
Fire Damage to the Soil Bacterial Structure and Function Depends on Burn Severity: Experimental Burnings at a Lysimetric Facility (MedForECOtron)

 ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Soil Sampling and Analysis
2.2.1. Physico-Chemical Analysis
2.2.2. Biological Analysis
2.3. Statistical Analysis
3. Results
3.1. Validating Burn Severity Levels
3.2. Changes in the Physico-Chemical Soil Properties
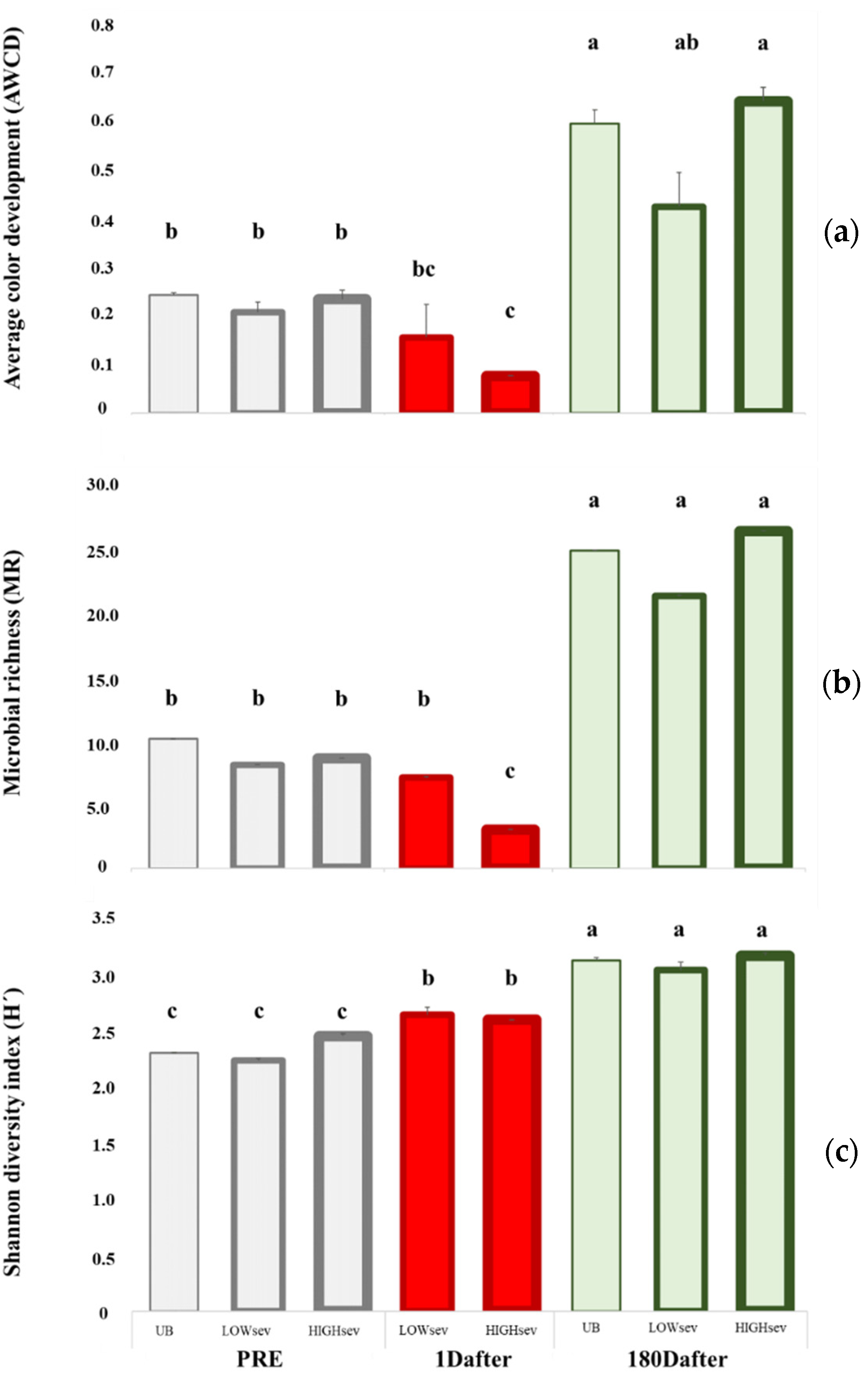
3.3. Changes in the Microbiological Soil Properties
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CLPP | community-level physiological profiles |
| AWCD | average well-color development |
| MR | microbial richness |
| H’ | Shannon diversity index |
| CA | carboxylic acids; AMN: amines/amides |
| AAC | amino acids |
| CH | carbohydrates |
| PHE | phenolic compounds |
| POL | polymers |
References
- European Commission Our Life Insurance. Our Natural Capital: An EU Biodiversity Strategy to 2020; European Commission: Brussels, Belgium, 2011; Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:52011DC0244&from=EN (accessed on 21 June 2022).
- Orgiazzi, A.; Panagos, P.; Yigini, Y.; Dunbar, M.B.; Gardi, C.; Montanarella, L.; Ballabio, C. A Knowledge-Based Approach to Estimating the Magnitude and Spatial Patterns of Potential Threats to Soil Biodiversity. Sci. Total Environ. 2016, 545–546, 11–20. [Google Scholar] [CrossRef]
- Doran, J.W.; Parkin, T.B. Defining and Assessing Soil Quality. In Defining Soil Quality for a Sustainable Environment; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1994; pp. 1–21. ISBN 978-0-89118-930-5. [Google Scholar]
- Fließbach, A.; Oberholzer, H.-R.; Gunst, L.; Mäder, P. Soil Organic Matter and Biological Soil Quality Indicators after 21 Years of Organic and Conventional Farming. Agric. Ecosyst. Environ. 2007, 118, 273–284. [Google Scholar] [CrossRef]
- Allison, S.D.; Martiny, J.B.H. Resistance, Resilience, and Redundancy in Microbial Communities. Proc. Natl. Acad. Sci. USA 2008, 105, 11512–11519. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, S.; Walker, B.; Anderies, J.M.; Abel, N. From Metaphor to Measurement: Resilience of What to What? Ecosystems 2001, 4, 765–781. [Google Scholar] [CrossRef]
- Gunderson, L.H. Ecological Resilience—In Theory and Application. Annu. Rev. Ecol. Syst. 2000, 31, 425–439. [Google Scholar] [CrossRef] [Green Version]
- Rüdisser, J.; Tasser, E.; Peham, T.; Meyer, E.; Tappeiner, U. The Dark Side of Biodiversity: Spatial Application of the Biological Soil Quality Indicator (BSQ). Ecol. Indic. 2015, 53, 240–246. [Google Scholar] [CrossRef]
- Kelly, L.T.; Giljohann, K.M.; Duane, A.; Aquilué, N.; Archibald, S.; Batllori, E.; Bennett, A.F.; Buckland, S.T.; Canelles, Q.; Clarke, M.F.; et al. Fire and Biodiversity in the Anthropocene. Science 2020, 370, eabb0355. [Google Scholar] [CrossRef]
- Certini, G.; Moya, D.; Lucas-Borja, M.E.; Mastrolonardo, G. The Impact of Fire on Soil-Dwelling Biota: A Review. For. Ecol. Manag. 2021, 488, 118989. [Google Scholar] [CrossRef]
- Acea, M.J.; Carballas, T. Microbial Fluctuations after Soil Heating and Organic Amendment. Bioresour. Technol. 1999, 67, 65–71. [Google Scholar] [CrossRef]
- Mataix-Solera, J.; Navarro-Pedreño, J.; Guerrero, C.; Gómez Lucas, I.; Marco, B.; Mataix, J. Effects of an Experimental Fire on Soil Microbial Populations in a Mediterranean Environment. In Man and Soil at the Third Millenium; Geoforma Ediciones: Logroño, Spain, 2002; pp. 1607–1614. ISBN 978-84-87779-47-3. [Google Scholar]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Forest Soil Bacteria: Diversity, Involvement in Ecosystem Processes, and Response to Global Change. Microbiol. Mol. Biol. Rev. 2017, 81, e00063-16. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Valera, E.; Verdú, M.; Navarro-Cano, J.A.; Goberna, M. Soil Microbiome Drives the Recovery of Ecosystem Functions after Fire. Soil Biol. Biochem. 2020, 149, 107948. [Google Scholar] [CrossRef]
- Moya, D.; Madrigal, J.; Fonturbel, T.; Marino, E.; Hernando, C.; Guijarro, M.; Fernandez, C.; Jimenez, E.; Lucas-Borja, M.; Vega, J.; et al. Fire Severity Assessments in Both the Laboratory and the Field. In Fire Effects on Soil Properties; Pereira, P., Mataix-Solera, J., Ubeda, X., Rein, G., Cerda, A., Eds.; CSIRO Publishing: Clayton, Australia, 2019; pp. 241–266. [Google Scholar]
- Alcañiz, M.; Outeiro, L.; Francos, M.; Úbeda, X. Effects of Prescribed Fires on Soil Properties: A Review. Sci. Total Environ. 2018, 613–614, 944–957. [Google Scholar] [CrossRef]
- Cawson, J.G.; Sheridan, G.J.; Smith, H.G.; Lane, P.N.J. Effects of Fire Severity and Burn Patchiness on Hillslope-Scale Surface Runoff, Erosion and Hydrologic Connectivity in a Prescribed Burn. For. Ecol. Manag. 2013, 310, 219–233. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Delgado-Baquerizo, M.; Muñoz-Rojas, M.; Plaza-Álvarez, P.A.; Gómez-Sanchez, M.E.; González-Romero, J.; Peña-Molina, E.; Moya, D.; de las Heras, J. Changes in Ecosystem Properties after Post-Fire Management Strategies in Wildfire-Affected Mediterranean Forests. J. Appl. Ecol. 2021, 58, 836–846. [Google Scholar] [CrossRef]
- Pérez-Valera, E.; Verdú, M.; Navarro-Cano, J.A.; Goberna, M. Resilience to Fire of Phylogenetic Diversity across Biological Domains. Mol. Ecol. 2018, 27, 2896–2908. [Google Scholar] [CrossRef]
- Muñoz-Rojas, M.; Barcenas-Moreno, G. Microbiology. In Fire Effects on Soil Properties; Pereira, P., Ubeda, X., Mataix-Solera, J., Rein, G., Cerda, A., Eds.; CSIRO Publishing: Clayton, Australia, 2019; pp. 157–174. [Google Scholar]
- Moya, D.; Fonturbel, M.T.; Lucas-Borja, M.E.; Peña, E.; Alfaro-Sanchez, R.; Plaza-Álvarez, P.A.; González-Romero, J.; de Las Heras, J. Burning Season and Vegetation Coverage Influenced the Community-Level Physiological Profile of Mediterranean Mixed-Mesogean Pine Forest Soils. J. Environ. Manag. 2021, 277, 111405. [Google Scholar] [CrossRef]
- Wang, Q.; Zhong, M.; Wang, S. A Meta-Analysis on the Response of Microbial Biomass, Dissolved Organic Matter, Respiration, and N Mineralization in Mineral Soil to Fire in Forest Ecosystems. For. Ecol. Manag. 2012, 271, 91–97. [Google Scholar] [CrossRef]
- Fontúrbel, M.T.; Barreiro, A.; Vega, J.A.; Martín, A.; Jiménez, E.; Carballas, T.; Fernández, C.; Díaz-Raviña, M. Effects of an Experimental Fire and Post-Fire Stabilization Treatments on Soil Microbial Communities. Geoderma 2012, 191, 51–60. [Google Scholar] [CrossRef]
- Fontúrbel, M.T.; Fernández, C.; Vega, J.A. Prescribed Burning versus Mechanical Treatments as Shrubland Management Options in NW Spain: Mid-Term Soil Microbial Response. Appl. Soil Ecol. 2016, 107, 334–346. [Google Scholar] [CrossRef]
- Pérez-Valera, E.; Goberna, M.; Verdú, M. Fire modulates ecosystem functioning through the phylogenetic structure of soil bacterial communities. Soil Biol. Biochem. 2019, 129, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Turan, V.; Schröder, P.; Bilen, S.; Insam, H.; Fernández-Delgado Juárez, M. Co-Inoculation Effect of Rhizobium and Achillea millefolium L. Oil Extracts on Growth of Common Bean (Phaseolus vulgaris L.) and Soil Microbial-Chemical Properties. Sci. Rep. 2019, 9, 15178. [Google Scholar] [CrossRef] [Green Version]
- Lucas-Borja, M.E.; Ortega, R.; Miralles, I.; Plaza-Álvarez, P.A.; González-Romero, J.; Peña-Molina, E.; Moya, D.; Zema, D.A.; Wagenbrenner, J.W.; de las Heras, J. Effects of Wildfire and Logging on Soil Functionality in the Short-Term in Pinus Halepensis M. Forests. Eur. J. For. Res. 2020, 139, 935–945. [Google Scholar] [CrossRef]
- Bastida, F.; Zsolnay, A.; Hernández, T.; García, C. Past, Present and Future of Soil Quality Indices: A Biological Perspective. Geoderma 2008, 147, 159–171. [Google Scholar] [CrossRef]
- Barrios, E. Soil Biota, Ecosystem Services and Land Productivity. Ecol. Econ. 2007, 64, 269–285. [Google Scholar] [CrossRef]
- Doblas-Miranda, E.; Martínez-Vilalta, J.; Lloret, F.; Álvarez, A.; Ávila, A.; Bonet, F.J.; Brotons, L.; Castro, J.; Yuste, J.C.; Díaz, M.; et al. Reassessing Global Change Research Priorities in Mediterranean Terrestrial Ecosystems: How Far Have We Come and Where Do We Go from Here? Glob. Ecol. Biogeogr. 2015, 24, 25–43. [Google Scholar] [CrossRef] [Green Version]
- Baraza-Ruiz, E.; Bota, J.; Romero-Munar, A.; Nogales, B. Aplicación de la técnica BiologTM ECO-plate para el estudio del perfil fisiológico de las comunidades microbianas del suelo agrícola. Ecosistemas 2019, 28, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Sofo, A.; Ricciuti, P. A Standardized Method for Estimating the Functional Diversity of Soil Bacterial Community by Biolog® EcoPlatesTM Assay—The Case Study of a Sustainable Olive Orchard. Appl. Sci. 2019, 9, 4035. [Google Scholar] [CrossRef] [Green Version]
- Lawton, J.H.; Naeem, S.; Woodfin, R.M.; Brown, V.K.; Gange, A.; Godfray, H.J.C.; Heads, P.A.; Lawler, S.; Magda, D.; Thomas, C.D.; et al. The Ecotron: A Controlled Environmental Facility for the Investigation of Population and Ecosystem Processes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1993, 341, 181–194. [Google Scholar] [CrossRef]
- Granjou, C.; Walker, J. Promises That Matter: Reconfiguring Ecology in the Ecotrons. Sci. Technol. Stud. 2016, 29, 49–67. [Google Scholar] [CrossRef]
- Alcañiz, M.; Outeiro, L.; Francos, M.; Farguell, J.; Úbeda, X. Long-term dynamics of soil chemical properties after a prescribed fire in a Mediterranean forest (Montgrí Massif, Catalonia, Spain). Sci. Total Environ. 2016, 572, 1329–1335. [Google Scholar] [CrossRef]
- Ammitzboll, H.; Jordan, G.J.; Baker, S.C.; Freeman, J.; Bissett, A. Contrasting Successional Responses of Soil Bacteria and Fungi to Post-Logging Burn Severity. For. Ecol. Manag. 2022, 508, 120059. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Miralles, I.; Ortega, R.; Plaza-Álvarez, P.A.; Gonzalez-Romero, J.; Sagra, J.; Soriano-Rodríguez, M.; Certini, G.; Moya, D.; Heras, J. Immediate Fire-Induced Changes in Soil Microbial Community Composition in an Outdoor Experimental Controlled System. Sci. Total Environ. 2019, 696, 134033. [Google Scholar] [CrossRef]
- López-Poma, R.; Bautista, S. Plant Regeneration Functional Groups Modulate the Response to Fire of Soil Enzyme Activities in a Mediterranean Shrubland. Soil Biol. Biochem. 2014, 79, 5–13. [Google Scholar] [CrossRef]
- Moya, D.; González-De Vega, S.; Lozano, E.; García-Orenes, F.; Mataix-Solera, J.; Lucas-Borja, M.E.; de las Heras, J. The Burn Severity and Plant Recovery Relationship Affect the Biological and Chemical Soil Properties of Pinus halepensis Mill. Stands in the Short and Mid-Terms after Wildfire. J. Environ. Manag. 2019, 235, 250–256. [Google Scholar] [CrossRef]
- Martinez-Salgado, M.M.; Gutiérrez-Romero, V.; Jannsens, M.; Ortega-Blu, R. Biological soil quality indicators: A review. Curr. Res. Technol. Educ. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 1, 319–328. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014.
- Andrews, P.L.; Bevins, C.D.; Seli, R.C. BehavePlus Fire Modeling System, Version 4.0: User’s Guide; General Technical Report; Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2008. [Google Scholar]
- Jiménez-Morillo, N.T.; Almendros, G.; De la Rosa, J.M.; Jordán, A.; Zavala, L.M.; Granged, A.J.P.; González-Pérez, J.A. Effect of a Wildfire and of Post-Fire Restoration Actions in the Organic Matter Structure in Soil Fractions. Sci. Total Environ. 2020, 728, 138715. [Google Scholar] [CrossRef]
- Lange, M.; Eisenhauer, N.; Sierra, C.A.; Bessler, H.; Engels, C.; Griffiths, R.I.; Mellado-Vázquez, P.G.; Malik, A.A.; Roy, J.; Scheu, S.; et al. Plant Diversity Increases Soil Microbial Activity and Soil Carbon Storage. Nat. Commun. 2015, 6, 6707. [Google Scholar] [CrossRef]
- Badía, D.; López-García, S.; Martí, C.; Ortíz-Perpiñá, O.; Girona-García, A.; Casanova-Gascón, J. Burn Effects on Soil Properties Associated to Heat Transfer under Contrasting Moisture Content. Sci. Total Environ. 2017, 601–602, 1119–1128. [Google Scholar] [CrossRef]
- Gee, W.G.; Or, D. Particle-Size Analysis. In Methods of Soil Analysis. Book Series: 5. Part 4; Dane, J., Topp, G.C., Eds.; Soil Science Society of America; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2002; pp. 255–293. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1996; pp. 961–1010. ISBN 978-0-89118-866-7. [Google Scholar]
- Lax, A.; Roig, A.; Costa, F. A Method for Determining the Cation-Exchange Capacity of Organic Materials. Plant Soil 1986, 94, 349–355. [Google Scholar] [CrossRef]
- Knudsen, D.; Peterson, G.A.; Pratt, P.F. Lithium, Sodium, and Potassium. In Methods of Soil Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1983; pp. 225–246. ISBN 978-0-89118-977-0. [Google Scholar]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen—Total. In Methods of Soil Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1983; pp. 595–624. ISBN 978-0-89118-977-0. [Google Scholar]
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1983; pp. 403–430. ISBN 978-0-89118-977-0. [Google Scholar]
- Rutgers, M.; Wouterse, M.; Drost, S.M.; Breure, A.M.; Mulder, C.; Stone, D.; Creamer, R.E.; Winding, A.; Bloem, J. Monitoring Soil Bacteria with Community-Level Physiological Profiles Using BiologTM ECO-Plates in the Netherlands and Europe. Appl. Soil Ecol. 2016, 97, 23–35. [Google Scholar] [CrossRef]
- Garland, J.L.; Mills, A.L. Classification and Characterization of Heterotrophic Microbial Communities on the Basis of Patterns of Community-Level Sole-Carbon-Source Utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [Green Version]
- Guénon, R.; Gros, R. Frequent-Wildfires with Shortened Time-since-Fire Affect Soil Microbial Functional Stability to Drying and Rewetting Events. Soil Biol. Biochem. 2013, 57, 663–674. [Google Scholar] [CrossRef]
- Zabinski, C.A.; Gannon, J.E. Effects of Recreational Impacts on Soil Microbial Communities. Environ. Manag. 1997, 21, 233–238. [Google Scholar] [CrossRef]
- Zak, J.C.; Willig, M.R.; Moorhead, D.L.; Wildman, H.G. Functional Diversity of Microbial Communities: A Quantitative Approach. Soil Biol. Biochem. 1994, 26, 1101–1108. [Google Scholar] [CrossRef]
- Bending, G.D.; Putland, C.; Rayns, F. Changes in Microbial Community Metabolism and Labile Organic Matter Fractions as Early Indicators of the Impact of Management on Soil Biological Quality. Biol. Fertil. Soils 2000, 31, 78–84. [Google Scholar] [CrossRef]
- RStudio Team RStudio: Integrated Development for R. RStudio. Available online: http://www.rstudio.com (accessed on 2 February 2021).
- IBM Corp IBM SPSS Statistics for Windows, Version 24.0. Available online: www.ibm.com/mysupport/s/question/0d50z00006pfp7zcad/how-do-you-cite-spss-version-24-in-a-publication (accessed on 2 February 2021).
- Vega, J.A.; Fontúrbel, T.; Merino, A.; Fernández, C.; Ferreiro, A.; Jiménez, E. Testing the Ability of Visual Indicators of Soil Burn Severity to Reflect Changes in Soil Chemical and Microbial Properties in Pine Forests and Shrubland. Plant Soil 2013, 369, 73–91. [Google Scholar] [CrossRef]
- Guénon, R.; Vennetier, M.; Dupuy, N.; Ziarelli, F.; Gros, R. Soil Organic Matter Quality and Microbial Catabolic Functions along a Gradient of Wildfire History in a Mediterranean Ecosystem. Appl. Soil Ecol. 2011, 48, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Fontúrbel, M.T.; Vega, J.A.; Bara, S.; Bernardez, I. Influence of Prescribed Burning of Pine Stands in NW Spain on Soil Microorganisms. Eur. J. Soil Biol. 1995, 31, 13–20. [Google Scholar]
- Doblas-Miranda, E.; Alonso, R.; Arnan, X.; Bermejo, V.; Brotons, L.; de las Heras, J.; Estiarte, M.; Hódar, J.A.; Llorens, P.; Lloret, F.; et al. A Review of the Combination among Global Change Factors in Forests, Shrublands and Pastures of the Mediterranean Region: Beyond Drought Effects. Glob. Planet. Chang. 2017, 148, 42–54. [Google Scholar] [CrossRef] [Green Version]
- Certini, G. Effects of Fire on Properties of Forest Soils: A Review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef]
- Brunel, C.; Da Silva, A.-M.F.; Gros, R. Environmental Drivers of Microbial Functioning in Mediterranean Forest Soils. Microb. Ecol. 2020, 80, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Knicker, H. How Does Fire Affect the Nature and Stability of Soil Organic Nitrogen and Carbon? A Review. Biogeochemistry 2007, 85, 91–118. [Google Scholar] [CrossRef]
- Merino, A.; Omil, B.; Fonturbel, M.T.; Vega, J.A.; Balboa, M.A. Reclamation of Intensively Managed Soils in Temperate Regions by Addition of Wood Bottom Ash Containing Charcoal: SOM Composition and Microbial Functional Diversity. Appl. Soil Ecol. 2016, 100, 195–206. [Google Scholar] [CrossRef]
- Hamman, S.T.; Burke, I.C.; Knapp, E.E. Soil Nutrients and Microbial Activity after Early and Late Season Prescribed Burns in a Sierra Nevada Mixed Conifer Forest. For. Ecol. Manag. 2008, 256, 367–374. [Google Scholar] [CrossRef]
- Aponte, C.; Marañón, T.; García, L.V. Microbial C, N and P in Soils of Mediterranean Oak Forests: Influence of Season, Canopy Cover and Soil Depth. Biogeochemistry 2010, 101, 77–92. [Google Scholar] [CrossRef] [Green Version]
- Lucas-Borja, M.E.; Plaza-Álvarez, P.A.; Ortega, R.; Miralles, I.; Gonzalez-Romero, J.; Sagra, J.; Moya, D.; Zema, D.A.; de las Heras, J. Short-term changes in soil functionality after wildfire and straw mulching in a Pinus halepensis M. forest. For. Ecol. Manag. 2020, 457, 117700. [Google Scholar] [CrossRef]
- Lombao, A.; Barreiro, A.; Fontúrbel, M.T.; Martín, A.; Carballas, T.; Díaz-Raviña, M. Key Factors Controlling Microbial Community Responses after a Fire: Importance of Severity and Recurrence. Sci. Total Environ. 2020, 741, 140363. [Google Scholar] [CrossRef]
- D’Ascoli, R.; Rutigliano, F.A.; Pascale, R.A.D.; Gentile, A.; Santo, A.V.D. Functional Diversity of the Microbial Community in Mediterranean Maquis Soils as Affected by Fires. Int. J. Wildland Fire 2005, 14, 355–363. [Google Scholar] [CrossRef]
- Staddon, W.J.; Duchesne, L.C.; Trevors, J.T. Microbial Diversity and Community Structure of Postdisturbance Forest Soils as Determined by Sole-Carbon-Source Utilization Patterns. Microb. Ecol. 1997, 34, 125–130. [Google Scholar] [CrossRef]
- Giai, C.; Boerner, R.E.J. Effects of Ecological Restoration on Microbial Activity, Microbial Functional Diversity, and Soil Organic Matter in Mixed-Oak Forests of Southern Ohio, USA. Appl. Soil Ecol. 2007, 35, 281–290. [Google Scholar] [CrossRef]
- Gomez, E.; Ferreras, L.; Toresani, S. Soil Bacterial Functional Diversity as Influenced by Organic Amendment Application. Bioresour. Technol. 2006, 97, 1484–1489. [Google Scholar] [CrossRef] [PubMed]
- Tedim, F.; Leone, V.; Amraoui, M.; Bouillon, C.; Coughlan, M.R.; Delogu, G.M.; Fernandes, P.M.; Ferreira, C.; McCaffrey, S.; McGee, T.K.; et al. Defining Extreme Wildfire Events: Difficulties, Challenges, and Impacts. Fire 2018, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Chuvieco, E.; Aguado, I.; Salas, J.; García, M.; Yebra, M.; Oliva, P. Satellite Remote Sensing Contributions to Wildland Fire Science and Management. Curr. For. Rep. 2020, 6, 81–96. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Burn Severity | Sampling Date | SOM | Corg | N | pH | EC | CEC | P | Mg2+ | K+ | Na+ | Ca2+ |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| UB | PRE | 4.28 ± 0.60 A | 2.49 ± 0.35 A | 0.17 ± 0.01 A | 8.25 ± 0.15 A | 1.32 ± 0.65 A | 9.75 ± 2.41 A | 6.30 ± 0.46 A | 6.94 ± 1.14 A | 0.62 ± 0.18 A | 1.06 ± 0.24 A | 35.24 ± 2.82 A |
| 1Dafter | 4.17 ± 0.17 A | 2.49 ± 0.35 A | 0.16 ± 0.02 A | 8.11 ± 0.02 A | 1.39 ± 0.59 A | 10.30 ± 2.95 A | 6.03 ± 0.08 A | 6.99 ± 1.15 A | 0.63 ± 0.14 A | 1.08 ± 0.24 A | 35.30 ± 2.90 A | |
| 180Dafter | 4.37 ± 0.10 A | 4.28 ± 0.01 A | 0.21 ± 0.01 A | 8.40 ± 0.02 A | 1.96 ± 0.21 B | 13.71 ± 1.75 A | 8.00 ± 3.00 A | 6.45 ± 0.11 A | 0.80 ± 0.23 A | 0.62 ± 0.08 A | 39.49 ± 1.74 B | |
| LOWsev | PRE | 3.59 ± 0.24 A | 2.08 ± 0.10 A | 0.15 ± 0.02 A | 8.45 ± 0.15 A | 0.74 ± 0.23 A | 10.82 ± 1.53 A | 5.03 ± 0.11 A | 5.42 ± 0.60 A | 0.55 ± 0.08 A | 0.79 ± 0.16 A | 32.09 ± 0.46 A |
| 1Dafter | 4.87 ± 1.01 A | 3.41 ± 1.17 A | 0.19 ± 0.06 A | 8.55 ± 0.21 A | 0.80 ± 0.22 A | 14.32 ± 0.05 A | 9.58 ± 0.38 B | 7.02 ± 0.52 A | 0.71 ± 0.06 A | 0.61 ± 0.05 A | 36.57 ± 2.47 A | |
| 180Dafter | 4.68 ± 0.28 A | 2.65 ± 0.15 A | 0.16 ± 0.01 A | 8.35 ± 0.05 A | 2.97 ± 0.39 C | 13.23 ± 0.18 A | 10.03 ± 0.01 B | 5.89 ± 0.22 A | 0.66 ± 0.03 A | 0.83 ± 0.26 A | 39.13 ± 0.14 B | |
| HIGHsev | PRE | 5.76 ± 0.74 B | 3.34 ± 0.43 A | 0.18 ± 0.03 A | 8.35 ± 0.05 A | 0.59 ± 0.07 A | 7.52 ± 0.68 A | 5.79 ± 1.43 A | 6.50 ± 0.02 A | 0.59 ± 0.02 A | 0.71 ± 0.25 A | 34.48 ± 0.93 A |
| 1Dafter | 5.58 ± 0.97 B | 3.24 ± 0.57 A | 0.18 ± 0.03 A | 8.69 ± 0.21 A | 0.64 ± 0.01 A | 14.29 ± 1.85 A | 11.43 ± 0.43 B | 7.22 ± 0.03 A | 0.90 ± 0.17 A | 0.83 ± 0.14 A | 36.03 ± 1.44 A | |
| 180Dafter | 6.85 ± 0.88 C | 3.90 ± 0.50 A | 0.24 ± 0.01 A | 8.20 ± 0.02 A | 4.52 ± 0.15 D | 15.57 ± 1.37 A | 18.00 ± 3.02 C | 9.23 ± 0.38 A | 0.75 ± 0.08 A | 0.90 ± 0.05 A | 45.54 ± 1.61 C | |
| p-value | 0.04 * | 0.17 | 0.463 | 0.06 | 0.000 * | 0.24 | 0.001 * | 0.10 | 0.72 | 0.21 | 0.001 * | |
| MSE | 1.317 | 0.668 | 0.57 | 0.032 | 0.171 | 8.802 | 7.644 | 1.161 | 2.19 | 0.06 | 7.274 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moya, D.; Fonturbel, T.; Peña, E.; Alfaro-Sanchez, R.; Plaza-Álvarez, P.A.; González-Romero, J.; Lucas-Borja, M.E.; de Las Heras, J. Fire Damage to the Soil Bacterial Structure and Function Depends on Burn Severity: Experimental Burnings at a Lysimetric Facility (MedForECOtron). Forests 2022, 13, 1118. https://doi.org/10.3390/f13071118
Moya D, Fonturbel T, Peña E, Alfaro-Sanchez R, Plaza-Álvarez PA, González-Romero J, Lucas-Borja ME, de Las Heras J. Fire Damage to the Soil Bacterial Structure and Function Depends on Burn Severity: Experimental Burnings at a Lysimetric Facility (MedForECOtron). Forests. 2022; 13(7):1118. https://doi.org/10.3390/f13071118
Chicago/Turabian StyleMoya, Daniel, Teresa Fonturbel, Esther Peña, Raquel Alfaro-Sanchez, Pedro Antonio Plaza-Álvarez, Javier González-Romero, Manuel Esteban Lucas-Borja, and Jorge de Las Heras. 2022. "Fire Damage to the Soil Bacterial Structure and Function Depends on Burn Severity: Experimental Burnings at a Lysimetric Facility (MedForECOtron)" Forests 13, no. 7: 1118. https://doi.org/10.3390/f13071118
APA StyleMoya, D., Fonturbel, T., Peña, E., Alfaro-Sanchez, R., Plaza-Álvarez, P. A., González-Romero, J., Lucas-Borja, M. E., & de Las Heras, J. (2022). Fire Damage to the Soil Bacterial Structure and Function Depends on Burn Severity: Experimental Burnings at a Lysimetric Facility (MedForECOtron). Forests, 13(7), 1118. https://doi.org/10.3390/f13071118