variation in Above-Ground Forest Biomass across Broad Climatic Gradients. Glob. Ecol. Biogeogr. 2011, 20, 744–754. [Google Scholar] [CrossRef]
- Xu, B.; Guo, Z.D.; Piao, S.L.; Fang, J.Y. Biomass Carbon Stocks in China’s Forests between 2000 and 2050: A Prediction Based on Forest Biomass-Age Relationships. Sci. China Life Sci. 2010, 53, 776–783. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, G.; Wang, Q.; Zhang, Y. How Temperature, Precipitation and Stand Age Control the Biomass Carbon Density of Global Mature Forests. Glob. Ecol. Biogeogr. 2014, 23, 323–333. [Google Scholar] [CrossRef]
- Shipley, B. Cause and Correlation in Biology: A User’s Guide to Path Analysis, Structural Equations and Causal Inference with R, 2nd ed.; Cambridge University Press: Cambridge, UK, 2016; ISBN 978-110-744-259-7. [Google Scholar]
- Delgado-Baquerizo, M.; Grinyer, J.; Reich, P.B.; Singh, B.K. Relative Importance of Soil Properties and Microbial Community for Soil Functionality: Insights from a Microbial Swap Experiment. Funct. Ecol. 2016, 30, 1862–1873. [Google Scholar] [CrossRef]
- Grace, J.B.; Anderson, T.M.; Olff, H.; Scheiner, S.M. On the Specification of Structural Equation Models for Ecological Systems. Ecol. Monogr. 2010, 80, 67–87. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Yan, E.-R.; Chen, H.Y.H.; Chang, S.X.; Zhao, Y.T.; Yang, X.D.; Xu, M.S. Stand Structural Diversity Rather than Species Diversity Enhances Aboveground Carbon Storage in Secondary Subtropical Forests in Eastern China. Biogeosciences 2016, 13, 4627–4635. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, M.; Wardle, D.A. Structural Equation Modelling Reveals Plant-Community Drivers of Carbon Storage in Boreal Forest Ecosystems. Biol. Lett. 2010, 6, 116–119. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, Q.; Wang, M. Similar Carbon Density of Natural and Planted Forests in the Lüliang Mountains, China. Ann. For. Sci. 2018, 75, 87. [Google Scholar] [CrossRef] [Green Version]
- Wang, N. Study on Distribution Patterns of Carbon Density and Carbon Stock in the Forest Ecosystem of Shanxi; Beijing Forestry University: Beijing, China, 2014. (In Chinese) [Google Scholar]
- Ma, Z. Vegetation of Shanxi Province; China Science and Technology Press: Beijing, China, 2001; ISBN 978-750-463-108-4. (In Chinese) [Google Scholar]
- Fan, X.; Wang, M. Change Trends of Air Temperature and Precipitation over Shanxi Province, China. Theor. Appl. Climatol. 2011, 103, 519–531. [Google Scholar] [CrossRef]
- Xiao, X. National Forest Inventory of China; China Forestry Publishing House: Beijing, China, 2005. (In Chinese) [Google Scholar]
- Sun, L.; Wang, M.; Fan, X. Spatial Pattern and Driving Factors of Biomass Carbon Density for Natural and Planted Coniferous Forests in Mountainous Terrain, Eastern Loess Plateau of China. For. Ecosyst. 2020, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Lamsal, S.; Cobb, R.C.; Hall Cushman, J.; Meng, Q.; Rizzo, D.M.; Meentemeyer, R.K. Spatial Estimation of the Density and Carbon Content of Host Populations for Phytophthora Ramorum in California and Oregon. For. Ecol. Manag. 2011, 262, 989–998. [Google Scholar] [CrossRef]
- Plouffe, C.C.F.; Robertson, C.; Chandrapala, L. Comparing Interpolation Techniques for Monthly Rainfall Mapping Using Multiple Evaluation Criteria and Auxiliary Data Sources: A Case Study of Sri Lanka. Environ. Model. Softw. 2015, 67, 57–71. [Google Scholar] [CrossRef]
- Eldeiry, A.A.; Garcia, L.A. Comparison of Ordinary Kriging, Regression Kriging, and Cokriging Techniques to Estimate Soil Salinity Using LANDSAT Images. J. Irrig. Drain. Eng. 2010, 136, 355–364. [Google Scholar] [CrossRef]
- Manepalli, U.R.R.; Bham, G.H.; Kandada, S. Evaluation of Hot-Spots Identification Using Kernel Density Estimation and Getis-Ord on I-630. In Proceedings of the 3rd International Conference on Road Safety and Simulation, Indianapolis, IN, USA, 14–16 September 2011; Volume 9, pp. 14–16. [Google Scholar]
- Rosseel, Y. Lavaan: An R Package for Structural Equation Modeling. J. Stat. Soft. 2012, 48, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Shanxi Geological Exploration Bureau. Mineral Exploration Results; Shanxi Geological Exploration Bureau: Taiyuan, China, 2018; Available online: http://www.sxdkj.com/ztzl/dzcj/2018-12-21/10221.html (accessed on 15 July 2022).
- Shi, J.R. Overview of geological and mineral resources in Shanxi Province. Land Resour. North China 2017, 4, 76–78. (In Chinese) [Google Scholar]
- Shanxi Fenwei Energy Development Consulting Co., Ltd. Shanxi Coal Resources Map. 2007. Available online: www.sxcoal.com (accessed on 15 July 2022).
- Zhang, F.; Wu, Y. Illuistration Handbook of Plants in Zhongtiao Mountains; China Forestry Publishing House: Beijing, China, 2015; ISBN 978-750-387-936-4. [Google Scholar]
- Chen, W.M.; Zhang, C.X.; Lu, J.R.; Dang, Z.F.; Li, S.P.; Cai, W.B.; Ning, Y.; Cao, Y.J. Polygenetic Mineralization of the Early Proterozoic Tongkuangyu Metaporphyry Copper Deposit in the Zhongtiao Mountains of China. Geotecton. Metallog. 1996, Z1, 110–111. [Google Scholar]
- Pregitzer, K.S.; Euskirchen, E.S. Carbon Cycling and Storage in World Forests: Biome Patterns Related to Forest Age. Glob. Change Biol. 2004, 10, 2052–2077. [Google Scholar] [CrossRef]
- Cheng, X.; Han, H.; Kang, F.; Song, Y.; Liu, K. Variation in Biomass and Carbon Storage by Stand Age in Pine (Pinus tabulaeformis) Planted Ecosystem in Mt. Taiyue, Shanxi, China. J. Plant Interact. 2014, 9, 521–528. [Google Scholar] [CrossRef]
- Fonseca, W.; Rey Benayas, J.M.; Alice, F.E. Carbon Accumulation in the Biomass and Soil of Different Aged Secondary Forests in the Humid Tropics of Costa Rica. For. Ecol. Manag. 2011, 262, 1400–1408. [Google Scholar] [CrossRef]
- Gower, S.T. Patterns and Mechanisms of the Forest Carbon Cycle. Ann. Rev. Env. Resour. 2003, 28, 169–204. [Google Scholar] [CrossRef]
- Mcdowell, G.R.; Harireche, O. Discrete Element Modelling of Yielding and Normal Compression of Sand. Géotechnique 2002, 52, 299–304. [Google Scholar] [CrossRef]
- Tang, J.; Luyssaert, S.; Richardson, A.D.; Kutsch, W.; Janssens, I.A. Steeper Declines in Forest Photosynthesis than Respiration Explain Age-Driven Decreases in Forest Growth. Proc. Natl. Acad. Sci. USA 2014, 111, 8856–8860. [Google Scholar] [CrossRef] [Green Version]
- Thorpe, H.C.; Daniels, L.D. Long-Term Trends in Tree Mortality Rates in the Alberta Foothills Are Driven by Stand Development. Can. J. For. Res. 2012, 42, 1687–1696. [Google Scholar] [CrossRef]
- Goulden, M.L.; Mcmillan, A.M.S.; Winston, G.C.; Rocha, A.V.; Manies, K.L.; Harden, J.W.; Bond-Lamberty, B.P. Patterns of NPP, GPP, Respiration, and NEP during Boreal Forest Succession: Carbon Dynamics during Boreal Succession. Glob. Change Biol. 2011, 17, 855–871. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Fan, X.; Wang, M. Recent Warming Amplification over High Elevation Regions across the Globe. Clim. Dynam. 2014, 43, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Fehse, J.; Hofstede, R.; Aguirre, N.; Paladines, C.; Kooijman, A.; Sevink, J. High Altitude Tropical Secondary Forests: A Competitive Carbon Sink? For. Ecol. Manag. 2002, 163, 9–25. [Google Scholar] [CrossRef]
- Seedre, M.; Kopáček, J.; Janda, P.; Bače, R.; Svoboda, M. Carbon Pools in a Montane Old-Growth Norway Spruce Ecosystem in Bohemian Forest: Effects of Stand Age and Elevation. For. Ecol. Manag. 2015, 346, 106–113. [Google Scholar] [CrossRef]
- Zhu, B.A.; Wang, X.P.; Fang, J.Y.; Piao, S.L.; Shen, H.H.; Zhao, S.Q.; Peng, C.H. Altitudinal Changes in Carbon Storage of Temperate Forests on Mt Changbai, Northeast China. J. Plant Res. 2010, 123, 439–452. [Google Scholar] [CrossRef] [PubMed]

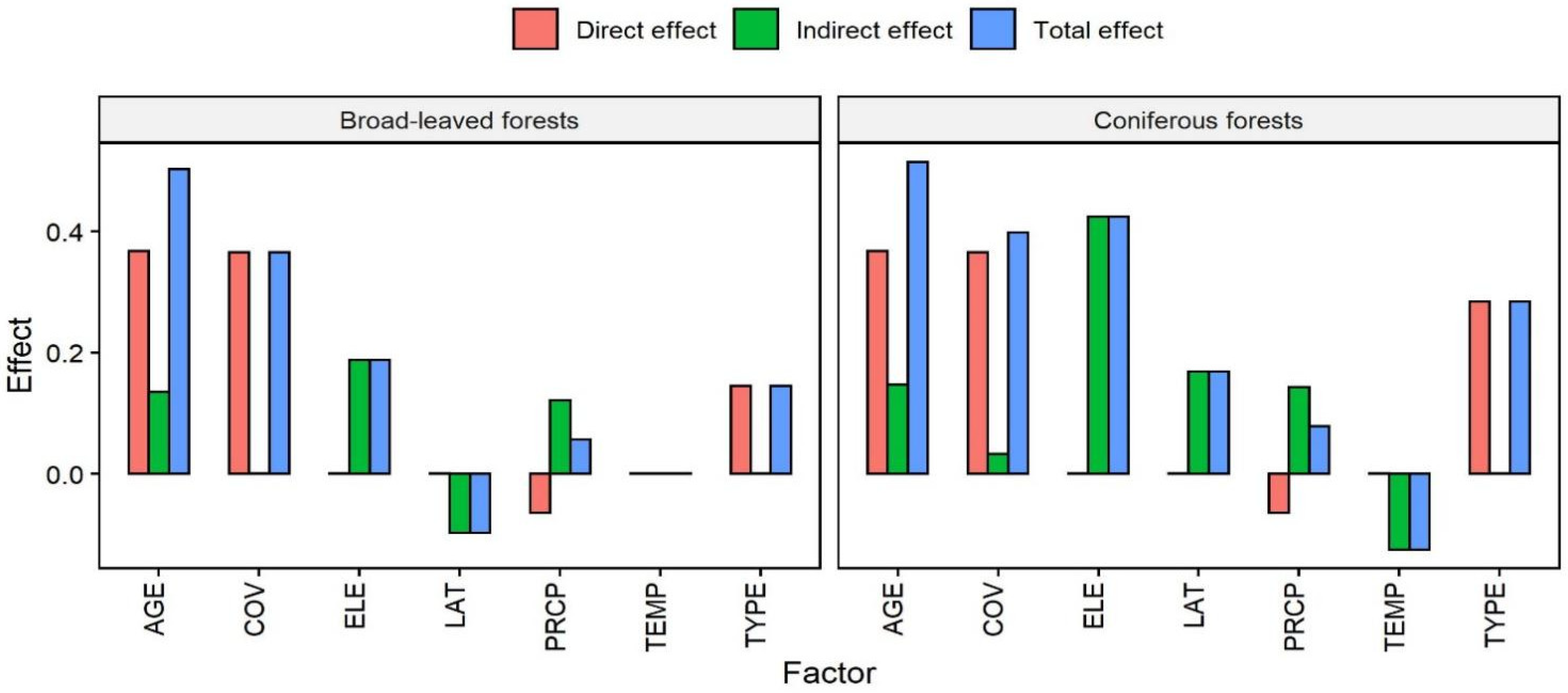

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Type | ELE (km) | LAT (°) | TEMP (°C) | PRCP (mm) | AGE (yr) | COV (%) | |
|---|---|---|---|---|---|---|---|
| Broad-leaved forests | Quercus liaotungensis | 1522.50 | 36.84 | 9.72 | 500.10 | 57 | 63 |
| Broad-leaved mixed | 1358.79 | 36.30 | 10.80 | 528.91 | 43 | 61 | |
| Betula platyphylla | 1924.57 | 38.45 | 6.87 | 474.03 | 45 | 59 | |
| Quercus variabilis | 1048.27 | 35.34 | 12.43 | 584.30 | 43 | 70 | |
| Quercus acutissima | 976.48 | 35.23 | 12.88 | 579.25 | 44 | 75 | |
| Populus davidiana | 1545.72 | 37.84 | 8.780 | 463.77 | 29 | 54 | |
| Others | 1247.84 | 36.2 | 10.55 | 543.05 | 30 | 58 | |
| Mean | 1421.01 | 36.6 | 10.17 | 518.57 | 46 | 62 | |
| Coniferous forests | Pinus tabuliformis | 1436.13 | 36.87 | 9.62 | 502.26 | 57 | 56 |
| Coniferous mixed | 1506.84 | 36.94 | 9.73 | 495.41 | 50 | 58 | |
| Platycladus orientalis | 970.90 | 36.07 | 11.20 | 520.21 | 50 | 50 | |
| Pinus bungeana | 1223.65 | 36.34 | 10.85 | 496.86 | 50 | 68 | |
| Picea wilsonii | 2311.46 | 38.75 | 5.67 | 469.67 | 70 | 69 | |
| Larix principis-rupprechtii | 2230.15 | 38.77 | 5.94 | 469.18 | 56 | 52 | |
| Mean | 1465.32 | 36.93 | 9.57 | 499.18 | 55 | 57 | |
| Mean | 1438.42 | 1438.42 | 9.94 | 509.77 | 50 | 60 | |
| Forest Type | Arbor | Undergrowth | All | No. of Plots | |
|---|---|---|---|---|---|
| Broad-leaved forests | Quercus liaotungensis | 39.73 | 5.97 | 45.7 | 204 |
| Broad-leaved mixed | 25 | 5.81 | 30.8 | 151 | |
| Betula platyphylla | 32.39 | 5.95 | 38.33 | 37 | |
| Quercus variabilis | 29.68 | 5.69 | 35.37 | 26 | |
| Quercus acutissima | 27.96 | 6.33 | 34.29 | 21 | |
| Populus davidiana | 18.66 | 6.35 | 25.01 | 18 | |
| Others | 15.24 | 6.32 | 21.56 | 76 | |
| Mean | 29.89 | 5.99 | 35.87 | 533 | |
| Coniferous forests | Pinus tabuliformis | 27.79 | 6.41 | 34.21 | 176 |
| Coniferous mixed | 32.64 | 7.91 | 40.55 | 93 | |
| Platycladus orientalis | 10.16 | 6.75 | 16.9 | 30 | |
| Pinus bungeana | 18.08 | 6.46 | 24.54 | 20 | |
| Picea wilsonii | 95.93 | 9.84 | 105.77 | 13 | |
| Larix principis-rupprechtii | 47.47 | 7.68 | 55.15 | 13 | |
| Mean | 30.31 | 7.02 | 37.34 | 345 | |
| Mean | 30.06 | 6.39 | 36.45 | 878 | |
| Free Parameter Whose between-Groups Was Constrained to Be Equal | MLχ2 | ΔMLχ2 | p-Value |
|---|---|---|---|
| none | 6.629 | ||
| path from ELE to AGE | 16.179 | 3.974 | 0.002 |
| path from AGE to BCD | 7.325 | 0.696 | 0.404 |
| path from PRCP to COV | 8.548 | 1.922 | 0.166 |
| path from LAT to PRCP | 15.436 | 9.041 | 0.003 |
| path from COV to BCD | 8.089 | 1.461 | 0.227 |
| path from TEMP to BCD | 17.457 | 0.026 | 0.001 |
| path from PRCP to BCD | 6.629 | 0.002 | 0.986 |
| path from LAT to AGE | 16.179 | 3.063 | 0.002 |
| path from AGE to COV | 6.837 | 0.208 | 0.648 |
| path from PRCP to TYPE | 14.913 | 2.620 | 0.004 |
| path from ELE to TEMP | 17.457 | 7.559 | 0.001 |
| Path from COV to TYPE | 17.457 | 3.046 | 0.001 |
| path from TYPE to BCD | 16.179 | 3.281 | 0.002 |
| path from ELE to TYPE | 38.211 | 31.582 | <0.001 |
| path from TEMP to TYPE | 20.396 | 13.767 | <0.001 |
| path from LAT to TEMP | 35.044 | 28.415 | <0.001 |
| path from LAT to ELE | 24.932 | 18.303 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.; Wang, Q.; Fan, X. An Integrative Approach to Study How Driving Factors Control Biomass Carbon Density for Natural Mountain Forests, in China’s Loess Plateau. Forests 2022, 13, 1114. https://doi.org/10.3390/f13071114
Sun L, Wang Q, Fan X. An Integrative Approach to Study How Driving Factors Control Biomass Carbon Density for Natural Mountain Forests, in China’s Loess Plateau. Forests. 2022; 13(7):1114. https://doi.org/10.3390/f13071114
Chicago/Turabian StyleSun, Lina, Qixiang Wang, and Xiaohui Fan. 2022. "An Integrative Approach to Study How Driving Factors Control Biomass Carbon Density for Natural Mountain Forests, in China’s Loess Plateau" Forests 13, no. 7: 1114. https://doi.org/10.3390/f13071114
APA StyleSun, L., Wang, Q., & Fan, X. (2022). An Integrative Approach to Study How Driving Factors Control Biomass Carbon Density for Natural Mountain Forests, in China’s Loess Plateau. Forests, 13(7), 1114. https://doi.org/10.3390/f13071114