Abstract
Understanding the factors influencing tree productivity is central to forest ecology. However, the relative contributions of neighborhood interactions, tree species diversity, and tree size to larch (Larix principis-rupprechtii) productivity require further study. Three plots in the Guandi Mountains, Shanxi Province, were set up for each of the following forest types: natural pure larch forest (PL), mixed larch and birch (Betula platyphylla) forest (LB), and mixed larch and spruce (Picea asperata) forest (LS). Based on the tree size-stratified sampling method, a total of 318 tree core samples were collected. A linear mixed model was used to analyze the effects of tree size, dominance, mixing, and neighborhood competition on larch productivity. Birch and spruce promoted larch growth at the stand and individual tree levels, and birch exhibited a more significant facilitating effect. Intraspecific competition was the main factor affecting larch growth. When the intensity of competition among trees was low, the basal area increment (BAI) of larch in the mixed forests was higher than that in the pure forest. However, with increasing competition, the BAI of larch was lower in the mixed forests than in the pure forest. Factors including tree size, dominance, and mingling were positively correlated with the BAI of larch. With increasing tree size, the BAI of larch was higher in the mixed forests than in the pure forest and higher in LB than in LS. When the dominance was less than 0.5, the BAI of larch was higher in the pure forest than in the mixed forests and higher in LS than in LB. With increasing dominance, the BAI of larch was higher in the mixed forests than in the pure forest. The BAI of larch increased with an increasing mixing degree in the mixed forests, and the increasing trend of BAI was larger in LB than in LS. Larch productivity was influenced mainly by neighborhood interactions and stand structure. Improving neighborhood tree diversity and increasing the large tree proportion and dominance of larch will be helpful for improving larch productivity in mixed forests.
1. Introduction
Forest productivity is one of the important indicators of forest quality, and it reflects the growth status of forest trees and the size of ecosystem function to a certain extent [1,2]. In recent decades, the relationship between species diversity and productivity has been an important focus of ecological research [3,4]. Studies have shown that continued reduction in species diversity will lead to an accelerated decline in global forest productivity [5]. Generally, mixed forests tend to have higher stand productivity and exhibit stronger recovery and anti-interference ability [6,7] than pure forests due to a high degree of tree species diversity. Studies have reported positive, negative, and irrelevant relationships between species diversity and productivity; inconsistencies in the results may be caused by differences in tree species interactions, tree composition, tree size, and neighborhood density [8,9,10,11].
The complementarity hypothesis provides a reasonable explanation for a positive correlation between tree species diversity and stand productivity [9,12]. According to this hypothesis, in mixed forests, mixed tree species with different leaf areas, canopy heights and growth rates can obtain and utilize resources more effectively, thus improving stand productivity [13]. However, most studies on tree diversity–productivity relationships have been based on stand-level analyses of diversity effects and have not considered the effect of individual tree-level neighborhood interactions on the relationship between tree diversity and productivity [5,7]. Competition reduction and facilitation are two types of species interactions and have been identified as the key mechanisms that drive the relationship between diversity and productivity; the two species interactions are linked and are often collectively described as complementarity [14,15]. Facilitation emphasizes the positive effects of species in mixed forests, while competition among them decreases due to the differentiation in resource needs (light, water, nutrients) between two or more species [7]. Competition is an important factor affecting tree growth, stand structure, and stand dynamics [16,17]. The intensity of competition between trees is related to the distance between target trees and competition trees and their size. The greater the distance is, the smaller the diameter at breast height (DBH) of the competition wood, and the smaller the competition pressure on the target wood [18]. For a long time, the single-tree competition model has been widely used to study forest tree competition based on distance [19,20].
The productivity effects of tree species interactions are related to tree size [17,21]. Small trees have a limited resource acquisition ability, whereas medium- and large-diameter trees have a stronger positive effect on tree growth [22]. However, the positive relationship between tree size and productivity weakens as the tree age increases [23]. In addition, stand density is also one of the key factors that determines the effect of interactions between tree species on productivity. Studies have shown that when the stand density is low, interactions between trees do not occur or are weak, so complementarity is not significant [7]. With increasing stand density, the interaction is enhanced, and trees can occupy more space and use more resources, resulting in increased complementarity [24]. In fact, interspecific complementarity in light capture has been suggested as a key mechanism by which mixed stands are more productive than monospecific stands [25]. Contrast to tree species in pure forests, coexisting tree species in mixed forests can maximize canopy space filling because of vertical stratification. Therefore, light competition among tree species in mixed forests is lower than that in pure forests [26]. However, when the stand density increases to a certain level and intraspecific or interspecific competition intensifies, stand productivity may decrease [27].
Larch (Larix principis-rupprechtii Mayr) is the main tree species for afforestation and timber in North China, exhibiting a variety of functional benefits. However, at present, the productivity of larch forests is generally low, and stands have been degraded. Most previous studies have focused on the unilateral effects of stand density [28], climatic factors [29], tree species composition, and stand density [11] on larch productivity. However, few studies have considered the effects of neighborhood interactions, species composition, and tree size on larch productivity. We analyzed the effects of neighborhood interaction and stand structure on larch productivity in pure and mixed larch forests to provide a scientific basis for improving the productivity and stand quality of larch.
2. Materials and Methods
2.1. Study Area
The study area was located in Pangquangou National Nature Reserve, Guandi Mountain, Shanxi Province, China (37°45′–37°55′ N, 111°22′–111°33′ E). The average annual temperature is 3 to 7 °C, the average temperature in the growing season is 16.5 °C, and the average annual precipitation is 822.6 mm. The main forest types in the study area are coniferous forest, broad-leaved forest, and mixed coniferous and broad-leaved forest. The main tree species include larch, birch (Betula platyphylla Sukaczev and Betula albosinensis Burkill), and spruce (Picea asperata Mast).
2.2. Sample Site Investigation
Using a stratified random sampling method from July to August 2019, we set up three plots in the pure larch forests (PL), larch–birch mixed forests (LB), and larch–spruce mixed forests (LS) with similar site conditions in the study area, respectively. Each separate stand was randomly distributed in the study area. One plot was set up for each stand. A total of nine plots were designed and surveyed. The plot size was 30 m × 30 m. All woody plants with a DBH larger than 5 cm in the plot were numbered, and the species name, DBH, tree height, relative position coordinates, and other information were determined and recorded (Table 1).

Table 1.
Basic conditions of the sampling plots.
According to the survey data of the sampling plots, at least one sample tree was selected from each diameter class of 2 cm for obtaining tree cores. The number of samples varied from 5 to 16 per plot. The criteria for selecting sample trees were as follows: trees that were large, straight, and disease-free, as well as evenly distributed in the sample plot to the greatest extent possible. Tree cores were extracted from the eastern and northern sides of each tree at a height of 1.3 m using an increment borer, and each core passed through the tree pith so that it provided the complete growth history of the DBH. After the tree core was removed from the increment borer, it was placed in a plastic tube and sealed with paper tape for temporary storage. The location, plot number, tree species, and core number were recorded on a label on the plastic tube. A total of 318 tree core samples were obtained. After the processing and polishing of the tree cores, the annual rings were determined and measured. This measurement revealed the breast diameters of the individual trees in 2008.
2.3. Calculation of Productivity
At the individual tree level, the BAI was calculated using the following formula:
where BAIi is the BAI of the I sampling tree (cm2·year−1) in T; Rn and Rn−T are the radii of n years and n−T years, respectively; T is the research period.
At the stand level, the BAI values of all sampled trees were weighted using the DBHc/DBHss and assigned to trees of the same diameter rank. The DBHc was calculated as the square of the diameter of the target tree. The DBHss represents the mean square diameter of all trees in this diameter order.
2.4. Calculation of Complementarity
To explore the effects of mixing on larch growth, based on Forrester’s method, the complementarity (C) was calculated as follows [14]:
Individual tree level:
Stand level:
where BAIMIX is the BAI of larch in the mixed forest and BAIMONO is the BAI in the pure larch forest. If the complementarity rate is positive, it indicates that the mixed species can promote the growth of larch; otherwise, it can inhibit the growth of larch.
2.5. Calculation of the Competition Index
The competition index can be divided into a competition index unrelated to distance and a competition index related to distance. The widely used Hegyi competition index was adopted in this study, and its calculation formula is as follows [17]:
where CIi represents the competition index of target tree i; CI represents the average competition index; n is the number of competing trees around target tree i; N represents the number of target trees; Dj is the DBH of the competing tree j; Di is the DBH of the target tree i; Lij is the distance between the target tree i and the competition tree j.
As shown in Figure 1, with the increase in the distance between competing trees, the change in the average CI of trees within the radius of 8 m decreased rapidly, while outside the radius of 8 m, the change in the CI was stable, indicating that competing trees outside the radius of 8 m had little influence on the target trees. Therefore, 8 m was identified as the competitive radius in this study.
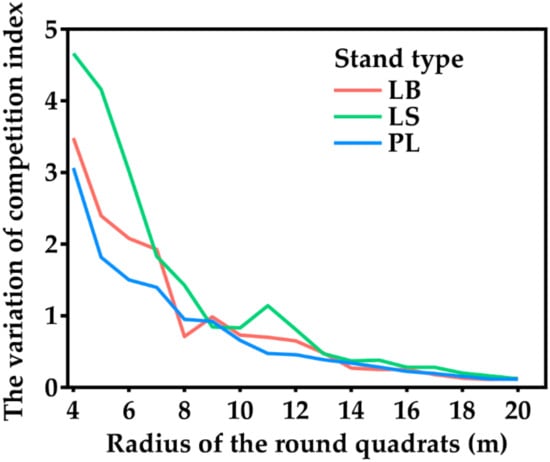
Figure 1.
Relationship between the radius of the round quadrats and the variation in the competition index.
2.6. Calculation of Mixing Index
The mingling index (Mi) was used to reflect the species richness around a particular individual tree [30].
where i represents the target tree, and n indicates the number of nearest neighbors included in the analysis (n = 4 in this study); vij is 0 if the neighbor j belongs to the same species as the target tree i or 1 if the neighbor j belongs to a different species from the target tree i.
2.7. Calculation of Dominance Index
The dominance index (Ui) reflects the relative dominance of an object tree in its neighborhood and is calculated as follows [31]:
where n represents the number of neighbor individuals of the target tree within an 8 m radius. If the DBH of the neighbor j is greater than that of the target tree i, kij is equal to 0. If the DBH of the neighbor j is less than or equal to the target tree I, kij is equal to 1.
2.8. Construction of the Linear Mixed Model
Based on the survey data, the DBH, intraspecific competition CIA, interspecific competition CIB, dominance index U, neighboring density Dens, and mingling index M were calculated. The results of a Pearson correlation analysis of these factor variables with BAI show that intraspecific competition significantly affected productivity in LB, but interspecific competition did not. In LS, interspecific competition significantly affected productivity, but intraspecific competition did not. The variance inflation factor (VIF) value of factor variables was calculated by linear regression to ensure that there was no multicollinearity between the variables and that the VIF values were all <10. In this study, logBAI was used as the dependent variable to fit the linear mixed model of a single-tree BAI for larch in different stands, and the determination coefficient (R2) was used to test the accuracy of the model to explore the radial growth limiting factor of larch in the Guandi Mountain forest region and its effect on BAI. The fitting of the linear model was carried out in the “nlme” software package in R 4.1.2 [32]. The interaction of the intraspecific CIA, dominance index U, DBH, and mingling index M on BAI was considered in LB. To remove the effect of spatial autocorrelation among the trees within an individual plot, we treated the plot and trees as nested random effects. The fitting form of the linear mixed model is as follows:
logBAIi = β0 + β1DBH + β2CIAi + β3CIBi + β4Ui + β5Mi +β7 (CIAi:Ui) + β8 (DBH: Mi)
In LS, the interaction effect of the neighborhood density Densi and tree species mixing index Mi on BAI was considered. In addition, the plot and trees as nested random effects were considered. The fitting form of the linear mixing model is as follows:
where BAIi is the BAI of the target tree i in the past 10 years (2008–2018) (cm2·year−1); DBH is the diameter at breast height of the target tree; CIAi is the intraspecific competition index of the target tree i; CIBi is the index of interspecific competition of the target tree i; Ui is the dominance index of the target tree i; Densi is the sum of the basal area of the four nearest neighbor trees of the target tree i; Mi is the index of the neighborhood mixing degree of the target tree i; DBH: Mi is the interaction between the DBH of the target tree i and the mixing index of its neighbors. Densi: Mi is the interaction between the neighborhood density and the neighborhood mingling index of the target tree i, and β0~β8 are the parameters to be estimated.
logBAIi = β0 + β1DBH + β2CIAi + β3CIBi + β4Ui + β5Mi + β6 (Densi) + β7 (Densi: Mi)
The calculation of the competition index and the fitting of the linear mixed model were carried out in R 4.1.2.
3. Results
At the level of individual trees, the productivity (BAI) of larch in the mixed forests was higher than that in the pure forest, and the BAI of larch in LB was higher than that in LS (Table 2). The complementarity ratio of larch was positive at both the individual and stand levels in mixed forests, indicating that both birch and spruce promoted larch growth, but birch had a more significant positive effect. In terms of the competition index, the competition of spruce to larch was higher than that of birch. At the individual tree level, intraspecific competition was greater than interspecific competition in both LB and LS.
Table 2.
Productivity, complementarity, and competition indices of tree species in different stand types.
The BAI was positively correlated with dominance in both the pure and mixed forests (p < 0.01) (Figure 2). When the dominance was low (Ui < 0.5), the BAI of the pure forest was higher than that of the mixed forests, and it was higher for LS than for LB. With the increase in dominance (Ui ≥ 0.5), the BAI of the mixed forests was slightly higher than that of the pure forest, especially for LB. In both the pure forest and mixed forests, the BAI decreased significantly with an increasing competition index (p < 0.01). The BAI of the mixed forests was higher than that of the pure forest when the competition among trees was small, and the BAI of the pure forest was higher than that of the mixed forests as the competition intensified. In both the pure and mixed forests, the BAI increased significantly with increasing tree size (p < 0.01), and LB > LS > PL. In the mixed forest, the BAI increased with an increase in mingling, and LB > LS.
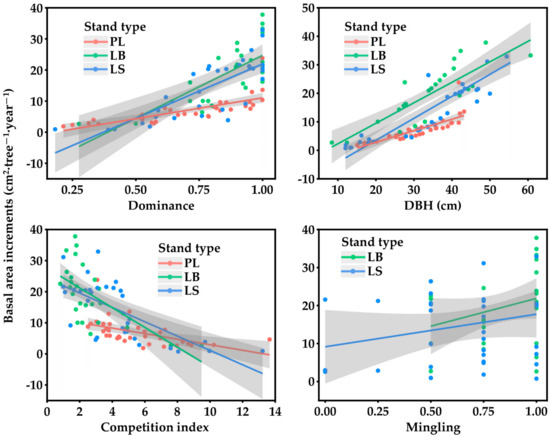
Figure 2.
Individual basal area increments in different larch stand types varied with dominance, DBH, competition index, and mingling. Grey areas represent 95% confidence interval for each stand type.
In LB, the DBH, dominance, mingling, and the interaction of the intraspecific competition and dominance were significantly positively correlated with the BAI (p < 0.05) (Table 3), and the BAI was negatively correlated with the intraspecific competition, DBH, and mingling (p < 0.01). In LS, interspecific competition and the interaction of neighborhood density and mingling exhibited a significant negative correlation with BAI (p < 0.05), while dominance and mingling exhibited a significant positive correlation with BAI (p < 0.05).
Table 3.
Parameter estimations of the linear mixed model of the individual basal area increments in larch mixed forests. (ns): p > 0.05; (*): p < 0.05; (**): p < 0.01; (***): p < 0.001.
4. Discussion
In general, mixed forests tend to have higher stand productivity than pure forests due to high tree species diversity [33,34]. In this study, larch productivity in the mixed forests was higher than that in the pure forest at both the individual and stand levels (Table 2). This finding is consistent with those of most studies [3,35]. At the neighborhood level, neighboring heterospecies can promote tree growth [33]. In the mixed larch–birch and larch–spruce forests, the individual productivity of larch increased with increasing mingling (Figure 2, Table 3), which is consistent with the results of previous studies [36,37]. The relationship between mingling and productivity depends on the net effect between the target tree and the neighboring tree species, and when facilitation dominates, it will have a positive effect [37].
The complementary effect hypothesis states that due to niche differentiation and promoting interactions, tree species are complementary in resource acquisition and utilization, thus improving productivity [38,39]. The results show that the complementarity rate of larch–birch mixed forest was higher than that of larch–spruce mixed forest at both the individual and stand levels (Table 2). This may be due to canopy structure differences that enable trees to make full use of light, water, and other resources and thus increase productivity in larch–birch mixed forests [40,41]; in contrast, larch and spruce are both middle- and late-succession species, with similar spatial niches and fewer complementary effects [42]. Neighborhood interspecific interactions largely determine tree growth rates [43]. In this study, we found that when the CI was less than 7, the individual productivity of larch in the mixed forest was higher than that in the pure forest (Figure 2). Birch and spruce promoted larch, and the competition that occurred in the mixed forests was mainly from intraspecific competition (Table 1). However, with the intensification of competition among trees, the productivity of the pure forest was higher than that of the mixed forests due to limited resources and competition among tree species (Figure 2). The interaction is related to the life history strategies of different tree species. When the competition intensity is low, acquisitive tree species benefit from more diverse neighborhoods, resulting in less competition. Conserved tree species lead to promotion when competition intensity is high [19,43]. How the mixing of larch with different species affects forest productivity remains a topic for further study.
Tree size is a key factor affecting tree growth [44]. This study showed that tree size had a significant positive effect on larch productivity at the individual tree level (Figure 2, Table 3), which is similar to the results of other studies [20]. Larger trees usually occupy a dominant position in the canopy, have easier access to light, and thus, have higher growth rates [45,46]. However, the relationship between tree size and individual tree productivity also varies with species composition. Forrester [40] showed that the influence of mixed tree species on light absorption was closely related to the vertical canopy structure, stand density, the presence of deciduous species, tree diameter, height, the allometric relationship between crowns, and other factors. Vertical stratification and canopy plasticity varied with tree composition, stand structure, and age [47]. This study showed that larch was more productive in mixed forests than in pure forests (Table 2). It may be because the canopy of larch in mixed forests has more space to extend and receive more light than that in pure forests. With the increase in tree size, the individual productivity of larch was always higher in the larch–birch mixed forest than in the larch–spruce mixed forest (Figure 2). It is likely that due to differences in the succession status and biological characteristics among tree species, tall larch tends to compete asymmetrically with birch relative to spruce, capturing a disproportionate increase in light relative to its size [43] and leading to higher productivity. However, when the dominance of larch was low, the productivity of larch was higher in the larch–spruce mixed forest than in the larch–birch mixed forest (Figure 2). This is probably because birch is a typical pioneer tree species in secondary succession [48], which grows rapidly in the early stage and has a higher competitive advantage against larch compared to spruce; larch growth is inhibited, and productivity is lower. As the dominance of larch increases, it gradually benefits from more resources, such as light, water, and nutrients, and from the reduced competition with birch due to niche complementarity [19]. Compared to spruce, larch had a more obvious competitive advantage over birch, resulting in a greater increase in productivity in the larch–birch mixed forest (Figure 2).
5. Conclusions
This study reveals that tree species diversity, neighborhood interactions, tree size, dominance, and mingling had significant effects on larch productivity. In mixed forests, birch and spruce both promoted larch growth due to niche complementarity and reduced competition, but birch had a more obvious positive effect on larch growth. The complementarity effect varied with tree competition and dominance. The individual productivity of larch in the mixed forests was higher than that in the pure forest when the competition between trees was low, the competition between species was intensified when the competition between trees was high, and the individual productivity of larch in the pure forest was higher than that in the mixed forests. When larch dominance was low, the complementary effect was not obvious, and the productivity of larch in the pure forest was higher than that in the mixed forests. With the increase in larch dominance, larch exhibited competitive advantages, especially in the larch–birch mixed forest. Larch productivity increased with the increase in mingling in the neighborhood, especially in the larch–birch mixed forest. In the management of larch forests, it is beneficial to improve larch productivity by constructing mixed larch and birch forests, increasing the diversity of tree species, and increasing the large tree proportion and dominance of larch.
Author Contributions
Conceptualization, J.Z. (Jing Zhang) and Z.Z.; methodology, software, visualization, J.Z. (Jing Zhang) and Z.G.; investigation, data curation, J.Z. (Jing Zhang), J.Z. (Jinman Zhao), and R.C.; writing—original draft preparation, J.Z. (Jing Zhang); writing—review and editing, Z.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Natural Science Foundation of China, grant number 32071759, the Natural Science Foundation of Hebei Province, China, grant number C2020204026, and the Hebei Province Key R & D Program of China, grant number 22326803D.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is available upon request to the corresponding authors.
Acknowledgments
The authors thank everyone who helped with the field survey and the anonymous reviewers for their valuable comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Huang, S.-L.; Ramirez, C.; Conway, S.; Kennedy, K.; Kohler, T.; Liu, J.-X. Mapping site index and volume increment from forest inventory, landsat, and ecological variables in Tahoe National Forest California. Can. J. Forest Res. 2016, 47, 113–124. [Google Scholar]
- Hansson, K.; Laclau, J.-P.; Saint-André, L.; Mareschal, L.; van der Heijden, G.; Nys, C.; Nicolas, M.; Ranger, J.; Legout, A. Chemical fertility of forest ecosystems. Part 1: Common soil chemical analyses were poor predictors of stand productivity across a wide range of acidic forest soils. For. Ecol. Manag. 2020, 461, 117843. [Google Scholar] [CrossRef]
- Lu, H.-C.; Mohren, G.M.J.; del Río, M.; Schelhaas, M.-J.; Bouwman, M.; Sterck, F.J. Species Mixing Effects on Forest Productivity: A case study at stand-, species- and tree-level in the Netherlands. Forests 2018, 9, 713. [Google Scholar] [CrossRef]
- Zeler, L.; Pretzsch, H. Effect of forest structure on stand productivity in Central European forests depends on developmental stage and tree species diversity. For. Ecol. Manag. 2019, 434, 193–204. [Google Scholar] [CrossRef]
- Liang, J.-J.; Crowther, T.W.; Picard, N.; Wiser, S.K. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, aaf8957. [Google Scholar] [CrossRef]
- Poorter, L.; van der Sande, M.T.; Thompson, J.; Arets, E.J.M.M.; Alarcon, A.; Alvarez-Sanchez, J.; Ascarrunz, N.; Balvanera, P.; Barajas-Guzman, G.; Boit, A.; et al. Diversity enhances carbon storage in tropical forests. Global. Ecol. Biogeogr. 2015, 24, 1314–1328. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J. A review of processes behind diversity-productivity relationships in forests. Curr. For. Rep. 2016, 2, 45–61. [Google Scholar] [CrossRef]
- Grace, J.B.; Anderson, T.M.; Smith, M.D.; Seabloom, E.; Andelman, S.J.; Meche, G.; Weiher, E.; Allain, L.K.; Jutila, H.; Sankaran, M.; et al. Does species diversity limit productivity in natural grassland communities? Ecol. Lett. 2007, 10, 680–689. [Google Scholar] [CrossRef]
- Van de Peer, T.; Verheyen, K.; Ponette, Q.; Setiawan, N.N.; Muys, B. Overyielding in young tree plantations is driven by local complementarity and selection effects related to shade tolerance. J. Ecol. 2018, 106, 1096–1105. [Google Scholar] [CrossRef]
- Sun, H.-G.; Diao, S.-F.; Liu, R.; Forrester, D.; Soares, A.; Saito, D.; Dong, R.-X.; Jiang, J.-M. Relationship between size inequality and stand productivity is modified by self-thinning, age, site and planting density in Sassafras tzumu plantations in central China. For. Ecol. Manag. 2018, 422, 199–206. [Google Scholar] [CrossRef]
- Torresan, C.; del Río, M.; Hilmers, T.; Notarangelo, M.; Bielak, K.; Binder, F.; Boncina, A.; Bosela, M.; Forrester, D.I.; Hobi, M.L.; et al. Importance of tree species size dominance and heterogeneity on the productivity of spruce-fir-beech mountain forest stands in Europe. For. Ecol. Manag. 2019, 457, 117716. [Google Scholar] [CrossRef]
- Madrigal-González, J.; Ruiz-Benito, P.; Ratcliffe, S.; Calatayud, J.; Kändler, G.; Lehtonen, A.; Dahlgren, J.; Wirth, C.; Zavala, M.A. Complementarity effects on tree growth are contingent on tree size and climatic conditions across Europe. Sci. Rep. 2016, 6, 32233. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Knops, J.; Wedin, D.; Reich, P.; Ritchie, M.; Siemann, E. The influence of functional diversity and composition on ecosystem processes. Science 1997, 277, 1300–1302. [Google Scholar] [CrossRef]
- Forrester, D.I. The spatial and temporal dynamics of species interactions in mixed-species forests: From pattern to process. For. Ecol. Manag. 2014, 312, 282–292. [Google Scholar] [CrossRef]
- Fichtner, A.; Härdtle, W.; Li, Y.; Bruelheide, H.; Kunz, M.; von Oheimb, G. From competition to facilitation: How tree species respond to neighbourhood diversity. Ecol. Lett. 2017, 20, 892–900. [Google Scholar] [CrossRef] [PubMed]
- Von Oheimb, G.; Lang, A.C.; Bruelheide, H.; Forrester, D.I.; Wäsche, I.; Yu, M.-J.; Härdtle, W. Individual-tree radial growth in a subtropical broad-leaved forest: The role of local neighbourhood competition. For. Ecol. Manag. 2011, 261, 499–507. [Google Scholar] [CrossRef]
- Forrester, D.I. Linking forest growth with stand structure: Tree size inequality, tree growth or resource partitioning and the asymmetry of competition. For. Ecol. Manag. 2019, 447, 139–157. [Google Scholar] [CrossRef]
- Hegyi, F. A simulation model for managing jack-pine stands. In Growth Models for Tree and Stand Simulation; Fries, J., Ed.; Royal College of Forestry: Stockholm, Sweden, 1974; pp. 74–90. [Google Scholar]
- Wu, C.-Y.; Chen, D.-S.; Sun, X.-M.; Zhang, S.-G. Contributions of competition on Larix kaempferi tree-ring growth were higher than long-term climate in China. Agric. Forest Meteorol. 2022, 320, 108967. [Google Scholar] [CrossRef]
- Fan, C.-Y.; Tan, L.-Z.; Zhang, C.-Y.; Zhao, X.-H.; Gao, L.-S.; von Gadow, K. Scale-dependent effects of neighborhood biodiversity on individual tree productivity in a coniferous and broad-leaved mixed forest in China. Ecol. Evol. 2020, 10, 8225–8234. [Google Scholar] [CrossRef]
- Soares, A.A.V.; Leite, H.G.; Souza, A.L.; Silvab, S.R.; Lourenco, H.M.; Forresterd, D.I. Increasing stand structural heterogeneity reduces productivity in Brazilian Eucalyptus monoclonal stands. For. Ecol. Manag. 2016, 373, 26–32. [Google Scholar] [CrossRef]
- Wright, A.; Schnitzer, S.A.; Reich, P.B. Living close to your neighbors: The importance of both competition and facilitation in plant communities. Ecology 2014, 95, 2213–2223. [Google Scholar] [CrossRef] [PubMed]
- Canham, C.D.; LePage, P.T.; Coates, K.D. A neighborhood analysis of canopy tree competition: Effects of shading versus crowding. Can. J. Forest Res. 2004, 34, 778–787. [Google Scholar] [CrossRef]
- Morin, X. Species richness promotes canopy packing: A promising step towards a better understanding of the mechanisms driving the diversity effects on forest functioning. Funct. Ecol. 2015, 29, 993–994. [Google Scholar] [CrossRef]
- Morin, X.; Fahse, L.; Scherer-Lorenzen, M.; Bugmann, H. Tree species richness promotes productivity in temperate forests through strong complementarity between species. Ecol. Lett. 2011, 14, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Pretzsch, H. Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures. For. Ecol. Manag. 2014, 327, 251–264. [Google Scholar] [CrossRef]
- Brunner, A.; Forrester, D.I. Tree species mixture effects on stem growth vary with stand density—An analysis based on individual tree responses. For. Ecol. Manag. 2020, 473, 118334. [Google Scholar] [CrossRef]
- Forrester, D.I.; Kohnle, U.; Albrecht, A.T.; Bauhus, J. Complementarity in mixed-species stands of Abies alba and Picea abies varies with climate, site quality and stand density. For. Ecol. Manag. 2013, 304, 233–242. [Google Scholar] [CrossRef]
- Rohner, B.; Waldner, P.; Lischke, H.; Ferretti, M.; Thürig, E. Predicting individual-tree growth of central European tree species as a function of site, stand, management, nutrient, and climate effects. Eur. J. Forest Res. 2018, 137, 29–44. [Google Scholar] [CrossRef]
- Gadow, K.V.; Zhang, C.Y.; Wehenkel, C.; Pommerening, A.; Corral-Rivas, J.; Korol, M.; Myklush, S.; Hui, G.Y.; Kiviste, A.; Zhao, X.H. Forest structure and diversity. In Managing Forest Ecosystems; Springer: Berlin/Heidelberg, Germany, 2012; pp. 29–83. [Google Scholar]
- Ni, R.-Q.; Baiketuerhan, Y.; Zhang, C.-Y.; Zhao, X.-H.; von Gadow, K. Analysing structural diversity in two temperate forests in northeastern China. For. Ecol. Manag. 2014, 316, 139–147. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Heisterkamp, S.; Van Willigen, B. Linear and Nonlinear Mixed Effects Models. R Package 2020, 3, 1–336. [Google Scholar]
- Houpert, L.; Rohner, B.; Forrester, D.I.; Mina, M.; Huber, M.O. Mixing effects in Norway spruce-European beech stands are modulated by site quality, stand age and moisture availability. Forests 2018, 9, 83. [Google Scholar] [CrossRef]
- Dănescu, A.; Albrecht, A.T.; Bauhus, J. Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany. Oecologia 2016, 182, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Fien, E.K.P.; Fraver, S.; Teets, A.; Weiskittela, A.R.; Hollinger, D.Y. Drivers of individual tree growth and mortality in an uneven-aged, mixed-species conifer forest. For. Ecol. Manag. 2019, 449, 117446. [Google Scholar] [CrossRef]
- Ding, Y.; Zang, R.-G. Determinants of aboveground biomass in forests across three climatic zones in China. For. Ecol. Manag. 2020, 482, 118805. [Google Scholar] [CrossRef]
- Wu, Z.-F.; Fan, C.-Y.; Zhang, C.-Y.; Zhao, X.-H.; von Gadow, K. Effects of biotic and abiotic drivers on the growth rates of individual trees in temperate natural forests. For. Ecol. Manag. 2021, 503, 119769. [Google Scholar] [CrossRef]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Forrester, D.I.; Ammer, C.; Annighöfer, P.J.; Barbeito, I.; Bielak, K.; Bravo-Oviedo, A.; Coll, L.; del Río, M.; Drössler, L.; Heym, M.; et al. Effects of crown architecture and stand structure on light absorption in mixed and monospecific Fagus sylvatica and Pinus sylvestris forests along a productivity and climate gradient through Europe. J. Ecol. 2017, 106, 746–760. [Google Scholar] [CrossRef]
- Sapijanskas, J.; Paquette, A.; Potvin, C.; Kunert, N.; Loreau, M. Tropical tree diversity enhances light capture through crown plasticity and spatial and temporal niche differences. Ecology 2014, 95, 2479–2492. [Google Scholar] [CrossRef]
- Sterba, H.; Drinberger, G.; Ritter, T. The contribution of forest structure to complementarity in mixed stands of Norway spruce (Picea abies L. Karst) and European larch (Larix decidua Mill.). Forests 2018, 9, 410. [Google Scholar] [CrossRef]
- Fichtner, A.; Härdtle, W.; Bruelheide, H.; Kunz, M.; Li, Y.; von Oheimb, G. Neighbourhood interactions drive overyielding in mixed-species tree communities. Nat. Commun. 2018, 9, 1144. [Google Scholar] [CrossRef] [PubMed]
- Mensah, S.; Veldtman, R.; Toit, B.D.; Kakaï, R.G.; Seifert, T. Aboveground biomass and carbon in a south african mistbelt forest and the relationships with tree species diversity and forest structures. Forests 2016, 7, 79. [Google Scholar] [CrossRef]
- Augusto, L.; Schrijver, A.D.; Vesterdal, L.; Smolander, A.; Prescott, C.; Ranger, J. Influences of evergreen gymnosperm and deciduous angiosperm tree species on the functioning of temperate and boreal forests. Biol. Rev. Camb. Philos. Soc. 2015, 90, 444–466. [Google Scholar] [CrossRef] [PubMed]
- Gersonde, R.; Battles, J.J.; O’Hara, K.L. Characterizing the light environment in Sierra Nevada mixed-conifer forests using a spatially explicit light model. Can. J. Forest Res. 2004, 34, 1332–1342. [Google Scholar] [CrossRef]
- Ishii, H.; Azuma, W.; Nabeshima, E. The need for a canopy perspective to understand the importance of phenotypic plasticity for promoting species coexistence and light-use complementarity in forest ecosystems. Ecol. Res. 2013, 28, 191–198. [Google Scholar] [CrossRef]
- Wu, B.; Lian, C.; Hogetsu, T. Development of microsatellite markers in white birch (Betula platyphylla var. japonica). Mol. Ecol. Resour. 2002, 2, 413–415. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).