1. Introduction
The spongy moth (formerly known as the gypsy moth) (
Lymantria dispar) is proficient in triggering severe impairment to ecosystems and individual trees through repetitive defoliation [
1]. This insect is polyphagous, and its caterpillars are particularly destructive defoliators, feeding on over 300 different types of trees [
2,
3] in coniferous and deciduous woodlands [
2,
3,
4,
5]. The preferred species genera are
Quercus,
Populus, Salix [
6,
7]
Pinus and
Acer [
8,
9]. This pest infests many fruit trees, ornamental trees and shrubs [
10], i.e.,
Alnus spp.,
Malus spp.,
Prunus spp.,
Pyrus spp.,
Tilia spp.,
Betula papyrifera,
Crataegus spp.,
Corylus spp.,
Hamamelis spp [
11,
12],
Vaccinium spp.,
Viburnum sp. and
Vitis spp [
7]. Forests are a vital source of sequestering CO
2 from the atmosphere [
13,
14,
15]; unlike many other forest pests that undermine commercial wood values [
16], the spongy moth wreaks havoc on forests, urban forest habitats and inhabitants [
17], causing significant losses to the carbon sink. Due to spongy moth attacks, more geographical areas are affected, which results in the decline of most economical tree species, such as pines and many fruit plants [
18]. Furthermore, outbreaks are more likely in severely degraded forests and woods (canopy cover rate less than 25%) than in natural areas [
19]. Based on females’ ability to fly, where they come from and their mitochondrial DNA, three subspecies of
L. dispar have been found [
20,
21], i.e.,
Lymantria dispar dispar [European spongy moth (ESM)],
Lymantria dispar asiatica [Asian spongy moth (ASM)] and
Lymantria dispar japonica [Japenese spongy moth (JSM)] [
22]. The European spongy moth (ESM) is distributed throughout Eurasia and spreads from the west of Portugal to the eastern mountains of the Ural regions [
23]. In contrast,
L. dispar asiatica occupies much of continental Asia and the eastern areas of Russia, and
L. dispar japonica is exclusively distributed in Japan [
20] and the Kuril Islands of Russia [
22].
L. dispar asiatica is found around the world in a variety of forest environments. Therefore,
L. dispar asiatica may produce progeny with a broader host specificity than the initial populations [
24].
The ESM invaded the US as early as 1869, and the
L. dispar asiatica invaded in 1991 [
25]. However,
L. dispar asiatica has been decimated on at least 20 sites across the United States since its first invasion [
25], but it must be considered that it has more evolved wings [
26] and a higher probability of new invasion than the ESM. Adult females of
L. dispar cannot fly for long periods, limiting the geographical distribution of egg masses [
27]. In addition, compared to
L. dispar asiatica with
L. dispar dispar females, the European spongy moth cannot fly due to more significant abdominal muscles, smaller wings and poorly developed flight muscles [
23,
28]. Therefore, the subspecies
Lymantria dispar dispar (ESM), introduced to North America, has flightless females [
29].
A female spongy moth’s flight capability and frequency are two of the chief criteria used to determine a spongy moth’s proliferation [
30]. Females that fly can deposit their eggs near sharp light sources or bright-colored surfaces [
26,
30]. On the other hand, virgin females fly less frequently than mated females because they must first find a partner for mating [
31]. Instead of confining the investigation to a subspecies level, it is critical to look at flying capabilities at a population level [
30]. Arakawa and Iwaizumi [
32] and Iwaizumi et al. [
33] related the female
L. dispar asiatica’s flight capabilities collected from five locations in Japan. The Tomakomai population could fly the longest distance than the other four localities. Pogue and Schaefer [
22] reviewed
Lymantria and included two sub-species of
Lymantria dispar (
Asiatica and
Japonica) and three different species in Japan (
Lymantria albescens Hori and Umeno,
L. umbrosa Butler and
L. postalba Inoue), and they concluded that the Asian spongy moth could fly.
The attack of the
L. dispar asiatica on forest land in China is severe. Every year, it destroys hundreds of hectares, resulting in thousands of dollars in losses. In the Sichuan basin, spongy moth infestations have caused harm to
Pinus massoniana (pine species) and broadleaf species [
34]. The spongy moth may be found across the arid river valley. Since 2002, forest health assessments and field inspections have seen it in all
Pinus radiata,
P. tabulaeformis and
P. armandii stands [
35]. To conserve forests from spongy moth invasions, it is necessary to study the features of this pest through morphological research, molecular research and flight research. To understand the spread rate of
L. dispar asiatica globally and in China, the flight ability of female
L. dispar asiatica must be studied.
We hypothesized that the age of moths and locality, by which morphological features differ, influence the flight ability of female
L. dispar asiatica in China. We established the layout of the current study to obtained the desire objectives (
Figure 1). To test this hypothesis, flight-related variables of female
L. dispar asiatica from five different geographical locations were investigated in China. Therefore, this research study was conducted to analyze the impact of the locality factor, age effect and mating status on female Asian spongy moths (
Lymantria dispar asiatica).
2. Materials and Methods
2.1. Collection of Egg Masses
Lymantria dispar asiatica egg masses were collected from 5 different geographical locations in China, i.e., the Chengdu, Xifeng, Heilongjiang, Kuduer and Longhua regions [
Figure 2,
Table 1]. The preferred host trees are distributed all around these regions; aside from that, a large
L. dispar asiatica population is present [
28]. Therefore, we selected these geographical locations for our experimental study. Trees from which egg masses were collected belong to the family ‘Pinaceae’ (
Pinus tabuliformis,
P. sylvestris,
P. radiata) and ‘Salicaceae’ (
Populus spp) at each location. Egg masses were then placed into a small PVC container and were transported back to the invasive pest laboratory at Beijing Forestry University for rearing and caring for the insects.
2.2. Rearing of Insects
After arrival in a lab, all the egg masses were kept in an incubator at 4 °C until diapause (inside an incubator, chilling temperature was maintained at 4° to terminate the
L. dispar asiatica; egg diapause took 10 days) was over. When the incubation ended, newly hatched larvae were raised in transparent plastic cups [
Figure 3]. Each cup contained 30 mL of synthetic diet [
36]. Two to four larvae were reared in each plastic cup until larvae pupated. Spongy moth larvae were kept in a laboratory under controlled conditions (at 25 °C), 40%–60% relative humidity (R.H.) and a light: dark (L:D) photoperiod (16:8 h). Before the moth’s emergence, pupae were kept separately in 300 mL clear PVC cups. Moth wings were permitted to swell and stiffen before use. No moth with unexpanded or ruffled wings was used in the experiment.
2.3. Flight Activity Measurement
Adult female Asian spongy moth flight was tracked using a computer-linked flight mill to observe their flight activities (
Figure 4) (Jiaduo Company, Beijing Forestry University, Beijing, China), comparable to those defined by Briegel et al. [
37] and Tanaka and Yamanaka [
38]. Six flight mills were working simultaneously with controlled conditions of temperature and humidity.
The flight mill consisted of four parts: the base, which serves as a support and the flight mill’s balance (reducing gravity pull); two flight arms (one end used for fixing the insect and the other for balancing); and the flight mill itself. A computer chip was set at the base that measures the insect’s movement (how many circles it completes, total distance and maximum speed). We put the Asian spongy moth females on a styrofoam plate and used insect pins to separate their wings from their bodies. With an ink brush, we tried to wipe them off gently. Then, we used super glue (cyanoacrylate) to attach a 2 cm-long piece of steel wire to the thorax of every female. The females were stationary on the foam plate for 3 to 4 minutes to permit the glue to set. Wire parts were linked to the arm of the flight mill with plasticizing, so they were linked to the opposite end of every rotor as counter masses.
Newly emerged moths were retrieved twice daily at 0900 and 1700 h, with age calculated as an average of nights post-eclosion. The influence of age on flying activity was investigated in virgin females 1–3 days following emergence (
n = 30 moths per age class). Mating status’s impact on flying behavior was investigated using age-matched unmated (
n = 30) and mated (
n = 30) females. All the females (unmated and mated) used during the experiment were 1 day old. The emerging females were maintained separately in PVC containers for the virgin treatment. To induce mating, freshly emerging females (30 per population) were housed in a net cage “50 cm × 50 cm × 50 cm” with 40 males for the mated treatment, and females did not start laying eggs until they completed their flight after mating or a scotophase [
36]. A scotophase is a period of darkness, especially one that is artificially imposed (an artificial night).
L. dispar asiatica females did not start laying eggs until they completed their flight after mating or beginning the dark period (night). After mating was observed, the mated females were employed. Every moth was engaged in a single flight mill run (i.e., every moth was only measured once). The moths in the age effects study were not the same as those in the mating status test. Overall flight investigations were carried out at a temperature of 25 °C. The number of spins of the flying arm every 5 s was recorded and utilized to determine the maximum flight speed, minimum flight speed, average flight speed, total flight duration and total flight distance.
2.4. Morphological Features (Thorax and Wing) Measurement
Following the flying test, female wings were cut, unfurled on a horizontal plane and snapped with a camera (Nikon D90) to calculate wing loading [wing loading is a measurement that compares an insect’s mass to its entire wing area. The wing-area-to-body weight proportion is expressed in milligrams per square centimeter (mg/cm2)]. Insect body mass is usually calculated from body length. Each insect body length was calculated using two methods: (i) a vernier caliper and (ii) digital imaging. In our case, we computed the body length with wing size to calculate wing loading. ImageJ (version 1.37 for Windows; National Institutes of Health, Bethesda, MD, USA) was used to draw the wing outline in the digital picture, and the wing areas were transformed from pixels on the inside of the wing outline to square centimeters using a conventional conversion factor. A vernier caliper was used to measure the thorax width.
2.5. Data Analyses
Data were calculated using 30 replications for each location (10 replications for day 1, 10 replications for day 2 and 10 replications for day 3). In total, 5 response variables were calculated, i.e., maximum speed (km/h), minimum speed (km/h), average speed (km/h), flight duration (h) and flight distance (km). Residual data from all five response variables were subjected to a normality probability test (QQ Plot) to check the normal distribution of the data and to a chi-squared test for homogeneity of variance. A two-way ANOVA with two main effects, i.e., location and age, was used to determine the significance of locality and age on the flight activity of the Asian spongy moth in China. Tukey’s HSD all-pairwise comparison tests were employed to compare different parameters at a significance level of p < 0.05. The mean value was calculated for each variable separately. Using a one-way ANOVA, the significance of each variable related to different localities/sites was determined separately. Mean values of day one, day two and day three of each response variable were calculated individually for each locality to calculate the effect of age on L. dispar asiatica flight. Using a two-way ANOVA, the significance of each response variable related to age and different localities/sites was determined separately. A t-test was performed using each response variable’s mean values to check the flight status of unmated and mated females. The mean values of flight distance (km), wing loading (mg/cm2) and thorax width (cm) were calculated for each locality separately; a linear regression model was used to check the association of body size with the flight ability of female L. dispar asiatica. All statistical work was conducted using SPSS Statistics (version 26, IBM Corporation, Armonk, NY, USA), MS-Excel Professional Plus (version 2016, Microsoft, Redmond, WA, USA) and Origin Pro (version 2016, OriginLab Corporation, Northampton, MA, USA).
3. Results
This study examined the association between age, mating status and flying ability in female
L. dispar asiatica from five distinct localities. According to a two-way ANOVA, locality and age significantly impacted all the variables associated with the flight ability of
L. dispar asiatica, i.e., maximum and minimum speed, average speed, flight duration and flight distance (
Table 2). Females from Xifeng were observed fastest with a mean maximum speed of 7.19 ± 0.34 km/h (
Figure 5a). Females from Chengdu were the slowest, with a mean minimum speed of 0.56 ± 0.03 km/h (
Figure 5b). The females from Heilongjiang had the highest values for mean average speed (2.35 ± 0.08 km/h) (
Figure 5c). Females from Xifeng exhibited the maximum flight time (0.94 ± 0.04 h) (
Figure 5d). Moreover, females from Xifeng flew the maximum distance (1.87 ± 0.18 km) (
Figure 5e).
Age significantly impacted flight activity metrics such as speed, flight duration and distance (
Table 3). At 1 day old, all females achieved their longest flying distance, with an average flight distance of 2.64 ± 0.32 km. Later, between 2 and 3 days of age, the flight distances of females dropped. The population from all five localities followed a similar flight speed and duration (
Table 4,
Figure 6). The flight speed of
L. dispar asiatica was the maximum on the first day and decreased on the second and third days (
Table 3). According to the observed results, the mean maximum speed of the female Asian spongy moths was 8.99 ± 0.49 km/h when the adults were 1 day old, and minimum of 3.76 ± 0.87 km/h was observed when the adults were 3 days old. Similarly, the mean minimum speed was higher on the first day (0.84 ± 0.09 km/h) and the lowest on the third day (0.46± 0.11 km/h). The mean average speed was more (2.51 ± 0.43 km/h) when the females were 1 day old and decreased to 1.34 ± 0.31 km/h when it reached the age of 3 days. The mean flight time of the first-day adult Asian spongy moth was 1.09 ± 0.14 h greater than that of the 3rd-day adults (0.54 ± 0.11 h). The mean total flight distance of the Asian spongy moth was 2.64 ± 0.32 km on the first day, which was much greater than the distance covered by an adult having an aging period of 3 days old (0.81 ± 0.24 km) (
Table 3).
There was no significant difference found among the flight variables of unmated and mated females, i.e., average speed (
p = 0.12), flight duration (
p = 0.06) and flight distance (
p = 0.14) (
Table 5). Unmated and mated female
Lymantria dispar asiatica from all five sites reached their maximum average speed, flight time and flight distance on the first day of flight, and they gradually decreased with age (
Figure 6;
Table 4). On the first day of flight, the highest mean maximum speed was 9.62 ± 0.18 km/h for females from Longhua. On the second day, it was 7.66 ± 0.06 km/h for females from Xifeng, and on the third day, it was 4.94 ± 0.61 km/h for females from Kuder (
Figure 6a). The mean minimum speed decreased as the age increased.
The highest value for the first day was 0.89 ± 0.07 km/h for females from Xifeng; for the second and third days, it was 0.84 ± 0.09 km/h and 0.61 ± 0.07 km/h for the females from Heilongjiang (
Figure 6b). On the first day, the flight mean average speed was the maximum with the highest value (2.85 ± 0.10 km/h) for the females from Kuder, which gradually reduced on the second and third days of flight, with the highest values of 2.78 ± 0.35 km/h and 1.88 ± 0.09 km/h for the females from Heilongjiang (
Figure 6c). The highest mean total flight time was observed (1.24 ± 0.06 h) for the females from Xifeng on the first day of the flight. On the second and third days, flight time reduced with maximum values of 0.91 ± 0.06 and 0.91 ± 0.05 h, and it was 0.68 ± 0.06 h for the females from Kuder and Xifeng (
Figure 6d). The age of females limited the mean total flight distance as well. The highest value of the distance traveled was 3.06 ± 0.05 km for the females from Xifeng, which was eventually observed to decrease on the second and third days of flight, with the highest values of 1.95 ± 0.04 km and 1.19 ± 0.09 km for the females from Kuduer and Heilongjiang (
Figure 6e).
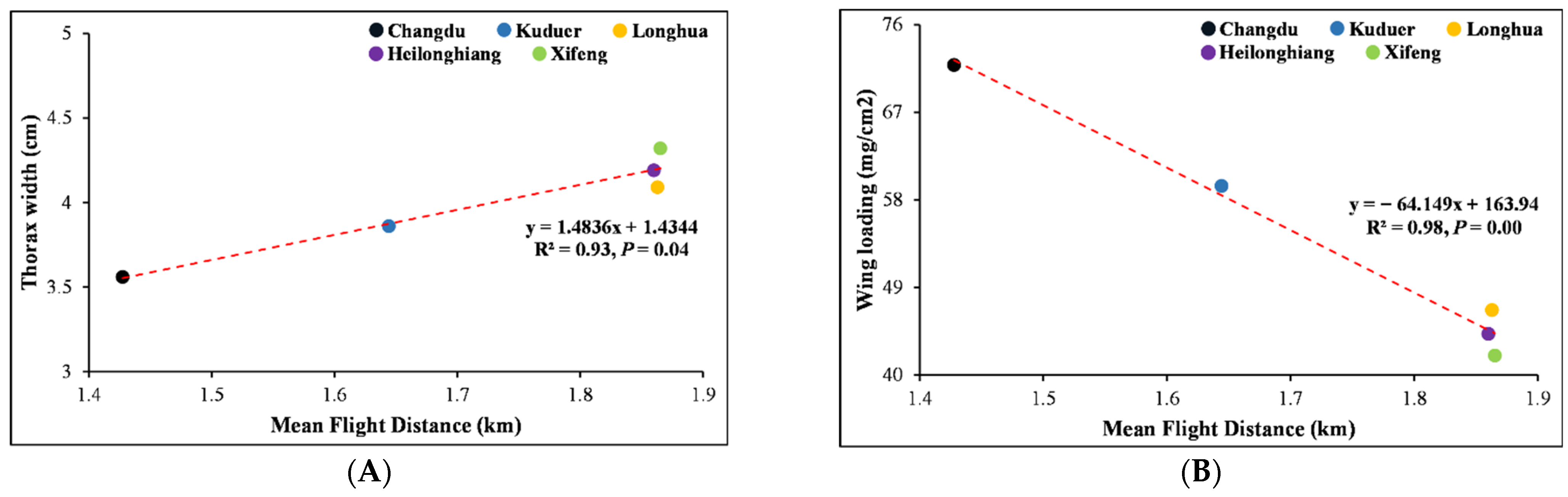
Thorax width and wing loading were also calculated for the females from five different geographical locations (
Table 6). The female population from Xifeng had a maximum thorax width of 4.32 ± 0.22 cm with a minimum wing loading of 41.99 ± 5.09 mg/cm
2, and the female population from Chengdu had a minimum thorax width of 3.56 ± 0.17 cm, with a maximum wing loading of 71.87 ± 5.64 mg/cm
2 (df = 9,
f = 74.98,
p = 0.0001;
p < 0.05). Furthermore, there was a significant linear relationship between body size and female flight ability (
Figure 7). There was positive linear relationship between female thorax width and female flight distance (
y = 1.4836
x + 1.4344,
R2 = 0.93,
p = 0.04;
p < 0.05). Females from Xifeng flew a maximum distance of 1.87 ± 0.18 km due to having more thorax width (4.32 ± 0.22 cm), and females from Chengdu flew the minimum distance (1.43 ± 0.15 km) due to having less thorax width (3.56 ± 0.17 cm). There was negative linear relationship between female wing loading and female flight distance (
y = −64.149
x + 163.94,
R2 = 0.98,
p = 0.00;
p < 0.05). The females from Chengdu with the maximum wing loading (71.87 ± 5.64 mg/cm
2) flew a shorter distance, whereas females from Xifeng with the minimum wing loading (41.99 ± 5.09 mg/cm
2) flew the longest distance.
4. Discussion
This research study aimed to demonstrate a comprehensive approach to investigating the effect of age, locality, mating and morphological features on the flight ability of L. dispar asiatica that could be implemented globally. The results of this research study significantly verified that age and locality influence the abilities of female L. dispar asiatica. Moreover, female populations from different locations have distinct morphological features.
During the experiment, what we have found was significant enough to conclude that age significantly impacted the flight duration and flight ability of female
L. dispar asiatica. Flight activity peaked when the females were one day old and gradually decreased (
Table 4). The species present in the Xifeng location observed the highest maximum speed (9.34 ± 0.10 km/h) on the 1st day, and it continually reduced on 2nd and 3rd days. However, the population from Chengdu showed the lowest range of maximum speed (2.64 ± 0.14 km/h) on the 3rd day (
Figure 6). Yang et al. [
39] found that flight activity peaked when the females of AGM were one day old and then decreased gradually. Chen et al. 2011 [
40] reported that
Dendroctonus armandi (Tsai et Li) flight decreases with aging. Muhammad et al. [
41] reported drop-in flight activity with the age of diamondback moths (
Plutella xylostella Linnaeus).
The physical atrophy of the flight muscles and decreasing enzymatic activity may be linked to the drop-in flight activity as adults age. Our results indicated that the flight ability of female
L. dispar asiatica is directly associated with thorax width and wing loading. Females with wider thoraxes had less wing loading, which considerably improved their capacity to fly, whereas females with the shortest thoraxes had more wing loading, which had an adverse effect on flying (
Figure 7). We also found prominent differences in the morphological features of female
L. dispar asiatica adults (
Table 6). Females from the Xifeng population had the longest flight due to their maximal thorax widths and lower wing loading (
Figure 7). As per our research results, females from Xifeng were capable fliers with a maximum speed of 7.19 ± 0.34 km/h and a total flight distance of more than 1.8 km. Light wing loading (41.99 ± 5.09 mg/cm
2) (
Table 5) helped females from Xifeng to fly long distances. Shirai [
42] investigated the link between flight activity variables and forewing length in
Plutella xylostella and discovered that flight time and distance were proportional to forewing length. Muhammad et al. [
41] revealed that small adults with long forewings of
Plutella xylostella flew more vigorously than large adults with short forewings. Casey [
43] discovered that, with the decrease in the body size, wing stroke frequency rose; hence, smaller insects of similar species exhibited more flying activity. Taylor et al. [
44] discovered that, in
Agrilus planipennis (Fairmaire), the correlation of flying speed with weight load was negative. Shi et al. [
28] conducted a morphometric study on 821 spongy moths from eight multiple strains and reported that average wing loading was much lower, but average wing area was significantly greater for females from flight-proficient populations.
We found no changes in average flight speed, flight duration and flight distance of mated and virgin
L. dispar asiatica females from all five geographically different locations (
Table 5), which is in line with Yang et al. [
39], who reported that mating did not affect the flying attributes of female
L. dispar asiatica.
Geographical locality significantly impacted the flight capability of
L. dispar asiatica. We observed a clear difference between the flight duration and distance among the
L. dispar asiatica population from five different geographical locations in China. The population from Xifeng exhibited a mean maximum flight duration and distance of 0.94 ± 0.04 h and 1.87 ± 0.18 km, the population from Longhua exhibited a mean minimum flight duration of 0.64 ± 0.04 h and the population from Chengdu exhibited a mean minimum flight distance of 1.43 ± 0.15 km. Arakawa and Iwaizumi [
32] and Iwaizumi et al. [
33] investigated the female
L. dispar asiatica flight capabilities collected from five different geographical locations in Japan. They stated that, out of the five locations, the female population from Tomakomai could fly the longest distance compared to those from the other four localities. Iwaizumi et al. [
33] monitored the flying ability of female
L. dispar asiatica using a video camera from five different geographical locations. They stated that the flight distance of Asian spongy moth females from Japan was restricted to 1 km on average. Aside from that, the female population from Tomakomai had a strong flight ability. Baranchikov and Sukachev [
45] reported that female
L. dispar asiatica flew 3–5 km. Yang et al. [
38] stated that the
L. dispar asiatica females from seven different geographical regions had differences in their flight capability. Chen et al. [
46] investigated the flying aptitude of
L. dispar asiatica females from seven nations, finding that 72 percent of the 567 females examined could fly for an extended period, whereas the remainder drifted within a confined spectrum.
We successfully predicted higher flight ability for the female population of L. dispar asiatica from Xifeng compared to those in Kuder, Chengdu, Heilongjiang and Longhua. We also predicted that age and locality would influence female moth flight ability, as morphological features differ with locality. Therefore, we accept our hypothesis and conclude that the flight capability of female L. dispar asiatica is age- and locality-dependent, and the morphological features of L. dispar asiatica are locality-dependent. In addition, mating did not influence female flight ability.
The flight periods perceived in this research study should be measured as maximum probable values. According to our research, adult life spans were no more than 3 days, and only a few lived to the 4th day [
47]. The impromptu conflict was not feasible, and the flight was driven by the absence of a landing base [
48]. The anchored flight distance may not be similar to that of moths in the field. Tethering was predicted to impact flight mill functionality [
44]. As a result, the flight mill system employed in the present research may not describe actual flight patterns. The flight lengths found in this study should be considered optimum values.
Spreading due to flight is an important aspect of the spongy moth life cycle; it is an essential transition from the birth host to a new reproductive host. Spongy moths are economically significant. Therefore, understanding this vital life history feature is essential for calculating the distribution and for managing the tree-killing spongy moth species. However, for females with the ability to fly, these small populations may be higher in numbers outside of commonly infected areas and further away from the edge of infection [
30]. As a result, more research on the
L. dispar asiatica flying behavior in its natural environment is required to comprehend its translocation pathways completely.
Early monitoring and deduction are highly advised. Methods such as predicting potential distributions via a model-based approach [
49] should be used to precisely indicate the suitable habitat for
L. dispar asiatica to develop. Pheromone traps baited with disparlure, cis-7,8-epoxy-2-methyl-octadecane, as used by Wallner et al. [
50], should be used to capture a significant amount of
L. dispar asiatica adults in Beijing and the surrounding areas, and emamectin benzoate (EMB) should be used, as recommended by Zhe at al. [
51], which induces digestive dysfunction and nutrition metabolism disease, which hinders the development of the spongy moth.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}