
Prediction of Native Seed Habitat Distribution According to SSP Scenario and Seed Transfer Zones: A Focus on Acer pictum subsp. mono and Quercus acuta


Abstract
:1. Introduction
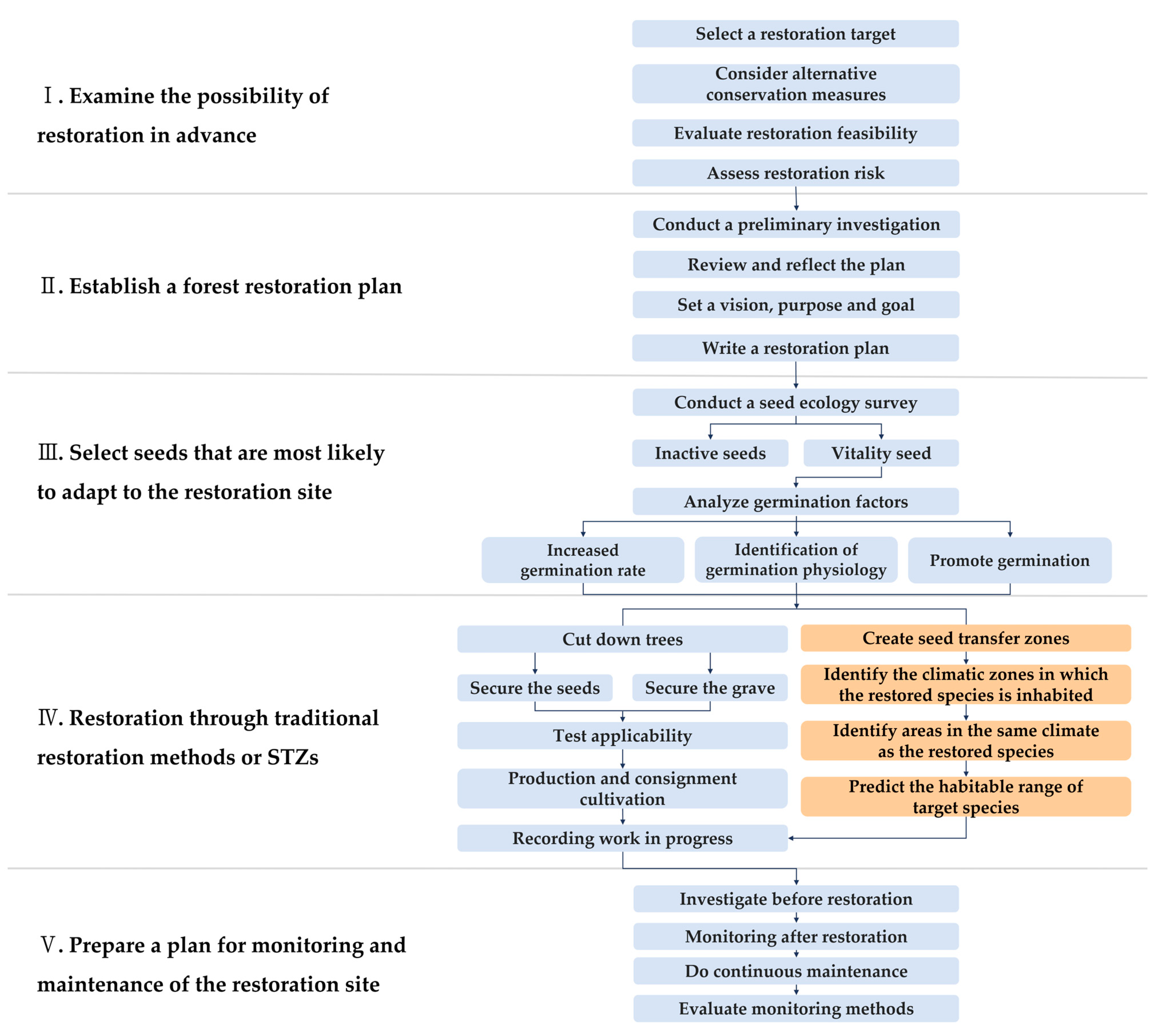
2. Materials and Methods
2.1. A. pictum and Q. acuta
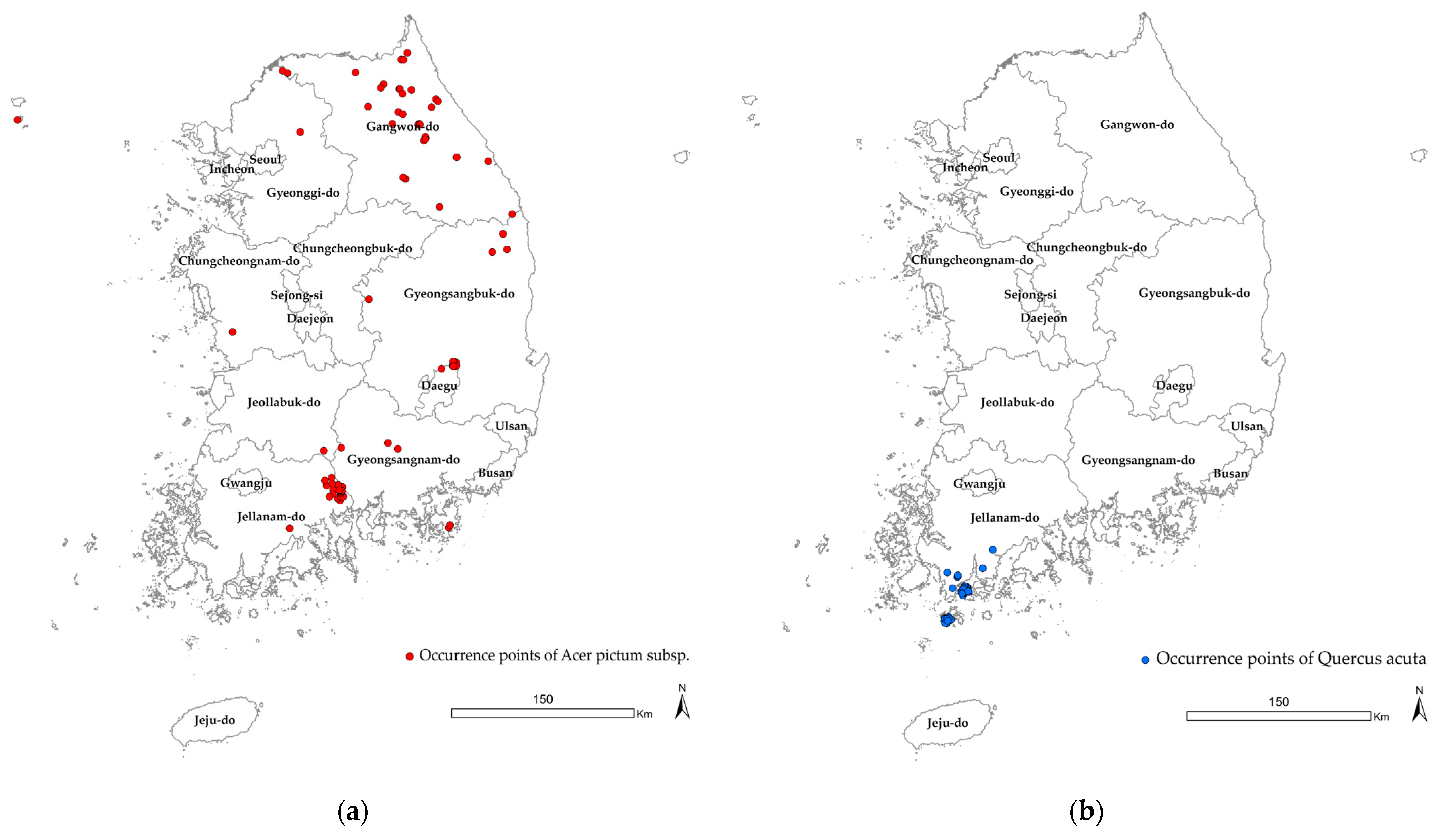
2.2. Species Occurrence Data
2.3. Environmental Data
2.4. South Korea Detailed Scenario
2.5. Construction of MaxEnt
2.6. Seed Transfer Zones
3. Results
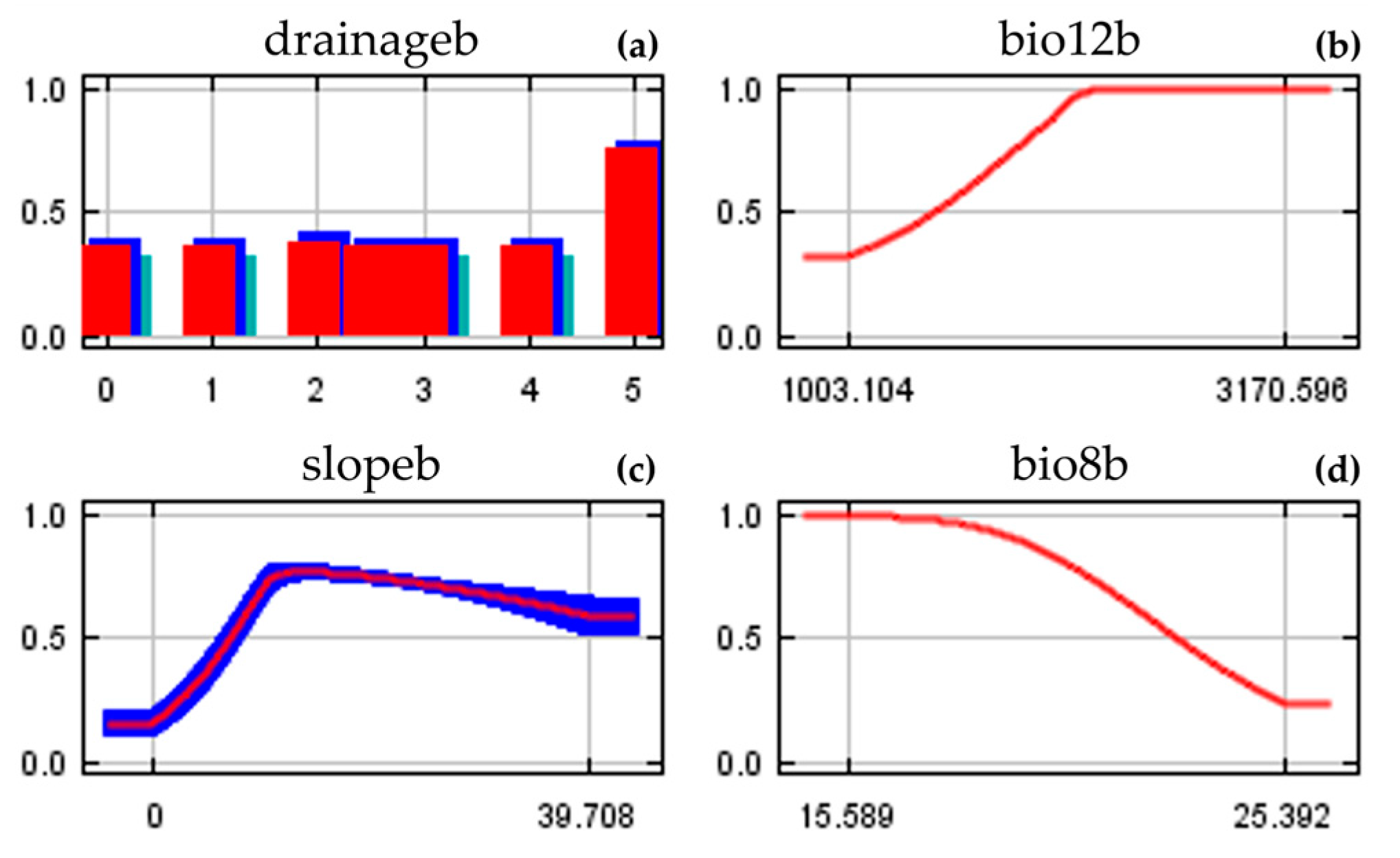
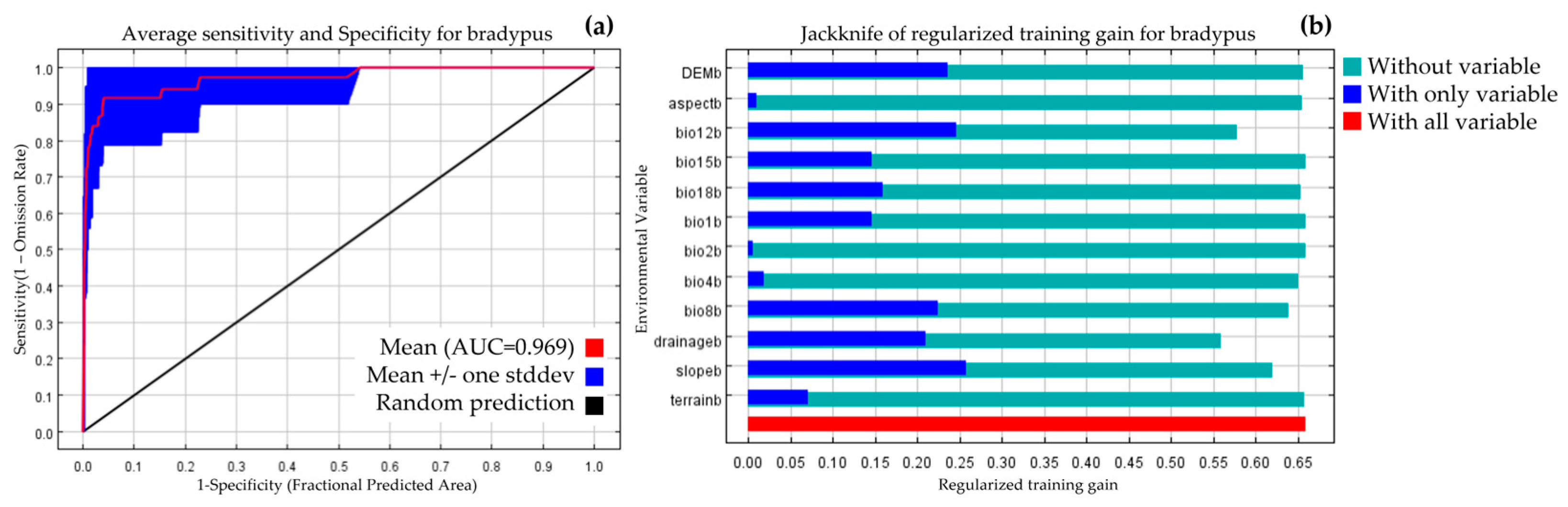
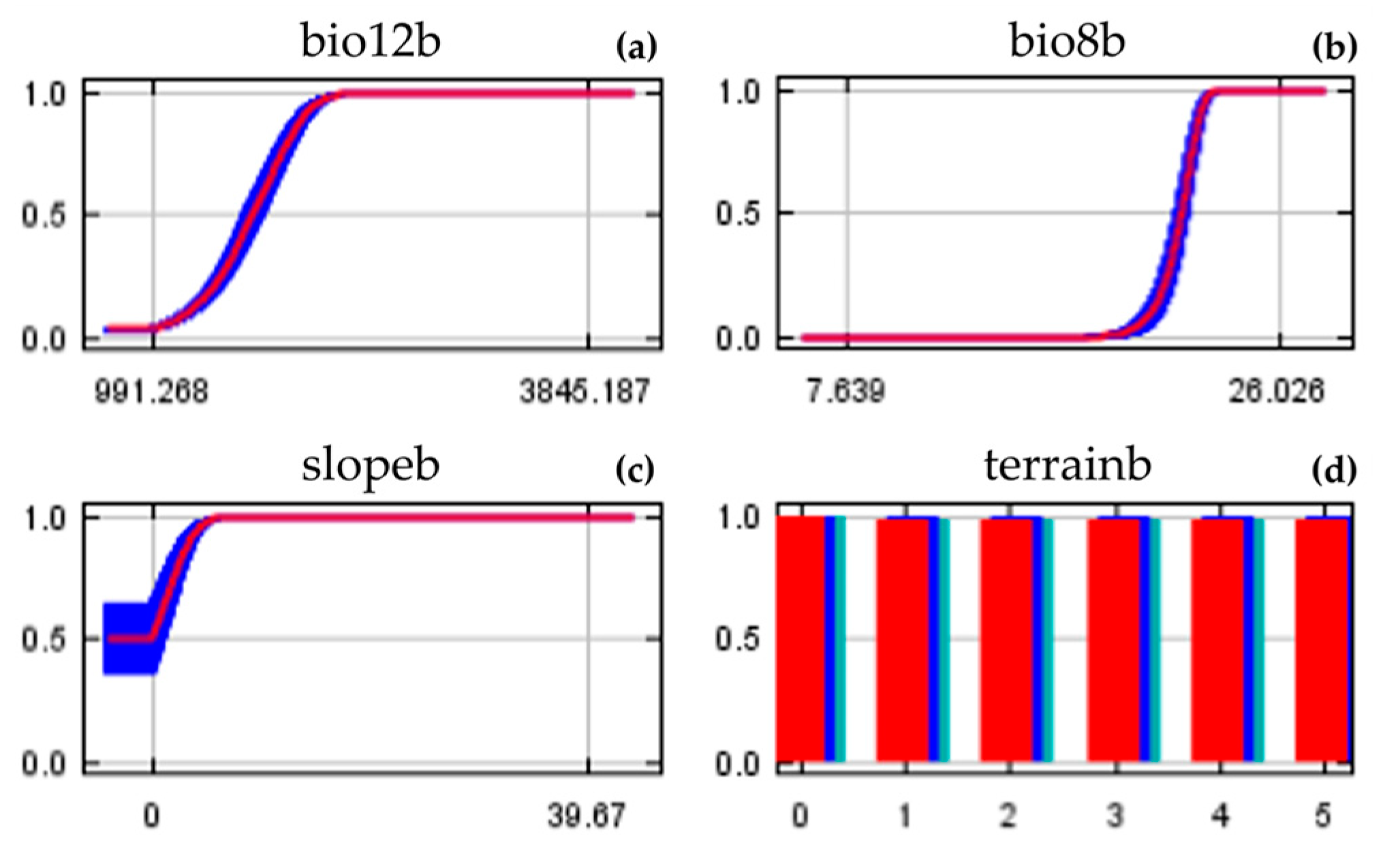
3.1. Model Performance and Variables Importance
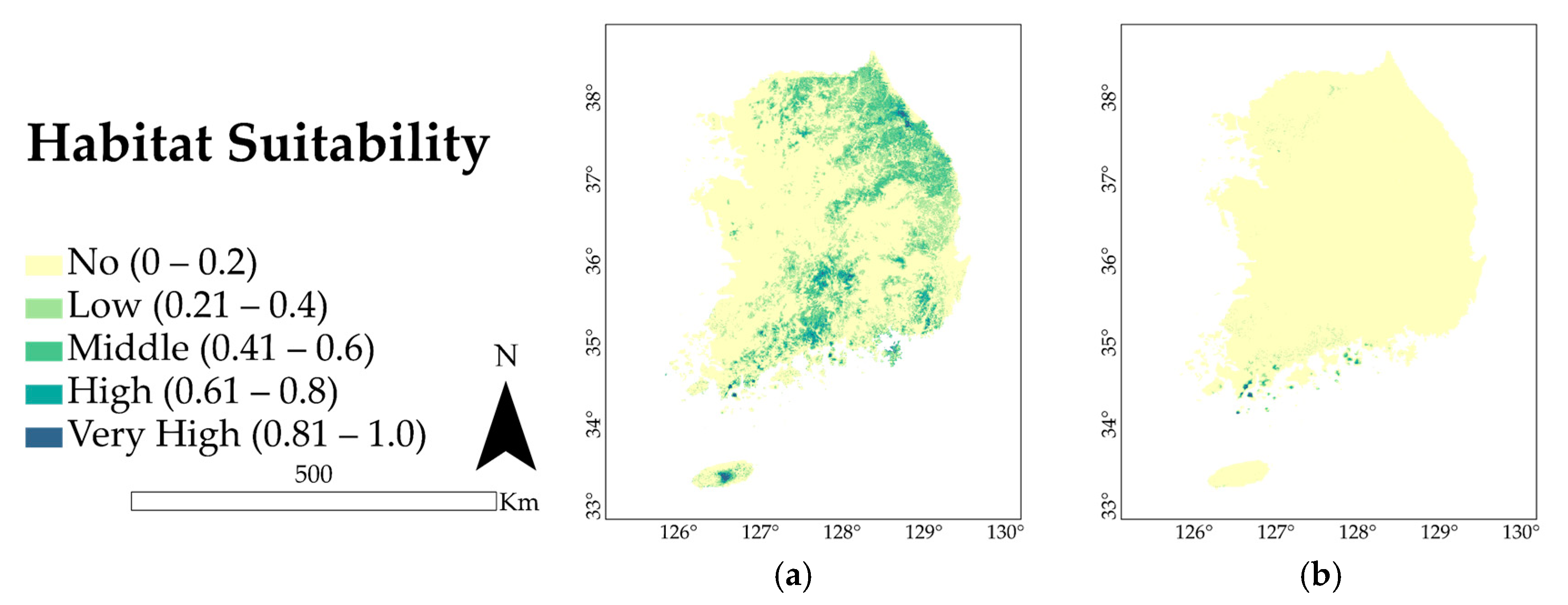
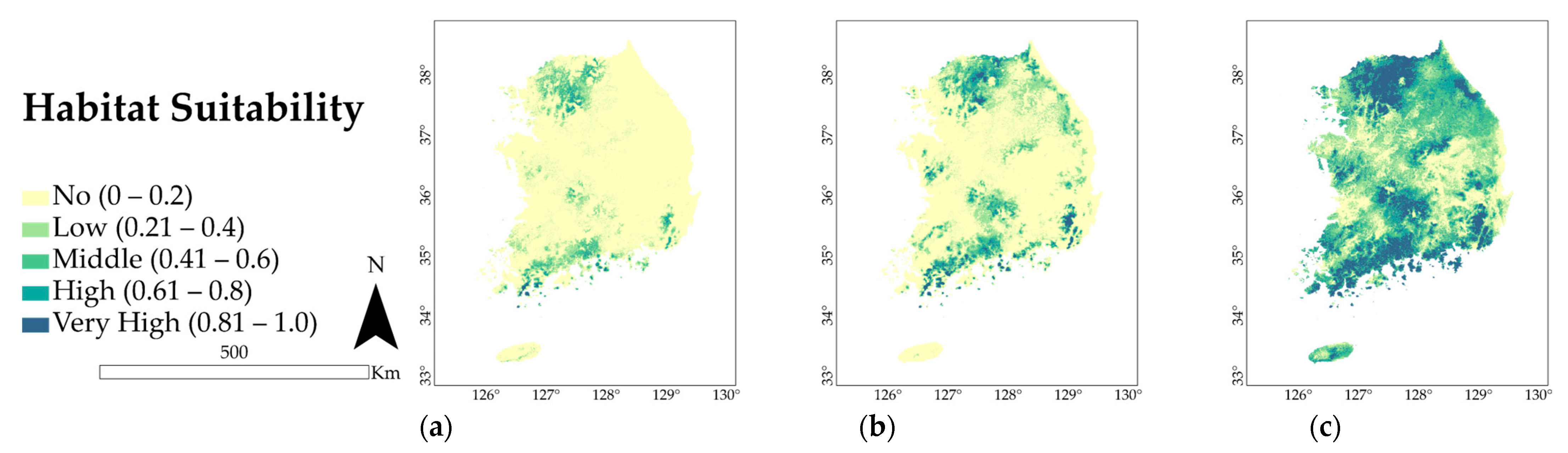
3.2. Present Distribution and Extent of Occurrence
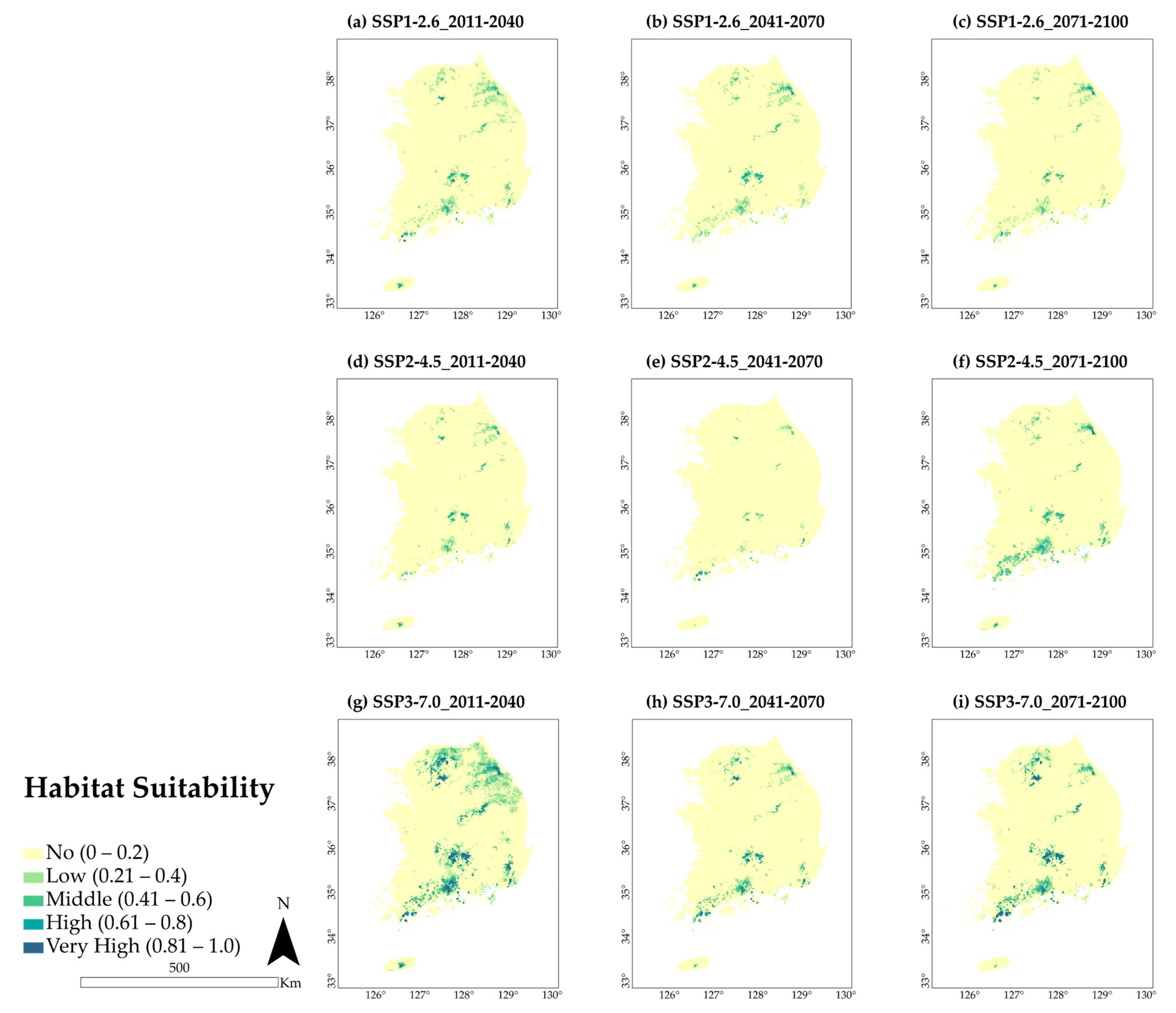
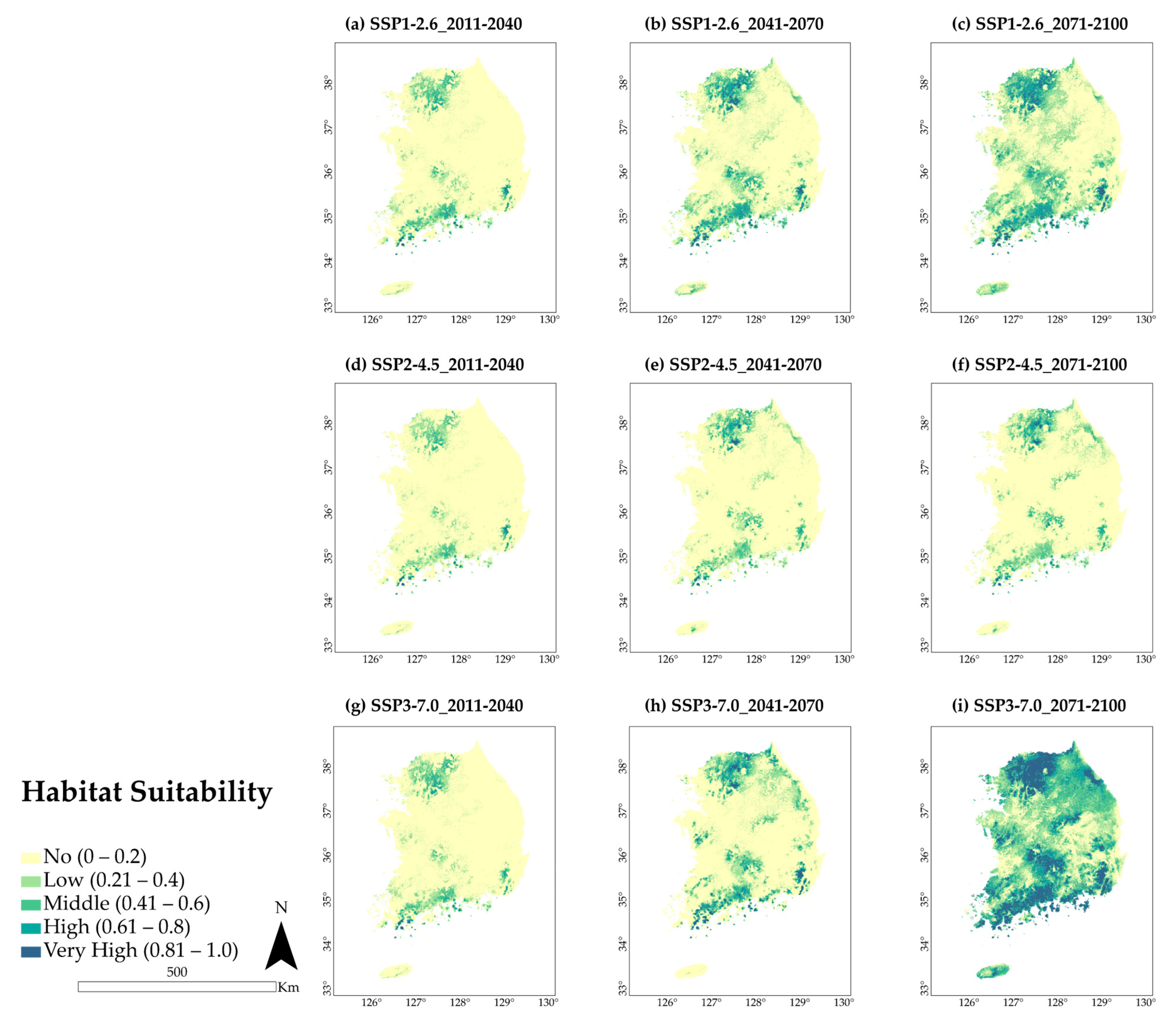
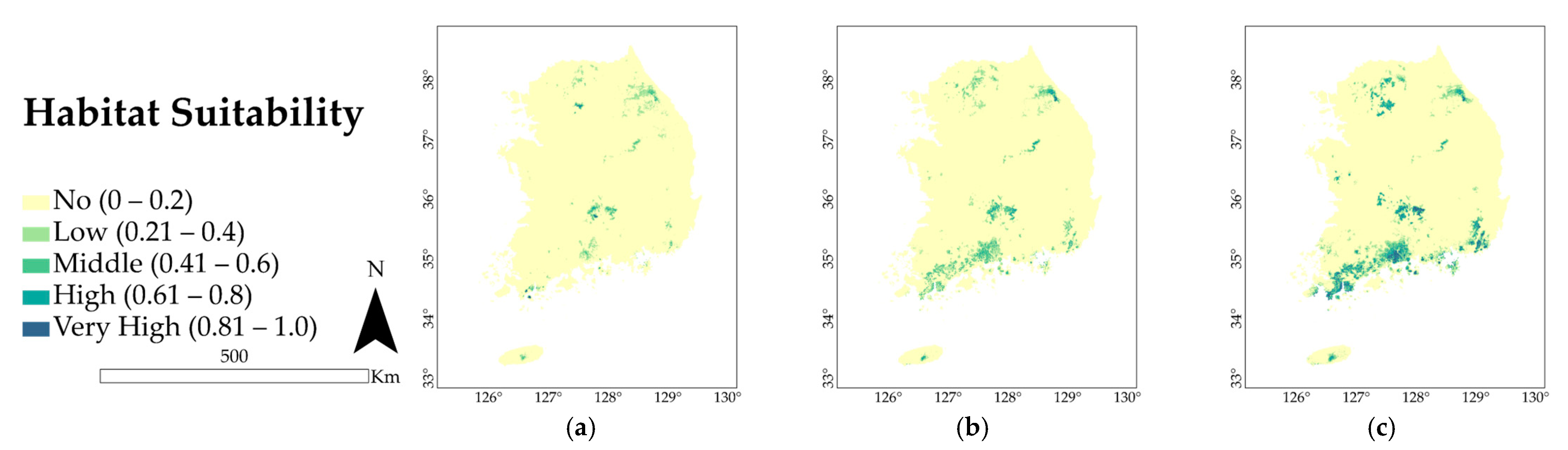
3.3. Future Predicted Distributions
3.4. South Korea STZs
4. Discussion
4.1. Key Environmental Variables and Current Spatial Distribution
4.2. Potential Distribution of A. pictum and Q. acuta under Future Climate Conditions
4.3. How to Use STZs
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Institution (Country) | GCMs | Resolution |
|---|---|---|
| Geophysical Fluid Dynamics Laboratory (USA) | GFDL-ESM4 | 360 × 180 |
| Meteorological Research Institute (Japan) | MRI-ESM2-0 | 320 × 160 |
| Centre National de Recherches Meteorologiques (France) | CNRM-CM-6-1 | 24,572 grids distributed over 128 latitude circles |
| CNRM-ESM2-1 | ||
| Institute Pierre-Simon Laplace (France) | IPSL-CM6A-LR | 144 × 143 |
| Max Planck Institute for Meteorology (Germany) | MPI-ESM1-2-HR | 384 × 192 |
| MPI-ESM1-2-LR | 192 × 96 | |
| Met Office Hadley Centre (UK) | UKESM1-0-LL | 192 × 144 |
| Commonwealth Scientific and Industrial Research Organisation, Australian Rsearch Council Centre of Excellence for Climate System Science (Australia) | ACCESS-CM2 | 192 × 144 |
| Commonwealth Scientific and Industrial Research Organisation (Australia) | ACCESS-ESM1-5 | 192 × 145 |
| Canadian Centre for Climate Modelling and Analysis (Canada) | CanESM5 | 128 × 64 |
| Institute for Numerical Mathematics (Russia) | INM-CM4-8 | 180 × 120 |
| INM-CM5-0 | 180 × 120 | |
| EC-Earth-Consortium | EC-Earth3 | 512 × 256 |
| Japan Agency for Marine-Earth Science and Technology/Atmosphere and Ocean Research Institute/National Institute for Environmental Studies/RIKEN Center for Computational Science (Japan) | MIROC6 | 256 × 128 |
| MIROC-ES2L | 128 × 64 | |
| NorESM Climate modeling Consortium consisting of CICERO (Norway) | NorESM2-LM | 144 × 96 |
| National Institute of Meteorological Sciences/Korea Meteorological Administration (Korea) | KACE-1-0-G | 192 × 144 |

References
- Mijnsbrugge, V.K.; Bischoff, A.; Smith, B. A question of origin: Where and how to collect seed for ecological restoration. Basic Appl. Ecol. 2010, 11, 300–311. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 3–32. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2018; pp. 3–24. [Google Scholar] [CrossRef]
- Shim, S.; Kwon, S.H.; Lim, Y.J.; Yum, S.S.; Byun, Y.H. Understanding climate change over East Asia under stabilized 1.5 and 2.0 °C global warming scenarios. Atmosphere 2019, 29, 391–401. [Google Scholar] [CrossRef]
- Oh, Y.J.; Kim, M.H.; Choi, S.K.; Kim, M.K.; Eo, J.U.; Yeob, S.J. Prediction of the spatial distribution of suitable habitats for Geranium carolinianum under SSP scenarios. Ecol. Resilient Infrastruct. 2021, 8, 154–163. [Google Scholar] [CrossRef]
- Xu, W.; Sun, H.; Jin, J.; Cheng, J. Predicting the Potential Distribution of Apple Canker Pathogen (Valsa mali) in China under Climate Change. Forests 2020, 11, 1126. [Google Scholar] [CrossRef]
- Canturk, U.; Kulaç, Ş. The effects of climate change scenarios on Tilia ssp. in Turkey. Environ. Monit. Assess. 2021, 193, 1–15. [Google Scholar] [CrossRef]
- Gao, X.; Liu, J.; Huang, Z. The impact of climate change on the distribution of rare and endangered tree Firmiana kwangsiensis using the Maxent modeling. Ecol. Evol. 2022, 12, e9165. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.J.; Kim, H.N. Changes in Major Crop Allocations and Shifts under Climate Change in Korea. Korean J. Agric. Manag. Policy 2022, 49, 191–211. [Google Scholar] [CrossRef]
- Yu, D.S.; Kwon, O.C.; Shin, M.S.; Kim, J.K.; Lee, S.H. Effects of Climatic Factors on the Nationwide Distribution of Wild Aculeata (Insecta: Hymenoptera). Korean J. Environ. Ecol. 2022, 36, 303–317. [Google Scholar] [CrossRef]
- O’Neill, B.C.; Kriegler, E.; Ebi, K.L.; Kem-Benedict, E.; Riahi, K.; Rothman, D.S.; Ruijven, B.J.; Vuuren, D.P.; Birkmann, J.; Kok, K.; et al. The roads ahead: Narratives for shared socioeconomic pathways describing world futures in the 21st century. Glob. Env. Chang. 2017, 42, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Booth, T.H. Why understanding the pioneering and continuing contributions of BIOCLIM to species distribution modelling is important. Austral Ecol. 2018, 43, 852–860. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L. SDM toolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explenation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and acomprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Kim, C.Y.; Kim, W.M.; Song, W.K.; Choi, J.Y. A study on the range of native seed habitat analysis using Seed zones and MaxEnt. J. Korean Soc. Environ. Restor. Technol. 2022, 25, 57–74. [Google Scholar] [CrossRef]
- Jian, S.; Zhu, T.; Wang, J.; Yan, D. The Current and Future Potential Geographical Distribution and Evolution Process of Catalpa bungei in China. Forests 2022, 13, 96. [Google Scholar] [CrossRef]
- Li, Y.; Shao, W.; Jiang, J. Predicting the potential global distribution of Sapindus mukorossi under climate change based on MaxEnt modelling. Environ. Sci. Pollut. Res. 2022, 29, 21751–21768. [Google Scholar] [CrossRef]
- Shao, M.; Wang, L.; Li, B.; Li, S.; Fan, J.; Li, C. Maxent Modeling for Identifying the Nature Reserve of Cistanche deserticola Ma under Effects of the Host (Haloxylon Bunge) Forest and Climate Changes in Xinjiang, China. Forests 2022, 13, 189. [Google Scholar] [CrossRef]
- Kwon, H.S.; Ryu, J.E.; Seo, C.W.; Kim, J.Y.; Lim, D.O.; Suh, M.H. A study on distribution characteristics of corylopsis coreana using SDM. J. Environ. Impact Assess. 2012, 21, 735–743. [Google Scholar] [CrossRef]
- Yu, S.B.; Kim, B.D.; Shin, H.T.; Kim, S.J. Habitat climate characteristics of Lauraceae evergreen broad-leaved trees and distribution change according to climate change. Korean J. Environ. Ecol. 2020, 34, 503–514. [Google Scholar] [CrossRef]
- Campbell, R.K. Soils, Seed-Zone Maps, and Physiography: Guidelines for Seed Transfer of Douglas-Fir in Southwestern Oregon. For. Sci. 1991, 37, 4973–4986. [Google Scholar] [CrossRef]
- Crow, T.M.; Albeke, S.E.; Buerkle, C.A.; Hufford, K.M. Provisional methods to guide species-specific seed transfer in ecological restoration. Ecosphere 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Omernik, J.M.; Chapman, S.S.; Lillie, R.A.; Dumke, R.T. Transactions of the Wisconsin Academy of Sciences, Arts and Letters. Ecoregions Wis. 2000, 88, 77–103. [Google Scholar]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; Loucks, C.J.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Bower, A.D.; Bradley, J.; Clair, S.; Erickson, V. Generalized provisional seed zones for native plants. Ecol. Appl. 2014, 24, 913–919. [Google Scholar] [CrossRef]
- Doherty, K.D.; Butterfield, B.J.; Wood, T.E. Matching seed to site by climate similarity: Techniques to prioritize plant materials development and use in restoration. Ecol. Appl. 2017, 27, 1010–1023. [Google Scholar] [CrossRef]
- Lee, B.; Chung, J.; Kwon, D.S. Analysis of site suitability of forest stands for extracting sap of Acer pictum var. mono using GIS and fuzzy sets. J. Korean Soc. For. Sci. 2006, 95, 38–44. [Google Scholar]
- Lee, C.B. Dendrology; Hangmoon Pub. Co.: Seoul, Republic of Korea, 1990; Volume 253. [Google Scholar]
- Um, T.W.; Kim, G.T. Distribution and Growth Characteristics of Acer pictum var mono in Relation to Topography and Soil in Mt. Joongwang, Gangwon Province. Korean Soc. Environ. Ecol. 2006, 20, 200–207. [Google Scholar]
- Song, J.H.; Hur, S.D. Analysis of leaf morphological variation of 11 natural populations of Acer pictum subsp mono (Maxim.) Ohashi. Korean J. Plant Resour. 2011, 24, 540–548. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.T.; Kim, H.J.; Lee, J.H. Studies on the Community Structure, Samara and Leaf Shape of Three Natural Acer pictum subsp mono Forest. Korean J. Environ. Ecol. 2014, 28, 55–61. [Google Scholar] [CrossRef]
- Lee, C.B. Coloured Flora of Korea; Hyangmunsa: Seoul, Republic of Korea, 2014; Volume 1828. [Google Scholar]
- Oh, K.K. Plant community Structure of Evergreen broad-Leaved Forest in Mt. Turyunsan, Korea. Korean J. Environ. Ecol. 1994, 8, 43–57. [Google Scholar]
- Shin, H.C.; Park, N.C.; Song, H.K. The Vegetation Structure and Community Clssification of Quercus acuta in Warm-Temperate Region of Korean Peninsula. KFRI J. For. Sci. 1999, 60, 11–25. [Google Scholar]
- Yeo, U.S. Natural Regeneration Patterns and Strategies of Quercus acuta in Wando, Korea. Ph.D. Dissertation, Seoul National University, Seoul, Republic of Korea, 2005. [Google Scholar]
- Park, I.H. Structure and Dynamics of Quercus acuta, Quercus acutissima and Pinus rigida forests in Wando island. Korean J. Environ. Ecol. 2012, 26, 406–411. [Google Scholar]
- Kim, S.; Park, I.H. Growth and Fruiting Characteristics and No. of Acorns/tree Allometric Equations of Quercus acuta Thunb. in Wando Island, Korea. Korean J. Environ. Ecol. 2019, 33, 440–446. [Google Scholar] [CrossRef]
- Kim, C.S.; Lee, K.Y.; Koh, J.G.; Ryu, K.O.; Kang, Y.J. Correlations between Growth and Isozyme Variation in Open-Pollinated Progenies of Machius thunbergii. Res. Rep. For. Gen. Res. Inst. Korea 1995, 31, 53–60. [Google Scholar]
- Lee, S.H.; Lee, H.S.; Park, Y.S.; Hwang, B.; Kim, J.H.; Lee, H.Y. Screening of Immune Activation Activities in the Leaves of Dendropanax morbifera Lev. Korean J. Med. Crop Sci. 2002, 10, 109–115. [Google Scholar]
- Cha, Y.J.; Lee, J.W.; Kim, J.H.; Park, M.H.; Lee, S.Y. Major Components of Teas Manufactured with Leaf and Flower of Korean Native Camellia japonica L. Korean J. Med. Crop Sci. 2004, 12, 183–190. [Google Scholar]
- Lee, S.T.; Son, Y.M.; Lee, K.J.; Hwang, J.; Choi, J.C.; Shin, H.C.; Park, N.C. Aboveground carbon storage of Quercus acuta stands by thinning intensity. Korean J. Agric. For. Meteorol. 2005, 7, 282–288. [Google Scholar]
- Kang, H.M. Vegetation Characteristics of Evergreen Broad-Leaved Forest in the Duryunsan Provincial Park—Focusing on the Daeheungsa(Temple) Area. Korean J. Environ. Ecol. 2019, 33, 552–564. [Google Scholar] [CrossRef]
- Padalia, H.; Srivastava, V.; Kushwaha, S.P.S. Modeling potential invasion range of alien invasive species, Hyptis suaveolens (L.) Poit. in India: Comparison of MaxEnt and GARP. Ecol. Inform. 2014, 22, 36–43. [Google Scholar] [CrossRef]
- Ma, B.; Sun, J. Predicting the distribution of Stipa purpurea across the Tibetan Plateau via the MaxEnt model. BMC Ecol. 2018, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, M.S.; Ignizio, D.A. Bioclimatic Predictors for Supporting Ecological Applications in the Conterminous United States. US Geol. Surv. Data Ser. 2012, 691, 4–9. Available online: https://pubs.usgs.gov/ds/691/ (accessed on 27 June 2022).
- Hong, K.O.; Suh, M.S.; Rha, D.K.; Chang, D.H.; Kim, C.; Kim, M.K. Estimation of high resolution gridded temperature using GIS and PRISM. Atmosphere 2007, 17, 255–268. [Google Scholar]
- Cho, J.P.; Kim, J.U.; Choi, S.K.; Hwang, S.W.; Jung, H.C. Variability analysis of climate extreme index using downscaled multi-models and grid-based CMIP5 climate change scenario data. J. Clim. Chang. Res. 2020, 11, 123–132. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M.; et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Yackulic, C.B.; Chandler, R.; Zipkin, E.F.; Royle, J.A.; Nichols, J.D.; Campbell Grant, E.H.; Veran, S. Presence-only modelling using MAXENT: When can we trust the inferences? Methods Ecol. Evol. 2013, 4, 236–243. [Google Scholar] [CrossRef]
- Swets, J. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Youtie, B.; Shaw, N.; Fisk, M.; Jensen, S. A strategy for maximizing native plant material diversity for ecological restoration, germplasm conservation and genecology research. In Proceedings of the 8th European Conference on Ecological Restoration, České Budějovice, Czech Republic, 9–14 September 2012; pp. 9–14. [Google Scholar]
- Bower, A.D. Ecological genetics and seed transfer guidelines for Pinus albicaulis (Pinaceae). Am. J. Bot. 2008, 95, 66–76. [Google Scholar] [CrossRef]
- Johnson, R.C.; Erickson, V.J.; Mandel, N.L.; St Clair, J.B.; Vance-Borland, K.W. Mapping genetic variation and seed zones for Bromus carinatus in the Blue Mountains of eastern Oregon, USA. Botany 2010, 88, 725–736. [Google Scholar] [CrossRef]
- Pike, C.; Potter, K.M.; Berrang, P.; Crane, B.; Baggs, J.; Leites, L.; Luther, T. New seed-collection zones for the eastern United States: The eastern seed zone forum. J. For. 2020, 118, 444–451. [Google Scholar] [CrossRef] [Green Version]
- Schubert, G.H.; Pitcher, J.A. A Provisional Tree Seed-Zone and Cone-Crop Rating System for Arizona and New Mexico; Rocky Mountain Forest and Range Experiment Station, Forest Service, US Department of Agriculture: Fort Collins, CO, USA, 1973; Volume 105. [Google Scholar]
- Ying, C.C.; Yanchu, A.D. The development of British Columbia’s tree seed transfer guidelines:Purpose, concept, methodology, and implementation. For. Ecol. Manag. 2006, 227, 1–13. [Google Scholar] [CrossRef]
- Bezeng, B.S.; Morales-Castilla, I.; Bank, M.; Yessoufou, K.; Daru, B.H.; Davies, T.J. Climate change may reduce the spread of non-native species. Ecosphere 2017, 8, e01694. [Google Scholar] [CrossRef]
- Hamann, A.; Wang, T.L. Models of climatic normals for genecology and climate change studies in British Columbia. Agric. For. Meteorol. 2005, 128, 211–221. [Google Scholar] [CrossRef]
- Lee, S.H.; Heo, I.H.; Lee, K.M.; Kwon, W.T. Classification of local climatic regions in Korea. J. Korean Meteorol. Soc. 2005, 41, 983–995. [Google Scholar]
- Byun, J.G.; Lee, W.K.; Nor, D.K.; Kim, S.H.; Choi, J.K.; Lee, Y.J. The relationship between tree radial growth and topographic and climatic factors in red pine and oak in central regions of Korea. J. Korean Soc. For. Sci. 2010, 99, 908–913. [Google Scholar]
- Lee, S.; Choi, S.; Lee, W.K.; Park, T.; Oh, S.; Kim, S.N. Vulnerability assessment of forest distribution by the climate change scenarios. J. Korean Soc. For. Sci. 2011, 100, 256–265. [Google Scholar] [CrossRef]
- Koo, K.A.; Kim, J.; Kong, W.S.; Jung, H.; Kim, G. Projecting the potential distribution of Abies koreana in Korea under the climate change based on RCP scenarios. J. Korean Soc. Environ. Restor. Technol. 2016, 19, 19–30. [Google Scholar] [CrossRef]
- Choi, Y.E.; Choi, J.Y.; Kim, W.M.; Kim, S.Y.; Song, W.K. Long-term Effects on Forest Biomass under Climate Change Scenarios Using LANDIS-II-A case study on Yoengdong-gun in Chungcheongbuk-do, Korea. J. Korean Soc. Environ. Restor. Technol. 2019, 22, 27–43. [Google Scholar] [CrossRef]
- Kim, C.Y.; Kim, W.M.; Song, W.K.; Choi, J.Y. A Study on the Domestic Appllication of the Concept of Seed Transfer Zone in the US. J. Korean Soc. Environ. Restor. Technol. 2021, 24, 39–56. [Google Scholar] [CrossRef]
- Park, I.H.; Kim, S.D.; Park, J.W.; Chae, K.S.; Kim, G.T.; Um, T.W. Flowering Characteristics and Acorn Production of Quercus auta Thunb. in Wando Island, Korea. Korean J. Environ. Ecol. 2014, 28, 326–330. [Google Scholar] [CrossRef]
- Shin, M.Y.; Chung, S.Y.; Han, W.S.; Lee, D.K. Effects of microclimate of different site types on tree growth in natural deciduous forest. Korean J. Agric. For. Meteorol. 2008, 10, 9–16. [Google Scholar] [CrossRef]
- Yang, H.M.; Kim, J.H. Selection of Desirable Species by the Estimation of Dominant and Potential Dominant Species in the Natural Deciduous Forest. For. Bioenergy 2002, 21, 77–88. [Google Scholar]
- Um, T.W.; Lee, D.K. Distribution of major deciduous tree species in relation to the characteristics of topography in Mt. Joongwang, Gangwon Province (I). J. Korean Soc. For. Sci. 2006, 95, 91–101. [Google Scholar]
- Shin, M.S.; Seo, C.; Park, S.U.; Hong, S.B.; Kim, J.Y.; Jeon, J.Y.; Lee, M. Prediction of potential habitat of Japanese evergreen oak (Quercus acuta Thunb.) considering dispersal ability under climate change. J. Environ. Impact Assess. 2018, 27, 291–306. [Google Scholar] [CrossRef]
- Park, J.H.; Jung, S.Y.; Lee, K.S.; Lee, H.S. The Characteristics and Survival Rates of Evergreen Broad-Leaved Tree Plantations in Korea. J. Korean Soc. For. Sci. 2019, 108, 513–521. [Google Scholar] [CrossRef]
- Yang, K.C.; Shim, J.K. Distribution of major plant communities based on the climatic conditions and topographic features in South Korea. Korean J. Environ. Biol. 2007, 25, 168–177. [Google Scholar]
- Nakao, K.; Matsui, T.; Horikawa, M.; Tsuyama, I.; Tanaka, N. Assessing the impact of land use and climate change on the evergreen broad-leaved species of Quercus acuta in Japan. Plant Ecol. 2011, 212, 229–243. [Google Scholar] [CrossRef]
- Yun, J.H.; Nakao, K.; Park, C.H.; Lee, B.Y.; Oh, K.H. Change prediction for potential habitats of warm-temperate evergreen broad-leaved trees in Korea by climate change. Korean J. Environ. Ecol. 2011, 25, 590–600. [Google Scholar]
- Oh, S.N.; Ha, K.J.; Kim, K.Y.; Kim, J.W. Effects of land hydrology in northeastern asia in a doubling CO2 climate experiment. Korean J. Atmos. Sci. 1998, 34, 293–305. [Google Scholar]
- Park, H.S. Acer pictum subsp. mono properties and proliferation. Landscaping Tree 2002, 70, 21–23. [Google Scholar]
- An, J.M.; Kim, J.S.; Kang, H.M. A study on Patterns of Sap Water Users of Acer mono. J. Korean Soc. For. Sci. 1998, 87, 510–518. [Google Scholar]
- Cha, D.W.; Oh, C.H. Flora and Vegetation Characteristics of Gwangyang Mt. Baegun ‘Acer pictum Thunb. var mono (Maxim.) Maxim. ex Franch’ Plantation. Proc. Korean Soc. Environ. Ecol. Conf. 2021, 31, 3. [Google Scholar]
- Kwon, S.H.; Kim, J.S.; Byun, Y.H.; Bu, K.O.; Seo, J.B.; Seon, M.A.; Seong, H.M.; Shim, S.B.; Lee, J.H.; Lim, Y.J. Revision of the Global Outlook Report; National Institute of Meteorological Sciences: Seogwipo, Republic of Korea, 2020. [Google Scholar]
- Park, S.U.; Koo, K.A.; Kong, W.S. Potential Impact of Climate Change on Distribution of Warm Temperate Evergreen Broad-leaved Trees in the Korean Peninsula. J. Korean Geogr. Soc. 2016, 51, 201–217. [Google Scholar]
- Kim, K.H.; Choi, J.Y. Ecological Restoration Methods Development Using Native Speciesin DMZ Vicinities; Ministry of Environment: Tokyo, Japan, 2017; pp. 1–1041. [Google Scholar] [CrossRef]
- Hu, X.G.; Wang, T.; Liu, S.S.; Jiao, S.Q.; Jia, K.H.; Zhou, S.S.; Jin, Y.; Li, Y.; El-Kassaby, Y.A.; Mao, J.F. Predicting future seed sourcing of Platycladus orientalis (L.) for future climates using climate niche models. Forests 2017, 8, 471. [Google Scholar] [CrossRef] [Green Version]
- Eckert, A.J.; Bower, A.D.; GONZÁLEZ-MARTÍNEZ, S.C.; Wegrzyn, J.L.; Coop, G.; Neale, D.B. Back to nature: Ecological genomics of loblolly pine (Pinus taeda, Pinaceae). Mol. Ecol. 2010, 19, 3789–3805. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.C.; Cashman, M.J.; Vance-Borland, K. Genecology and seed zones for Indian ricegrass collected in the southwestern United States. Rangel. Ecol. Manag. 2012, 65, 523–532. [Google Scholar] [CrossRef]
- Keller, M.; Kollmann, J. Effects of seed provenance on germination of herbs for agricultural compensation sites. Agric. Ecosyst. Environ. 1999, 72, 87–99. [Google Scholar] [CrossRef]
- Pike, C.C.; Hernandez, G.; Crane, B.; Berrang, P. Development for Seed Zones in the Eastern United States: Request for Input and Collaboration! General Technical Report PNW-GTR-963; U.S. Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2017; Volume 30. Available online: https://www.fs.usda.gov/research/treesearch/55237 (accessed on 21 October 2022).
- John, J.G.; Blanton, C.; McHugh, C.; Radhakrishnan, A.; Rand, K.; Vahlenkamp, H.; Wilson, C.; Zadeh, N.T.; Dunne, J.P.; Dussin, R.; et al. NOAA-GFDL GFDL-ESM4 Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2018. [Google Scholar] [CrossRef]
- Yukimoto, S.; Koshiro, T.; Kawai, H.; Oshima, N.; Yoshida, K.; Urakawa, S.; Tsujino, H.; Deushi, M.; Tanaka, T.; Hosaka, M.; et al. MRI MRI-ESM2.0 Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Voldoire, A. CNRM-CERFACS CNRM-CM6-1 Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Seferian, R. CNRM-CERFACS CNRM-ESM2-1 Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Boucher, O.; Denvil, S.; Levavasseur, G.; Cozic, A.; Caubel, A.; Foujols, M.A.; Meurdesoif, Y.; Cadule, P.; Devilliers, M.; Dupont, E.; et al. IPSL IPSL-CM6A-LR Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Schupfner, M.; Wieners, K.H.; Wachsmann, F.; Steger, C.; Bittner, M.; Jungclaus, J.; Früh, B.; Pankatz, K.; Giorgetta, M.; Reick, C.; et al. CMIP6 CMIP DKRZ MPI-ESM1-2-HR amip-RCM-Forcing Data; World Data Center for Climate (WDCC) at DKRZ: Hamburg, Germany, 2019. [Google Scholar] [CrossRef]
- Wieners, K.H.; Giorgetta, M.; Jungclaus, J.; Reick, C.; Esch, M.; Bittner, M.; Gayler, V.; Haak, H.; de Vrese, P.; Raddatz, T.; et al. MPI-M MPIESM1.2-LR Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Good, P.; Sellar, A.; Tang, Y.; Rumbold, S.; Ellis, R.; Kelley, D.; Kuhlbrodt, T.; Walton, J. MOHC UKESM1.0-LL Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Dix, M.; Bi, D.; Dobrohotoff, P.; Fiedler, R.; Harman, I.; Law, R.; Mackallah, C.; Marsland, S.; O’Farrell, S.; Rashid, H.; et al. CSIRO-ARCCSS ACCESS-CM2 Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Ziehn, T.; Chamberlain, M.; Lenton, A.; Law, R.; Bodman, R.; Dix, M.; Wang, Y.; Dobrohotoff, P.; Srbinovsky, J.; Stevens, L.; et al. CSIRO ACCESS-ESM1.5 Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Swart, N.C.; Cole, J.N.S.; Kharin, V.V.; Lazare, M.; Scinocca, J.F.; Gillett, N.P.; Anstey, J.; Arora, V.; Christian, J.R.; Jiao, Y.; et al. CCCma CanESM5 model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Volodin, E.; Mortikov, E.; Gritsun, A.; Lykossov, V.; Galin, V.; Diansky, N.; Gusev, A.; Kostrykin, S.; Iakovlev, N.; Shestakova, A.; et al. INM INM-CM4-8 Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Volodin, E.; Mortikov, E.; Gritsun, A.; Lykossov, V.; Galin, V.; Diansky, N.; Gusev, A.; Kostrykin, S.; Iakovlev, N.; Shestakova, A.; et al. INM INM-CM5-0 Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- EC-Earth Consortium (EC-Earth). EC-Earth-Consortium EC-Earth3 Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Shiogama, H.; Abe, M.; Tatebe, H. MIROC MIROC6 Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Tachiiri, K.; Abe, M.; Hajima, T.; Arakawa, O.; Suzuki, T.; Komuro, Y.; Ogochi, K.; Watanabe, M.; Yamamoto, A.; Tatebe, H.; et al. MIROC MIROC-ES2L Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Seland, Ø.; Bentsen, M.; Oliviè, D.J.L.; Toniazzo, T.; Gjermundsen, A.; Graff, L.S.; Debernard, J.B.; Gupta, A.K.; He, Y.; Kirkevåg, A.; et al. NCC NorESM2-LM Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]
- Byun, Y.H.; Lim, Y.J.; Shim, S.; Sung, H.M.; Sun, M.; Kim, J.; Kim, B.H.; Lee, J.H.; Moon, H. NIMS-KMA KACE1.0-G Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation: Seattle, WA, USA, 2019. [Google Scholar] [CrossRef]

| Environmental Variables | Description | Unit | Data Type | Data Soure | |
|---|---|---|---|---|---|
| Climate | Bio01 | Annual Mean Temperature | °C | Continuous | Rural Development Administration based SSPs Scenario |
| Bio02 | Mean Diurnal Range | °C | Continuous | ||
| Bio04 | Temperature Seasonality | °C | Continuous | ||
| Bio08 | Mean Temperature of Wettest Quarter | °C | Continuous | ||
| Bio12 | Annual Precipitation | mm | Continuous | ||
| Bio15 | Precipitation Seasonality | % | Continuous | ||
| Bio18 | Precipitation of Warmest Quarter | mm | Continuous | ||
| Topography | Elevation | DEM | m | Continuous | Ministry of Environment, Korea (2020) |
| Slope | Slope | % | Continuous | ||
| Aspect | Aspect | ° | Continuous | ||
| Drainage | Soil drainage grade | - | Categorical | Forest Service, Korea (2020) | |
| Terrain | Forest Soil Map | - | Categorical |
| Environmental Variables | A. pictum | Q. acuta | ||
|---|---|---|---|---|
| Percent Contribution (%) | Permutation Importance (%) | Percent Contribution (%) | Permutation Importance (%) | |
| Bio01 | 0.1 | 0.0 | 0.0 | 0.0 |
| Bio02 | 0.0 | 0.0 | 2.5 | 3.8 |
| Bio04 | 2.9 | 2.3 | 5.4 | 3.9 |
| Bio08 | 11.2 | 28.0 | 14.7 | 41.5 |
| Bio12 | 23.8 | 31.7 | 56.7 | 24.4 |
| Bio15 | 0.6 | 0.9 | 0.0 | 0.0 |
| Bio18 | 1.2 | 2.0 | 1.3 | 0.0 |
| Elevation | 9.1 | 3.0 | 1.4 | 0.4 |
| Slope | 23.0 | 17.6 | 8.4 | 20.9 |
| Aspect | 0.9 | 0.0 | 0.1 | 0.0 |
| Drainage | 26.9 | 13.8 | 1.3 | 1.6 |
| Terrain | 0.3 | 0.6 | 8.1 | 3.3 |
| Suitability Grade | SSP Historical Period | SSP1-2.6 | SSP2-4.5 | SSP3-7.0 | SSP5-8.5 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Near Future | Mid Future | Far Future | Near Future | Mid Future | Far Future | Near Future | Mid Future | Far Future | Near Future | Mid Future | Far Future | ||
| No (%) | 57.83 | 92.90 | 94.36 | 96.05 | 96.20 | 98.38 | 93.44 | 81.44 | 93.89 | 91.18 | 96.90 | 93.46 | 89.75 |
| Low (%) | 19.54 | 5.41 | 4.15 | 2.88 | 2.69 | 1.15 | 3.79 | 10.75 | 3.50 | 4.55 | 2.46 | 4.18 | 4.60 |
| Middle (%) | 11.23 | 1.15 | 1.10 | 0.80 | 0.75 | 0.26 | 1.88 | 3.99 | 1.34 | 2.11 | 0.48 | 1.86 | 2.53 |
| High (%) | 7.97 | 0.11 | 0.35 | 0.22 | 0.32 | 0.15 | 0.84 | 2.05 | 0.84 | 1.23 | 0.11 | 0.47 | 2.38 |
| Very High (%) | 3.41 | 0.10 | 0.02 | 0.02 | 0.04 | 0.06 | 0.05 | 1.77 | 0.43 | 0.92 | 0.01 | 0.01 | 0.72 |
| Suitability Grade | SSP Historical Period | SSP1-2.6 | SSP2-4.5 | SSP3-7.0 | SSP5-8.5 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Near Future | Mid Future | Far Future | Near Future | Mid Future | Far Future | Near Future | Mid Future | Far Future | Near Future | Mid Future | Far Future | ||
| No (%) | 99.88 | 89.88 | 70.40 | 54.74 | 87.26 | 82.21 | 78.85 | 86.88 | 56.01 | 41.65 | 84.76 | 70.72 | 20.18 |
| Low (%) | 0.09 | 9.89 | 15.91 | 23.00 | 8.52 | 10.86 | 12.92 | 8.76 | 23.24 | 24.53 | 9.66 | 15.23 | 22.42 |
| Middle (%) | 0.02 | 4.30 | 7.25 | 11.60 | 3.02 | 4.75 | 5.63 | 3.11 | 10.75 | 16.46 | 3.99 | 7.46 | 22.60 |
| High (%) | 0.008 | 1.61 | 4.84 | 7.81 | 0.98 | 1.79 | 2.14 | 0.99 | 6.88 | 11.50 | 1.31 | 4.58 | 17.62 |
| Very High (%) | 0.005 | 0.32 | 1.60 | 2.85 | 0.22 | 0.39 | 0.45 | 0.27 | 3.12 | 5.86 | 0.28 | 2.01 | 17.18 |
| No | A. pictum | Q. acuta | |||
|---|---|---|---|---|---|
| Scenario | Seed Zone (°F, °C/m) | Ratio (%) | Seed Zone (°F, °C/m) | Ratio (%) | |
| Historical | 15–20, 16–19 | 14.55 | 25–30, 19–21 | 39.10 | |
| SSP1-2.6 | Near future | 10–15, 16–19 | 15.07 | 10–15, 19–21 | 21.58 |
| Mid future | 10–15, 16–19 | 21.77 | 10–15, 19–21 | 19.25 | |
| Far future | 25–30, 19–21 | 21.35 | 10–15, 19–21 | 16.93 | |
| SSP2-4.5 | Near future | 15–20, 16–19 | 17.98 | 10–15, 19–21 | 24.93 |
| Mid future | 25–30, 19–21 | 37.78 | 10–15, 19–21 | 21.36 | |
| Far future | 20–25, 16–19 | 22.72 | 10–15, 16–19 | 18.11 | |
| SSP3-7.0 | Near future | 15–20, 16–19 | 17.20 | 20–25, 19–21 | 22.52 |
| Mid future | 20–25, 16–19 | 18.85 | 20–25, 19–21 | 17.96 | |
| Far future | 15–20, 16–19 | 17.59 | 20–25, 19–21 | 16.53 | |
| SSP5-8.5 | Near future | 25–30, 19–21 | 18.40 | 10–15, 19–21 | 28.13 |
| Mid future | 20–25, 16–19 | 23.13 | 10–15, 19–21 | 16.86 | |
| Far future | 20–25, 19–21 | 20.63 | 15–20, 19–21 | 20.08 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, C.; Kim, W.; Song, W.; Cho, J.; Choi, J. Prediction of Native Seed Habitat Distribution According to SSP Scenario and Seed Transfer Zones: A Focus on Acer pictum subsp. mono and Quercus acuta. Forests 2023, 14, 87. https://doi.org/10.3390/f14010087
Kim C, Kim W, Song W, Cho J, Choi J. Prediction of Native Seed Habitat Distribution According to SSP Scenario and Seed Transfer Zones: A Focus on Acer pictum subsp. mono and Quercus acuta. Forests. 2023; 14(1):87. https://doi.org/10.3390/f14010087
Chicago/Turabian StyleKim, Chaeyoung, Wheemoon Kim, Wonkyong Song, Jaepil Cho, and Jaeyong Choi. 2023. "Prediction of Native Seed Habitat Distribution According to SSP Scenario and Seed Transfer Zones: A Focus on Acer pictum subsp. mono and Quercus acuta" Forests 14, no. 1: 87. https://doi.org/10.3390/f14010087