

Net Primary Productivity of Forest Ecosystems in the Southwest Karst Region from the Perspective of Carbon Neutralization


Abstract
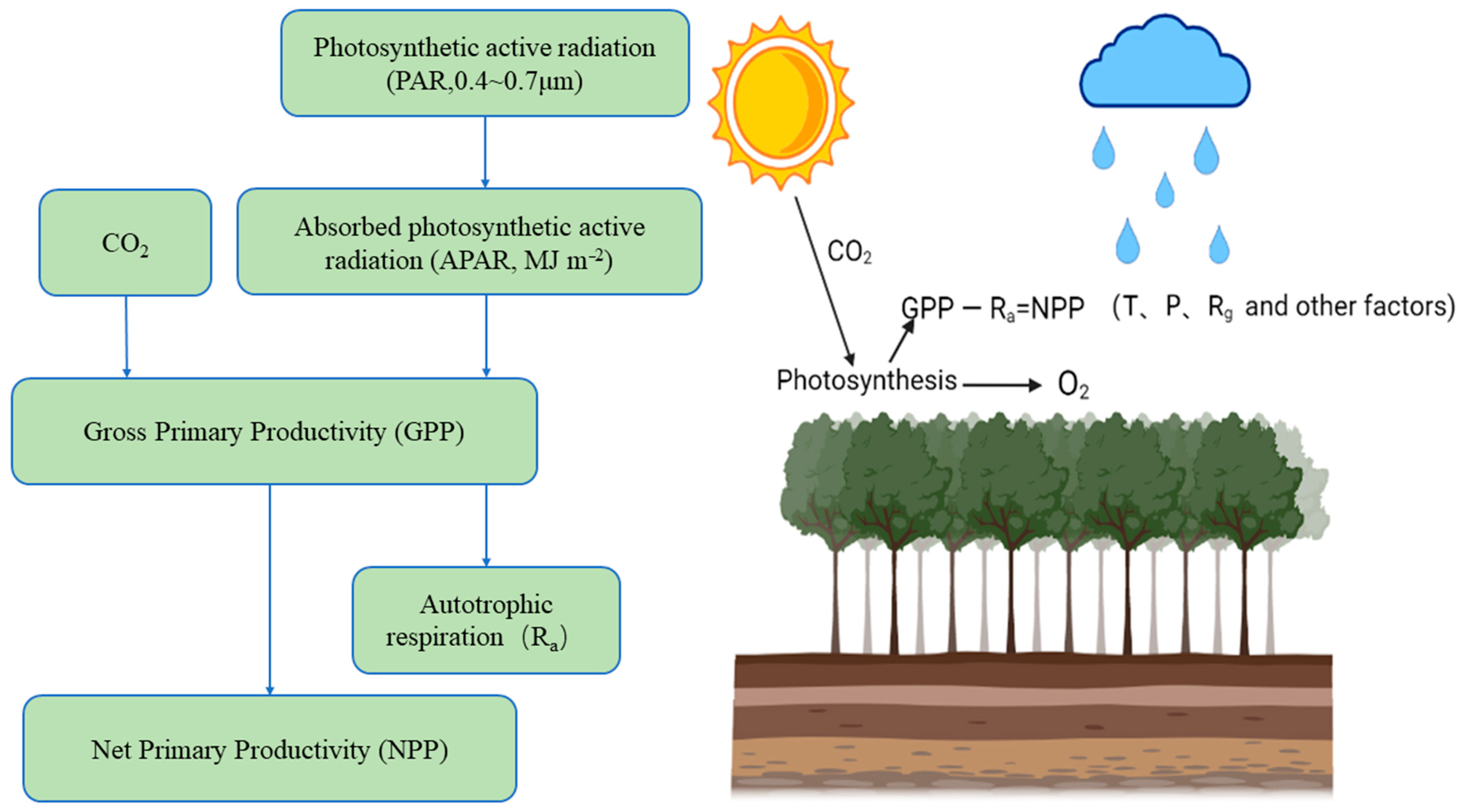
:1. Preface
2. Research Progress on NPP in Karst Areas of Southwest China
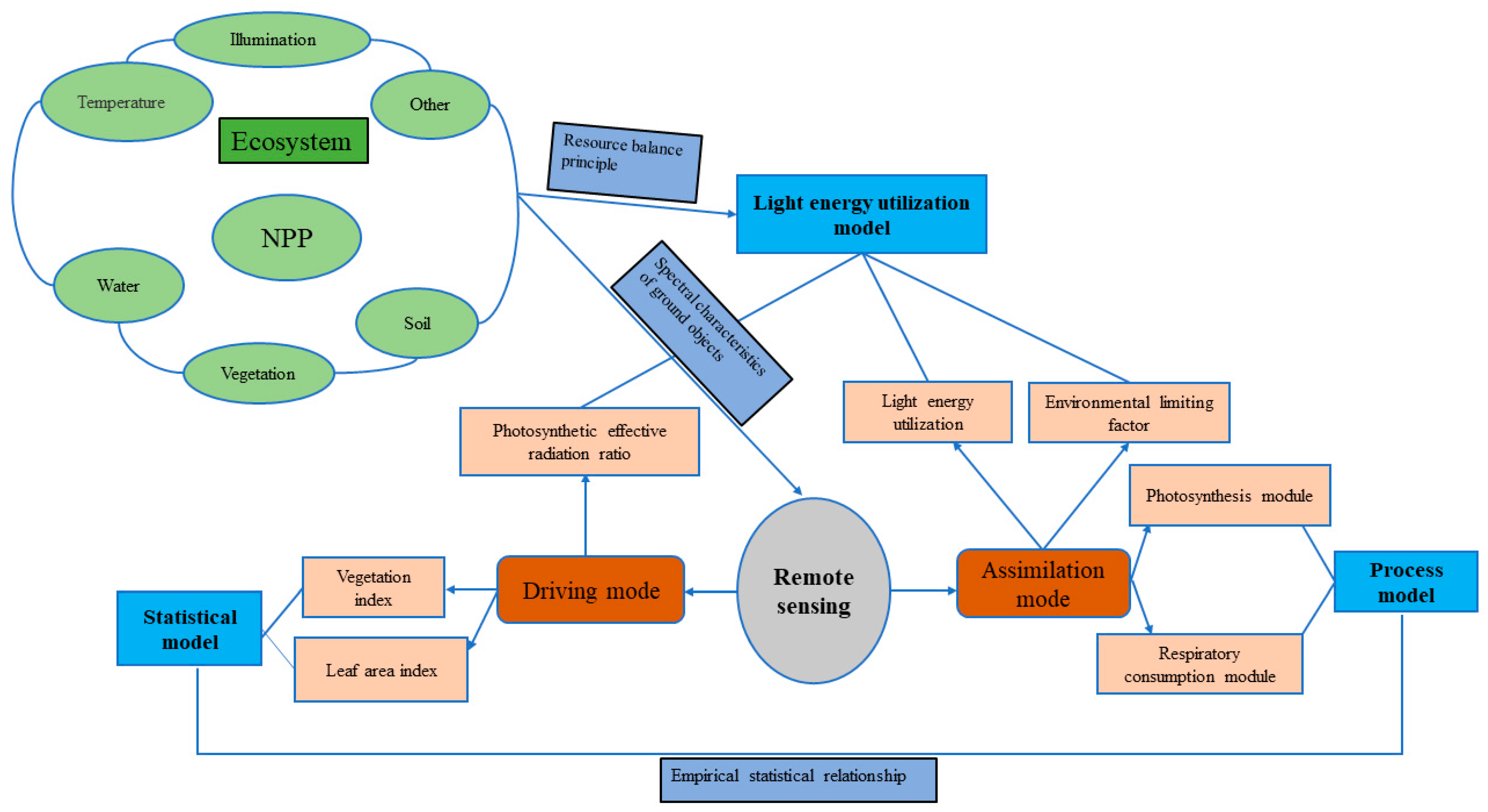
2.1. Overview of NPP Research Models
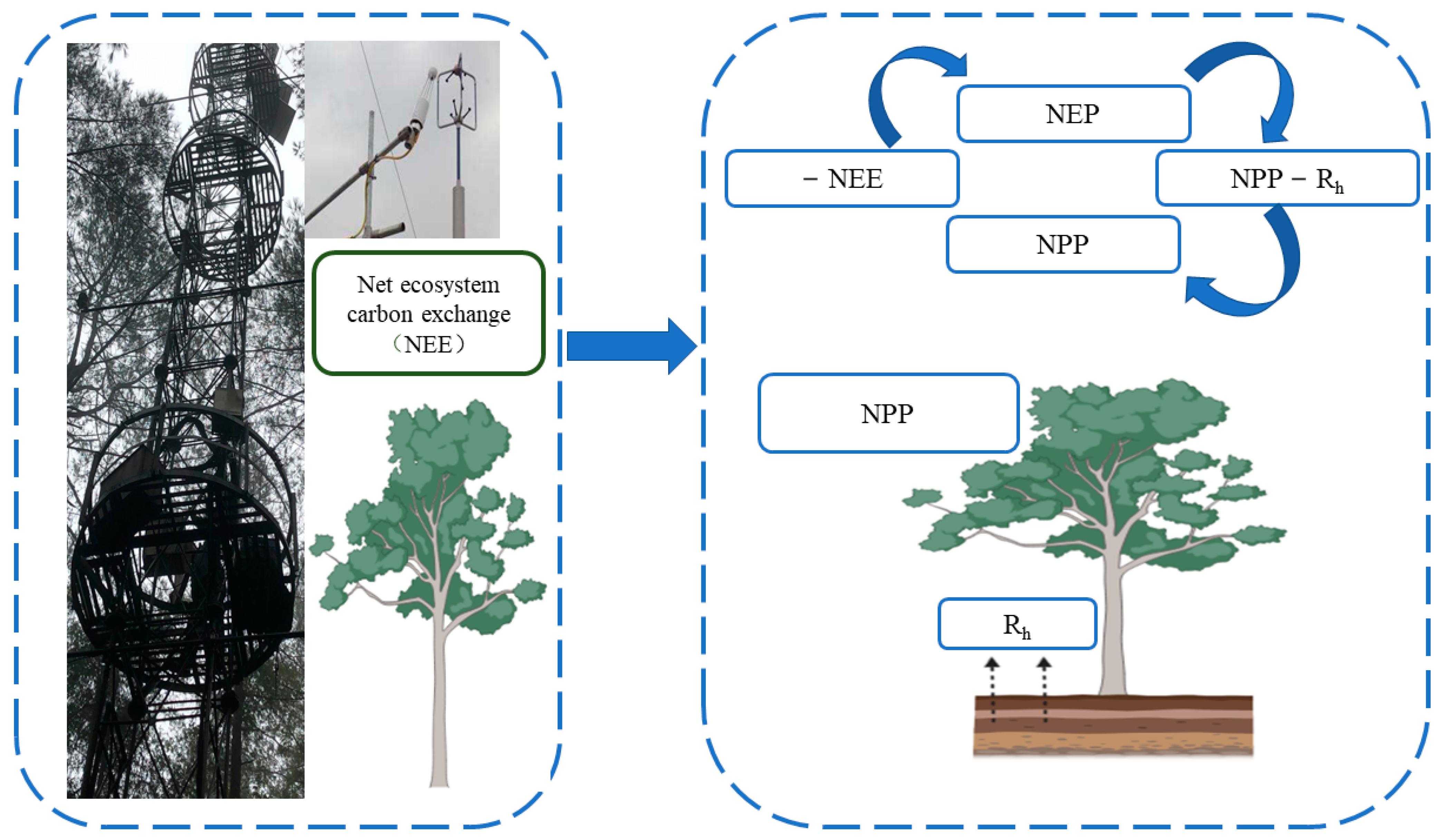
2.1.1. Field Observation Method
2.1.2. Statistical Model
2.1.3. Process Model
2.1.4. Light Energy Utilization Model
2.2. Response of the NPP to Climate Change in Karst Areas of Southwest China
2.2.1. Effect of Temperature Change on NPP
2.2.2. Impact of Precipitation Change on NPP
2.2.3. Impact of VPD on NPP
3. Current Situation and Change Trend of the NPP of Forest Ecosystems in Karst Areas
4. Summary and Perspectives
4.1. Establishment of a High-Precision and Comprehensive Coupling Model
4.2. Developing High-Precision Climate Data Sets
4.3. Further Research on Interaction Mechanisms
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Year | Reference | Source | Website/Doi |
|---|---|---|---|---|
| Hong, C., et al. | 2017 | [11] | Progress in Geographical Science | https://cnki.net/ |
| Lieth, H. | 1975 | [18] | Primary Productivity of the Biosphere | https://doi.org/10.1007/978-3-642-80913-2_12 |
| Lieth, H. | 1972 | [19] | pub in climatology | https://www.researchgate.net/publication/284261627 |
| Zenbei, U. and S. Hiroshi | 1985 | [20] | Journal of Agricultural Meteorology | https://doi.org/10.2480/agrmet.40.343 |
| Zhou, G. and X. Zhang | 1995 | [21] | Journal of plant ecology | https://cnki.net/ |
| Shen, B. | 2019 | [39] | Chinese Academy of Agricultural Sciences | https://cnki.net/ |
| Zhu, W. | 2005 | [43] | Beijing Normal University | https://cnki.net/ |
| Piao, S., J. Fang, and Q. Guo | 2001 | [44] | Journal of plant ecology | https://cnki.net/ |
| Sun, C., et al. | 2013 | [53] | Journal of Yangzhou University | https://doi.org/10.16872/j.cnki.1671-4652.2013.04.012 |
| Sun, J., H. Niu, and M. Yuan | 2020 | [54] | Journal of agricultural machinery | https://cnki.net/ |
| Sun, Z. and S. Xie | 2021 | [55] | Journal of ecology | https://doi.org/10.13292/j.1000-4890.202111.033 |
| Wu, G. | 2015 | [56] | Nanjing Forestry University | https://cnki.net/ |
| Zhao, M., et al. | 2019 | [57] | Journal of ecological environment | https://doi.org/10.16258/j.cnki.1674-5906.2019.02.001 |
| Li, M., et al | 2019 | [58] | Environmental Science Research | https://doi.org/10.13198/j.issn.1001-6929.2018.11.14 |
| Hong, X., Y. Huang, and T. SUn | 2021 | [59] | Journal of ecology | https://cnki.net/ |
| Lin, X., et al. | 2015 | [60] | Environmental Protection Science | https://doi.org/10.16803/j.cnki.issn.1004-6216.2015.02.020 |
| Xiong, X., et al. | 2019 | [61] | Journal of Guangxi Normal University | https://doi.org/10.16088/j.issn.1001-6600.2019.03.022 |
| Xu, Y., Y. Zhang, and W. Pan | 2018 | [62] | Environment and sustainable development | https://doi.org/10.19758/j.cnki.issn1673-288x.2018.01.024 |
| Zhao, Z., et al. | 2015 | [63] | Journal of ecology | https://cnki.net/ |
| Zhang, R., et al. | 2015 | [64] | JOURNAL OF SOUTHWEST UNIVERSITY | https://cnki.net/ |
| Liu, S., et al. | 2013 | [65] | Journal of Sichuan Agricultural University | https://cnki.net/ |
| Guan, X., et al. | 2017 | [92] | Environmental Earth Sciences | https://doi.org/10.3390/rs9101082 |
| Zhang, R., et al. | 2017 | [93] | Remote Sensing | https://doi.org/10.3390/rs9050477 |
| Gu, X., et al. | 2007 | [80] | Journal of natural resources | https://cnki.net/ |
| Zhang, X. | 2011 | [94] | Remote Sensing | https://cnki.net/ |
| Zhao, X. | 2019 | [95] | China Academy of Forestry Sciences | https://cnki.net/ |
| Liu, X. | 2019 | [96] | Southwest University | https://doi.org/10.27284/d.cnki.gsxiu.2019.000161 |
| Yu, W., D. Dong, and J. Ni | 2010 | [97] | Shanxi University | https://doi.org/10.19687/j.cnki.1673-7105.2010.02.004 |
| Ni, J., Z. Xin-Shi, and M.O.S. Jonathan | 2001 | [98] | Journal of subtropical resources and environment | www.edpsciences.org |
| Liu, L.B., et al. | 2016 | [90] | Forests | https://doi.org/10.3390/f7080173 |
References
- Fang, J.; Yu, G.; Liu, L.; Hu, S.; Chapin, F. Climate change, human impacts, and carbon sequestration in China. Proc. Natl. Acad. Sci. USA 2018, 115, 4015–4020. [Google Scholar] [CrossRef]
- Wang, B.; Niu, X.; Song, Q. Analysis of forest carbon neutralization capacity in China based on full caliber carbon sink monitoring. Environ. Prot. 2021, 49, 30–34. [Google Scholar]
- Rafique, R.; Zhao, F.; Jong, R.; Zeng, N.; Asrar, G. Global and Regional Variability and Change in Terrestrial Ecosystems Net Primary Production and NDVI: A Model-Data Comparison. Remote Sens. 2016, 8, 117. [Google Scholar] [CrossRef]
- Ji, Y.; Zhou, G.; Luo, T.; Dan, Y.; Zhou, L.; Lv, X. Variation of net primary productivity and its drivers in China’s forests during 2000–2018. For. Ecosyst. 2020, 7, 15. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, K.; Zeng, Z.; Zou, Z.; Xu, Y.; Zeng, F. Multiple Factors Drive Variation of Forest Root Biomass in Southwestern China. Forests 2018, 9, 456. [Google Scholar] [CrossRef]
- Piao, S.; Fang, J.; Ciais, P.; Peylin, P.; Huang, Y.; Sitch, S.; Wang, T. The carbon balance of terrestrial ecosystems in China. Nature 2009, 458, 1009–1013. [Google Scholar] [CrossRef]
- Tang, X.; Zhao, X.; Bai, Y.; Tang, Z.; Wang, W.; Zhao, Y.; Wan, H.; Xie, Z.; Shi, X.; Wu, B.; et al. Carbon pools in China’s terrestrial ecosystems: New estimates based on an intensive field survey. Proc. Natl. Acad. Sci. USA 2018, 115, 4021–4026. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Wang, G.; Liu, G.; Xu, S. Forest biomass of China: An estimate based on the biomass–volume relationship. Ecol. Appl. 1998, 8, 1084–1091. [Google Scholar]
- Xu, E.; Nie, Y.; Bing, X. Analysis on the forest land use efficiency change based on forest resource inventory data. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2022, 1–13. Available online: http://kns.cnki.net/kcms/detail/32.1161.S.20220620.1359.002.html (accessed on 16 August 2022).
- Zeng, S.; Liu, Z.; Kaufmann, G. Sensitivity of the global carbonate weathering carbon-sink flux to climate and land-use changes. Nat. Commun. 2019, 10, 5749. [Google Scholar] [CrossRef]
- Hong, C.; Jing, X.; Cheng, C.; Wang, S.; Yang, X.; Xiang, X. A review of land net primary productivity estimation models based on integrated remote sensing data. Prog. Geogr. Sci. 2017, 36, 924–939. [Google Scholar]
- Li, M.; Wang, b.; Fan, w.; Zhao, D. Simulation study on net primary productivity of northeast forest region and forest fire disturbance in Daxinganling region. J. Plant Ecol. 2015, 39, 322–332. [Google Scholar]
- Li, Z. Study on Maize Growth Monitoring Method Based on Light Energy Utilization Model and Quantitative Remote Sensing. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2014. [Google Scholar]
- Fang, J.; Guo, Z.; Piao, S.; Chen, A. Terrestrial vegetation carbon sinks in China,1981–2000. Sci. China (Ser. D Earth Sci.) 2007, 50, 1341–1350. [Google Scholar] [CrossRef]
- Dennis, D.B. How eddy covariance flux measurements have contributed to our understanding of Global Change Biology. Glob. Chang. Biol. 2019, 26, 242–260. [Google Scholar]
- Marc, A. Eddy covariance CO2 flux measurements in nocturnal conditions: An analysis of the problem. Ecol. Appl. 2008, 18, 1368–1378. [Google Scholar]
- Piao, S.; He, Y.; Wang, X.; Chen, F. Carbon sequestration estimation of terrestrial ecosystems in China: Methods, progress and Prospects. Chin. Sci. Earth Sci. 2022, 52, 1010–1020. [Google Scholar]
- Lieth, H. Modeling the primary productivity of the world. In Primary Productivity of the Biosphere; Springer: Berlin/Heidelberg, Germany, 1975; pp. 237–263. [Google Scholar]
- Lieth, H. Evapotranspiration and primary productivity: CW Thornthwaite memorial model. Publ. Climatol. 1972, 25, 37–46. [Google Scholar]
- Zenbei, U.; Hiroshi, S. Agroclimatic Evaluation of Net Primary Productivity of Natural Vegetations. J. Agric. Meteorol. 1985, 40, 343–352. [Google Scholar]
- Zhou, G.; Zhang, X. A preliminary study on the net primary productivity model of natural vegetation. J. Plant Ecol. 1995, 19, 193–200. [Google Scholar]
- Adams, B.; White, A.; Lenton, T.M. An analysis of some diverse approaches to modelling terrestrial net primary productivity. Ecol. Model. 2004, 177, 353–391. [Google Scholar] [CrossRef]
- Sun, J.; Guan, D.; Wu, J.; Jin, C.; Yuan, F. Research advances in net primary productivity of terrestrial vegetation. World For. Res. 2012, 25, 1–6. [Google Scholar]
- Shvidenko, A.; Schepaschenko, D.; Nilsson, S.; Bouloui, Y. Semi-empirical models for assessing biological productivity of Northern Eurasian forests. Ecol. Model. 2007, 204, 163–179. [Google Scholar] [CrossRef]
- Zaks, D.; Ramankutty, N.; Barford, C.; Foley, J. From Miami to Madison: Investigating the relationship between climate and terrestrial net primary production. Glob. Biogeochem. Cycles 2007, 21, 3. [Google Scholar] [CrossRef]
- Wang, Z.; Guo, Z.; Song, K.; Liu, D.; Zhang, B.; Zhang, S.; Li, F. Effects of land use/Cover Change on net primary productivity of vegetation in Sanjiang Plain from 2000 to 2005. J. Nat. Resour. 2009, 24, 136–146. [Google Scholar]
- Zhu, W.; Chen, Y.; Xu, D.; Dong, J. Advances in the calculation models of net primary productivity of terrestrial vegetation. J. Ecol. 2005, 24, 296–300. [Google Scholar]
- Zhang, M.; Jiang, W.; Chen, Q.; Zhao, Y.; Liu, X.; Kicklighter, D.; Grace, A.; Moore, B.; Vorosmarty, C. Research Progress on estimation model of grassland net primary productivity. Grassl. J. 2011, 19, 356–366. [Google Scholar]
- McGuire, A.; Melillo, J.; Joyce, L.; Kicklighter, D.; Grace, A.; Moore, B.; Vorosmarty, C. Interactions between carbon and nitrogen dynamics in estimating net primary productivity for potential vegetation in North America. Glob. Biogeochem. Cycles 1992, 6, 102–124. [Google Scholar] [CrossRef]
- Running, S.W.; Gower, S.T. FOREST-BGC, A general model of forest ecosystem processes for regional applications. II. Dynamic carbon allocation and nitrogen budgets. Tree Physiol. 1991, 9, 147–160. [Google Scholar] [CrossRef]
- Vemap, P. Vegetation/ecosystem modeling and analysis project: Comparing biogeography and biogeochemistry models in a continental-scale study of terrestrial ecosystem responses to climate change and CO2 doubling. Glob. Biogeochem. Cycles 1995, 9, 407. [Google Scholar]
- Steven, W.R.; Joseph, C.C. A general model of forest ecosystem processes for regional applications I. Hydrologic balance, canopy gas exchange and primary production processes. Ecol. Model. 1988, 42, 125–154. [Google Scholar]
- Monteith, J.L. Solar Radiation and Productivity in Tropical Ecosystems. J. Appl. Ecol. 1972, 9, 747–766. [Google Scholar] [CrossRef]
- Nikolaos, M.F.; Lisa Patrick, B.; Alexander, S.; Gregory, P.; Owen, K.; Sandra, D.; Brian, J.; William, F.; Emanuel, G.; Rossella, G.; et al. Solar radiation and functional traits explain the decline of forest primary productivity along a tropical elevation gradient. Ecol. Lett. 2017, 20, 730–740. [Google Scholar]
- Prince, S.D.; Goward, S.N. Global primary production: A remote sensing approach. J. Biogeogr. 1995, 22, 815–835. [Google Scholar] [CrossRef]
- Veroustraete, F.; Sabbe, H.; Eerens, H. Estimation of carbon mass fluxes over Europe using the C-Fix model and Euroflux data. Remote Sens. Environ. 2002, 83, 376–399. [Google Scholar] [CrossRef]
- Box, E.O.; Holben, B.N.; Kalb, V. Accuracy of the AVHRR vegetation index as a predictor of biomass, primary productivity and net CO2 flux. Vegetatio 1989, 80, 71–89. [Google Scholar] [CrossRef]
- Yuan, W.; Cai, W.; Liu, D.; Dong, W. Satellite-based vegetation production models of terrestrial ecosystem: An overview. Adv. Earth Sci. 2014, 29, 541–550. [Google Scholar]
- Shen, B. The Simulation and Analysis of NPP Based on CASA Model in Hulunber Grassland. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2019. [Google Scholar]
- Potter, C.S.; Randerson, J.; Field, C.; Matson, P.; Vitousek, P.; Mooney, H.; Klooster, S. Terrestrial ecosystem production: A process model based on global satellite and surface data. Glob. Biogeochem. Cycles 1993, 7, 811–841. [Google Scholar] [CrossRef]
- Li, C.; Fan, Y.; Cao, H.; Han, H. Quantitative study on human impact of primary productivity of Vegetation Based on CASA model—A case study of Shiyang River Basin. Geogr. Arid. Areas 2018, 41, 142–151. [Google Scholar]
- Liu, Z.; Hu, M.; Hu, Y.; Wang, G. Estimation of net primary productivity of forests by modified CASA models and remotely sensed data. Int. J. Remote Sens. 2017, 39, 1092–1116. [Google Scholar] [CrossRef]
- Zhu, W. Remote Sensing Estimation of Net Primary Productivity of Terrestrial Ecosystems in China and its Relationship with Climate Change. Ph.D. Thesis, Beijing Normal University, Beijing, China, 2005. [Google Scholar]
- Piao, S.; Fang, J.; Guo, Q. Estimation of net primary productivity of vegetation in China using CASA model. J. Plant Ecol. 2001, 25, 603–608. [Google Scholar]
- Dong, D.; Ni, J. Simulation of net primary productivity of Karst Vegetation in Southwest China using CASA model. J. Ecol. 2011, 31, 1855–1866. [Google Scholar]
- Li, C.; Li, C.; Fan, Y.; Cao, H.; Han, H.; Sun, H.; Wang, Y. NPP remote sensing estimation and analysis of CASA model based on Correction—Taking Hexi Corridor as an example. J. Ecol. 2019, 39, 1616–1626. [Google Scholar]
- Yan, Y.; Liu, X.; Ou, J.; Li, X.; Wen, Y. Assimilating multi-source remotely sensed data into a light use efficiency model for net primary productivity estimation. Int. J. Appl. Earth Obs. Geoinf. 2018, 72, 11–25. [Google Scholar] [CrossRef]
- Lee, J.-E.; Frankenberg, C.; van der Tol, C.; Berry, J.; Guanter, L.; Boyce, C.; Fisher, J.; Morrow, E.; Worden, J.; Asefi, S. Forest productivity and water stress in Amazonia: Observations from GOSAT chlorophyll fluorescence. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130171. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Heinsch, F.; Nemani, R.; Running, S. Improvements of the MODIS terrestrial gross and net primary production global data set. Remote Sens. Environ. 2005, 95, 164–176. [Google Scholar] [CrossRef]
- Jenkins, J.; Richardson, A.; Braswell, B.; Ollinger, S.; Hollinger, D.; Smith, M. Refining light-use efficiency calculations for a deciduous forest canopy using simultaneous tower-based carbon flux and radiometric measurements. Agric. For. Meteorol. 2007, 143, 64–79. [Google Scholar] [CrossRef]
- Gustau, C.-V.; Manuel, C.; Álvaro, M.; Sophia, W.; Grégory, D.; Alessandro, C.; Miguel, D.; Jordi, M.; Francisco Javier, G.; Luis, G.; et al. A unified vegetation index for quantifying the terrestrial biosphere. Sci. Adv. 2021, 7, eabc7447. [Google Scholar]
- César, H.-H.; Goulden, M.L. Plant Traits Help Explain the Tight Relationship between Vegetation Indices and Gross Primary Production. Remote Sens. 2020, 12, 1405. [Google Scholar]
- Sun, C.; Chen, Y.; Wu, W.; Chen, W.; Han, Z.; Li, J. Study on spatial distribution pattern of grassland NPP in southern China based on climate productivity model. J. Yangzhou Univ. 2013, 34, 56–61. [Google Scholar]
- Sun, J.; Niu, H.; Yuan, M. Analysis on NPP spatial pattern change of terrestrial vegetation ecosystem in China. J. Agric. Mach. 2020, 51, 162–168. [Google Scholar]
- Sun, Z.; Xie, S. Spatial and temporal evolution and factor detection of net primary productivity in Yunnan Province Based on geographical detectors. J. Ecol. 2021, 40, 3836–3848. [Google Scholar] [CrossRef]
- Wu, G. Estimation of Net Primary Productivity and Carbon Storage of Forest Vegetation in Jiangxi Province. Ph.D. Thesis, Nanjing Forestry University, Nanjing, China, 2015. [Google Scholar]
- Zhao, M.; Liu, Y.; Yang, J.; Liu, Y.; Zhao, N.; Yue, T. Spatial and temporal variation of NPP in Chinese Vegetation Based on HASM and its relationship with climate. J. Ecol. Environ. 2019, 28, 215–225. [Google Scholar]
- Li, M.; Yao, W.; Ren, X.; Zhang, L.; Xu, W.; Gu, F.; He, H. Estimation of net primary productivity of forest ecosystem in Shennongjia forest region from 1981 to 2015. Environ. Sci. Res. 2019, 32, 749–757. [Google Scholar]
- Hong, X.; Huang, Y.; Sun, T. Spatial and temporal evolution of net primary productivity of vegetation in karst areas of Southwest China from 2001 to 2018. J. Ecol. 2021, 41, 9836–9846. [Google Scholar]
- Lin, X.; Lin, X.; Cai, X.; Wang, J.; Mei, Z.; Dai, L. Spatial and temporal changes of net primary productivity of vegetation in Guizhou Province from 2000 to 2010. Environ. Prot. Sci. 2015, 41, 91–95+136. [Google Scholar] [CrossRef]
- Xiong, X.; Liao, C.; Hu, B.; Chen, Y.; Zhu, H. Spatial and temporal differentiation and driving forces of NPP in Guangxi vegetation based on MODIS. J. Guangxi Norm. Univ. 2019, 37, 187–195. [Google Scholar]
- Xu, Y.; Zhang, Y.; Pan, W. Dynamic changes of spatial and temporal pattern of net primary productivity (NPP) of vegetation in Yunnan Guizhou Plateau. Environ. Sustain. Dev. 2018, 43, 96–99. [Google Scholar]
- Zhao, Z.; Wu, X.; Li, G.; Li, J. Drought degree and its impact on net primary productivity of vegetation in Southwest China from 2009 to 2011. J. Ecol. 2015, 35, 350–360. [Google Scholar]
- Zhang, R.; Luo, H.; Zhang, R.; Liu, G.; Zheng, H.; Yang, T. Estimation of net primary productivity of vegetation and evaluation of its ecological service value in Chongqing. J. Southwest Univ. 2015, 37, 40–46. [Google Scholar]
- Liu, S.; Lu, T.; Tang, B.; Wu, Y.; Wu, N. Analysis of net primary productivity and its spatial-temporal pattern of vegetation in Sichuan Based on CASA model. J. Sichuan Agric. Univ. 2013, 31, 269–276+282. [Google Scholar]
- Ge, W.; Deng, L.; Wang, F.; Han, J. Quantifying the contributions of human activities and climate change to vegetation net primary productivity dynamics in China from 2001 to 2016. Sci. Total Environ. 2021, 773, 145648. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.; Wang, L.; Zhao, K. The causes of rocky desertification and Its Ecological Restoration Countermeasures in karst mountainous areas of Southwest China. J. West China Norm. Univ. 2004, 04, 440–443. [Google Scholar]
- Xu, Y.; Xiao, F.; Yu, L. Spatial and temporal distribution of net primary productivity of forest ecosystems in China and its response to climate change. J. Ecol. 2020, 40, 4710–4723. [Google Scholar]
- Yang, J.; Zhang, X.; Luo, Z.; Yu, X. Nonlinear Variations of Net Primary Productivity and Its Relationship with Climate and Vegetation Phenology, China. Forests 2017, 8, 361. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.; Allen, A.; Savage, V.; West, G. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Šímová, I.; Storch, D. The enigma of terrestrial primary productivity: Measurements, models, scales and the diversity–productivity relationship. Ecography 2017, 40, 239–252. [Google Scholar] [CrossRef]
- Zhou, J.; Ma, M.; Xiao, Q.; Wen, J. Vegetation cover dynamics and its relationship with climatic factors in Southwest China. Remote Sens. Technol. Appl. 2017, 32, 966–972. [Google Scholar]
- Yin, L.; Dai, E.; Zheng, D.; Wang, Y.; Ma, L.; Tong, M. What drives the vegetation dynamics in the Hengduan Mountain region, southwest China: Climate change or human activity? Ecol. Indic. 2020, 112, 106013. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, M.; Lin, Z.; Xu, X. Spatial heterogeneity of the relationship between vegetation dynamics and climate change and their driving forces at multiple time scales in Southwest China. Agric. For. Meteorol. 2018, 256, 10–21. [Google Scholar] [CrossRef]
- Li, Q. Study on Spatial and Temporal Changes and Driving Forces of Net Primary Productivity of Vegetation in Panxi Region. Master’s Thesis, Chengdu University of Technology, Chengdu, China, 2019. [Google Scholar]
- Luo, Y. Study on the Influence of Regional Climate on Net Primary Productivity of Vegetation in Guizhou, in Lanzhou University. Master’s Thesis, Lanzhou University, Lanzhou, China, 2009. [Google Scholar]
- Liang, W.; Yang, Y.; Fan, D.; Guan, H.; Zhang, T.; Long, D.; Zhou, Y.; Bai, D. Analysis of spatial and temporal patterns of net primary production and their climate controls in China from 1982 to 2010. Agric. For. Meteorol. 2015, 204, 22–36. [Google Scholar] [CrossRef]
- Liu, S.; Guo, Q.; Wang, B. Prediction of the response of forest productivity to climate change in China. J. Ecol. 1998, 05, 32–37. [Google Scholar]
- Fei, X. Carbon exchange in typical forest ecosystems in Yunnan and its response to climate change. In Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences; University of Chinese Academy of Sciences: Beijing, China, 2018; p. 220. [Google Scholar]
- Gu, X.; Huang, W.; Ji, J.; Wu, Z. Effects of climate change on net primary productivity of vegetation in Southwest China in recent 20 years. J. Nat. Resour. 2007, 7, 251–259+324. [Google Scholar]
- Li, X.; Li, Y.; Chen, A.; Gao, M.; Slette, I.; Piao, S. The impact of the 2009/2010 drought on vegetation growth and terrestrial carbon balance in Southwest China. Agric. For. Meteorol. 2019, 269, 239–248. [Google Scholar] [CrossRef]
- Yuan, W.; Zheng, Y.; Piao, S.; Ciais, P.; Lombardozzi, D.; Wang, Y.; Ryu, Y.; Chen, G.; Dong, W.; Hu, Z.; et al. Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci. Adv. 2019, 5, eaax1396. [Google Scholar] [CrossRef] [PubMed]
- Madeline, R.; Jordan, G.J.; Brodribb, T.J. Acclimation to humidity modifies the link between leaf size and the density of veins and stomata. Plant Cell Env. 2014, 37, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Shamshiri, R.R.; Jones, J.; Thorp, K.; Ahmad, D.; Man, H.; Taheri, S. Review of optimum temperature, humidity, and vapour pressure deficit for microclimate evaluation and control in greenhouse cultivation of tomato: A review. Int. Agrophys. 2018, 32, 287–302. [Google Scholar] [CrossRef]
- Lopez, J.; Way, D.A.; Sadok, W. Systemic effects of rising atmospheric vapor pressure deficit on plant physiology and productivity. Glob. Chang. Biol. 2021, 27, 1704–1720. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Wang, S.; Yuan, G.; Yang, Y.; Cao, M. Evapotranspiration of a tropical rain forest in Xishuangbanna, southwest China. Hydrol. Processes 2010, 24, 2405–2416. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, Y.; Wu, Q.; Liu, S.; Song, C.; Xiao, J.; Band, L.; Vose, J. Vegetation structural change and CO2 fertilization more than offset gross primary production decline caused by reduced solar radiation in China. Agric. For. Meteorol. 2021, 296, 108207. [Google Scholar] [CrossRef]
- Kubiske, M.E.; Pregitzer, K.; Mikan, C.; Zak, D.; Maziasz, J.; Teeri, A. Populus tremuloides photosynthesis and crown architecture in response to elevated CO2 and soil N availability. Oecologia 1997, 110, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wang, S.; Lai, P.; He, B.; Ma, M.; Han, X. Remote sensing monitoring and spatial-temporal analysis of forest loss characteristics in Southwest China from 2001 to 2019. Remote Sens. Technol. Appl. 2021, 36, 552–563. [Google Scholar]
- Liu, L.; Yang, H.M.; Xu, Y.; Guo, Y.M.; Ni, J. Forest Biomass and Net Primary Productivity in Southwestern China: A Meta-Analysis Focusing on Environmental Driving Factors. Forests 2016, 7, 173. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, W.; Yang, T.; Zhao, G.; Wang, S. Change characteristics of net primary productivity of different vegetation types in China. J. Fudan Univ. 2012, 51, 377–381. [Google Scholar]
- Guan, X.; Shen, H.; Gan, W.; Yang, G.; Wang, L.; Li, X.; Zhang, L. A 33-year NPP monitoring study in southwest China by the fusion of multi-source remote sensing and station data. Remote Sens. 2017, 9, 1082. [Google Scholar] [CrossRef]
- Zhang, R.; Zhou, Y.; Luo, H.; Wang, F.; Wang, S. Estimation and analysis of spatiotemporal dynamics of the net primary productivity integrating efficiency model with process model in karst area. Remote Sens. 2017, 9, 477. [Google Scholar] [CrossRef]
- Zhang, X. Spatiotemporal Dynamics of NDVI and NPP and Their Correlation with Climatic Factors in Southwest China. Master’s Thesis, China Academy of Forestry Sciences, Beijing, China, 2011. [Google Scholar]
- Zhao, X. Spatial and Temporal Differentiation of Net Primary Productivity of Vegetation in Qinba Mountain Area from 2000 to 2015. Master’s Thesis, Southwest University, Chongqing, China, 2019. [Google Scholar]
- Liu, X. Productivity of Main Terrestrial Vegetation Types in China and Its Relationship with Climatic Factors. Master’s Thesis, Shanxi University, Taiyuan, China, 2019. [Google Scholar]
- Yu, W.; Dong, D.; Ni, J. Comparison of biomass and productivity between karst and non karst forests in Southwest China. J. Subtrop. Resour. Environ. 2010, 5, 25–30. [Google Scholar]
- Ni, J.; Zhang, X.; Scurlock, J. Synthesis and analysis of biomass and net primary productivity in Chinese forests. Ann. For. Sci. 2001, 58, 351–384. [Google Scholar] [CrossRef]
- Luo, T. Patterns of biological production and its mathematical models for main forest types of China. Ph.D. Dissertation, Committee of Synthesis Investigation of Natural Resources, Chinese Academy of Sciences, Beijing, China, 1996. [Google Scholar]
- Cai, W.; Yuan, W.; Liang, S.; Zhang, X.; Dong, W.; Xia, J.; Fu, Y.; Chen, Y.; Liu, D.; Zhang, Q. Improved estimations of gross primary production using satellite-derived photosynthetically active radiation. J. Geophys. Res. Biogeosci. 2014, 119, 110–123. [Google Scholar] [CrossRef]
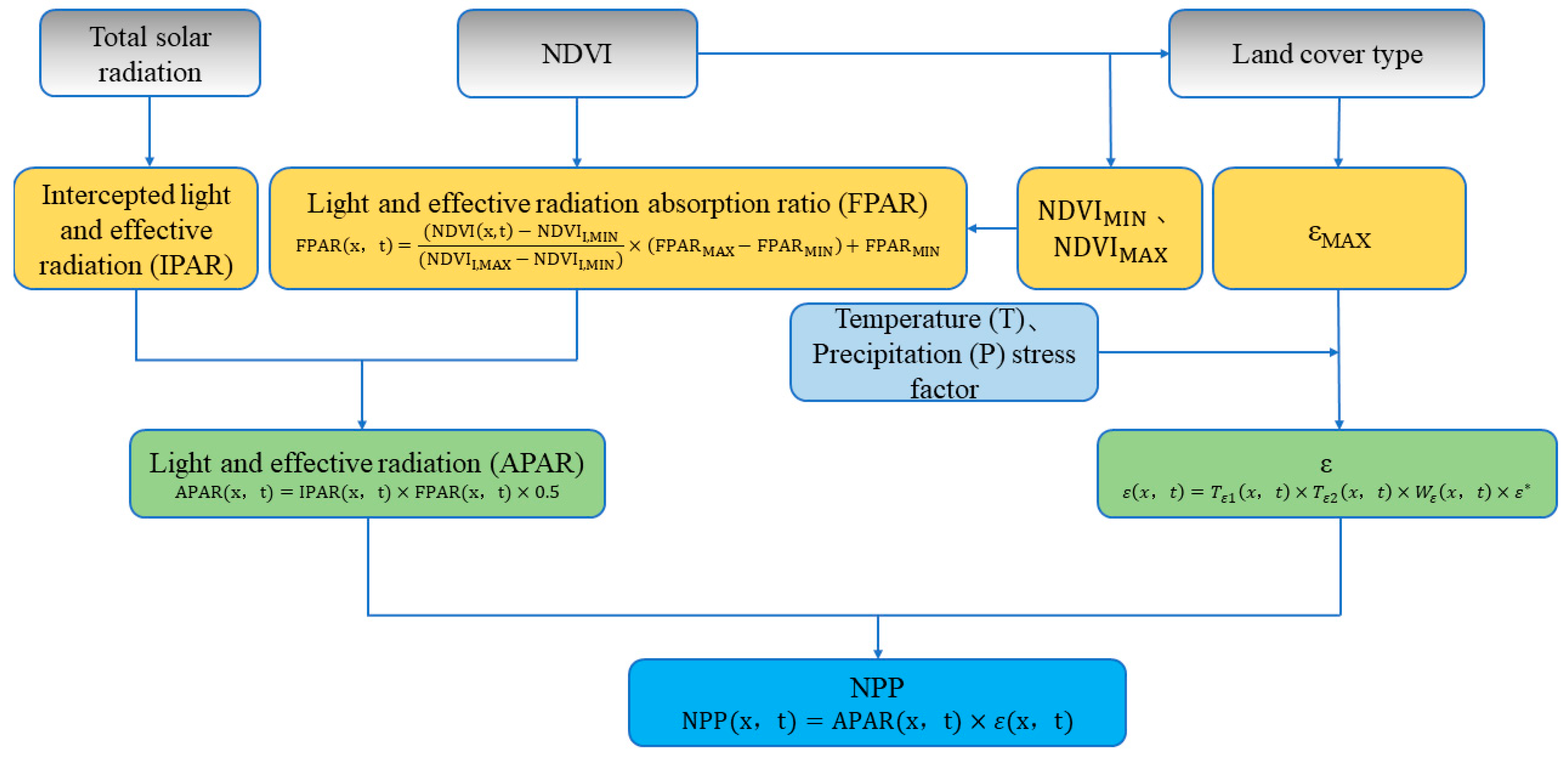
| Model | Expression | Parameters | Reference |
|---|---|---|---|
| Miami model | t: Temperature, r: Precipitation | [18] | |
| Thornthwaite Memorial model | E: Actual evapotranspiration, L; Maximum evapotranspiration | [19] | |
| Chikugo model | RDI: Radiation dryness, Rn: Net radiation, Lr: Latent heat of evaporation | [20] | |
| Integrated model | PER: Potential evapotranspiration rate; BT: Biological temperature | [21] |
| Model | CASA Model | GLO-PEM Model | C-FIX Model |
|---|---|---|---|
| Expression | |||
| Parameter | APAR: Photosynthetic active radiation, ε: Actual light energy utilization | : Photosynthetic active radiation, : Photosynthetic active radiation ratio, : Autotrophic respiration | : Autotrophic respiration rate of vegetation |
| Advantage |
| Strong real-time performance |
|
| Shortcoming | Difficult to determine parameters | Limited data source | The calculation of autotrophic respiration rate is too simplified |
| Common ground | Based on the concept of plant photosynthesis process and light energy utilization rate, solar radiation is taken as input parameter to obtain or estimate APAR and then get the value of ε and generate NPP | ||
| Vegetation Type | PAR | ε | NPP |
|---|---|---|---|
| Broad-leaf evergreen forests | 1867 | 0.265 | 525 |
| Broad-leaf deciduous forests | 1108 | 0.202 | 304 |
| Needle-leaf and broad-leaf mixed forests | 1346 | 0.137 | 330 |
| Needle-leaf evergreen forests | 1253 | 0.243 | 354 |
| Needle-leaf deciduous forests | 1585 | 0.133 | 432 |
| Broad-leaf trees with groundcover | 927 | 0.217 | 283 |
| Model | Advantage | Shortcoming | Representative | Study Time | Study Area | NPP gC/m2·a | Reference |
|---|---|---|---|---|---|---|---|
| Statistical model | Simple, fewer parameters required |
| Miami | 1980–2010 | Grassland in southern China | 1418.83 | [53] |
| Thornthwaite | 2000–2015 | China | 899.40 | [54] | |||
| Process model |
|
| Biome-BGC | 2000–2015 | Yunnan Province | 892.83 | [55] |
| BEPS | 2001–2010 | Jiangxi Province | 692.96 | [56] | |||
| LPJ-GUESS | 2001–2015 | Southwest China | 531.81 | [57] | |||
| CEVSA | 1981–2015 | Shennongjia Forestry District | 628.27 | [58] | |||
| Light energy utilization model |
|
| MODIS | 2001–2018 | Southwest China | 710.44 | [59] |
| CASA | 2000–2010 | Guizhou Province | 828.1 | [60] | |||
| MODIS | 2000–2015 | Guangxi Province | 724.5 | [61] | |||
| MODIS | 2000–2014 | Yunnan Guizhou Plateau | 768 | [62] | |||
| GLO-PEM | 2001–2011 | Southwest China | 540.33 | [63] | |||
| CASA | 2002–2011 | Chongqing Province | 500.45 | [64] | |||
| CASA | 2000–2011 | Sichuan Province | 303.27 | [65] |
| Region | Year | NPP(gC/m2·a) | Reference | ||||
|---|---|---|---|---|---|---|---|
| Broad-Leaf Evergreen Forests | Broad-Leaf Deciduous Forests | Needle-Leaf Evergreen Forests | Needle-Leaf Deciduous Forests | Needle-Leaf and Broad-Leaf Mixed Forests | |||
| Yunnan Province | 1982–2014 | 996 | 778 | / | 536 | / | [92] |
| Chongqing Province | 2001–2011 | 830.4 | 726.74 | 663.51 | 525.81 | / | [93] |
| Yunnan Province | 1981–2000 | 1000 | / | / | / | 400~800 | [80] |
| Southwest China | 1982–2006 | 925.29 | / | 911.01 | / | 976.45 | [94] |
| Qinba Mountain | 2000–2015 | 554.71 | 445.67 | 557.97 | 463.00 | 483.49 | [95] |
| China | 1982–2015 | 1323.71 | 637.21 | 497.59 | 442.35 | 832.06 | [96] |
| Region | Year | NPP gC/m2·a | Reference |
|---|---|---|---|
| Yunnan Guizhou Plateau | 2000–2014 | 875.69 | [62] |
| Guangxi Province | 1989–1993 | 1000.3 ± 5.97 | [97] |
| Guizhou Province | 1989–1993 | 1100.31 ± 6.39 | [97] |
| Sichuan Province | 1989–1993 | 600.35 ± 3.40 | [97] |
| Yunnan Province | 1989–1993 | 700.75 ± 4.88 | [97] |
| China | 1982–2015 | 650.73 | [96] |
| Vegetation Type | NPP(t·ha−1year−1) | ||
|---|---|---|---|
| [97] | [98] | [90] | |
| Broad-leaf evergreen forests | 25.4 | 21.9 ± 5.3 | 17.6 ± 7.2 |
| Needle-leaf evergreen forests | 18.66 | 13.5 ± 3.0 | 12.1 ± 6.5 |
| Needle-leaf and broad-leaf mixed forests | / | 9.9 ± 5.0 | 9.0 ± 5.0 |
| Tropical rainforest | 16.85 | 27.1 ± 9.2 | 20.9 ± 6.7 |
| Needle-leaf deciduous and broad-leaf mixed forests | / | 15.2 ± 3.0 | 11.9 ± 2.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, B.; Liao, Z.; Chen, S.; Jia, H.; Zhu, J.; Fei, X. Net Primary Productivity of Forest Ecosystems in the Southwest Karst Region from the Perspective of Carbon Neutralization. Forests 2022, 13, 1367. https://doi.org/10.3390/f13091367
Zhou B, Liao Z, Chen S, Jia H, Zhu J, Fei X. Net Primary Productivity of Forest Ecosystems in the Southwest Karst Region from the Perspective of Carbon Neutralization. Forests. 2022; 13(9):1367. https://doi.org/10.3390/f13091367
Chicago/Turabian StyleZhou, Binghuang, Zhangze Liao, Sirui Chen, Hongyu Jia, Jingyu Zhu, and Xuehai Fei. 2022. "Net Primary Productivity of Forest Ecosystems in the Southwest Karst Region from the Perspective of Carbon Neutralization" Forests 13, no. 9: 1367. https://doi.org/10.3390/f13091367
APA StyleZhou, B., Liao, Z., Chen, S., Jia, H., Zhu, J., & Fei, X. (2022). Net Primary Productivity of Forest Ecosystems in the Southwest Karst Region from the Perspective of Carbon Neutralization. Forests, 13(9), 1367. https://doi.org/10.3390/f13091367